
Abstract
Porcine circovirus 2 (PCV2) capsid protein (Cap) is the major structural protein that is responsible for neutralizing antibodies development and protective immunity, thus it is usually used to develop vaccines against porcine circovirus-associated disease (PCVAD). Porcine CD40 ligand (CD40L) and granulocyte-macrophage colony-stimulating factor (GMCSF) have positive immunostimulatory effects on immunocytes and have been applied in vaccine efficacy improvement as attractive adjuvant cytokines, respectively. However, whether these two cytokines can produce synergistic effect in vaccines still need to be further studied. In this study, porcine CD40L and GMCSF were inserted into recombinant adenoviruses to test the immunogenicity of PCV2 adenovirus vaccine in mice. Western blot and indirect immunofluorescence assay showed that Ad-Cap, Ad-CD40L-Cap, Ad-Cap-GMCSF, and Ad-CD40L-Cap-GMCSF were successfully constructed. Indirect ELISA and virus neutralizing assay showed that CD40L and GMCSF could enhance humoral immune responses, and PCV2 Cap-specific antibody titer and neutralizing activities were significantly higher in Ad-CD40L-Cap-GMCSF group than that in the other groups that just inserted either porcine CD40L or GMCSF in recombinant adenoviruses. Moreover, lymphocyte proliferation assay and cytokine release assay showed that CD40L and GMCSF enhanced the cellular immune responses of Ad-Cap, and had synergistic effects in lymphocyte proliferative activities and Th1-type cytokine production. Following PCV2 challenge, the viral loads in lungs of Ad-CD40L-Cap-GMCSF group were significantly lower compared with Ad-Cap, Ad-CD40L-Cap, and Ad-Cap-GMCSF group. Taken together, the results of this study demonstrated that CD40L and GMCSF could synergistically enhance the protective immune responses of PCV2 adenovirus vaccine, which would be used as a potent vaccine for the prevention and control of PCVAD.
Introduction
P
Previous studies have shown that cytokines could be applied as molecular adjuvants to increase the immunogenicity of vaccines when coexpressed with the antigen, such as interferon (IFN)-γ, interleukin (IL)-15, CD40 ligand (CD40L), and granulocyte-macrophage colony-stimulating factor (GMCSF), (13,20,21,24,32,54). CD40L, a member of tumor necrosis factor super family, contributes to the development and activation of adaptive immune responses in different species (9,19,34). The interaction of CD40L and its receptor, CD40, stimulates B-cell activation, differentiation, and antibody production, and switches antibody isotype (2,18). GMCSF, a member of colony-stimulating factors, is responsible for stimulating the proliferation and differentiation of medullary mononuclear cells, especially for dendritic cells (DC) (1,14,15). Therefore, it is found to be a nice way to employ CD40L and GMCSF as adjuvants of PCV2 adenovirus vaccine to increase its efficacy.
In the present study, we modified the recombinant adenoviruses expressing Cap with CD40L and GMCSF, alone or both, and evaluated their immune responses in mice. Mouse model has been widely used in PCV2 vaccine researches (13,28,52,54). The results indicated that both CD40L and GMCSF could significantly increase the immune responses and protective efficacy of PCV2 adenovirus vaccine in mice and exhibited synergistic effect.
Materials and Methods
Viruses and cell lines
PCV2 strain (GenBank no. EU366323) was used in this study. PCV2 was propagated and titered in porcine kidney 15 cells (PK15). The tissue culture infectious dose 50 (TCID50) of PCV2 was determined by performed indirect immunofluorescence assay (IFA) according to a previous study (12). PK15 PCV1-free cell line and human embryonic kidney 293 (HEK293) cell line (ATCC, CRL-1573) were cultured in Dulbecco's minimal essential medium (DMEM; Gibco BRL) with 10% heat-inactivated fetal bovine serum (Gibco BRL) at 37°C in an incubator with 5% CO2.
Recombinant adenoviruses construction
The ORF2 gene (without the nuclear localization signal peptide of 41 amino acid residues at N-terminus) was amplified from PCV2 strain and cloned into the puc57 plasmid (GenScript) and named as puc-Cap. The primers for amplifying CD40L and GMCSF were based on porcine CD40L gene (GenBank accession no. NM214126) and porcine GMCSF gene (GenBank accession no. NM214118). CD40L was inserted upstream of Cap, GMCSF was inserted downstream of Cap, and named as puc-CD40L-Cap-GMCSF. The 2A gene of foot-and-mouth disease virus (GenBank accession no. M10975) encodes a 17 amino acid self-cleaving 2A protease (NFDLLKLAGDVESNPGP). The 2A gene was inserted downstream of CD40L and upstream of GMCSF (46) (Fig. 1). Four fragments (Cap, CD40L-Cap, Cap-GMCSF, and CD40L-Cap-GMCSF) were amplified from puc-CD40L-Cap-GMCSF, with XhoI in the upstream and EcoRV in the downstream, respectively. Primers were listed in Table 1. Amplicons were digested with XhoI and EcoRV and cloned into the transfer vector pShuttle-CMV following the instruction of AdEasy™ Adenoviral Vector System (Agilent Technologies). The recombinant plasmids, named as PS-Cap, PS-CD40L-Cap, PS-Cap-GMCSF, and PS-CD40L-Cap-GMCSF, were identified by polymerase chain reaction (PCR), XhoI and EcoRV digestion, and sequence. PmeI-linearized plasmids were cotransformed into Escherichia coli bacteria strain BJ5183 with pAdEasy-1 vector by electroporation with a Bio-Rad Gene Pulser at 2.5 kV, 25 μF and 200 Ω. The transformed bacteria were plated on a lysogeny broth (LB) plate with 50 μg/mL kanamycin and cultured at 37°C for 18 h. Then the recombinant plasmids, named as rAd-Cap, rAd-CD40L-Cap, rAd-Cap-GMCSF and, rAd-CD40L-Cap-GMCSF, were extracted from the culture medium that were shaken over night at 37°C and identified by PCR. A plasmid without Cap was also constructed and named as rAd-wild.
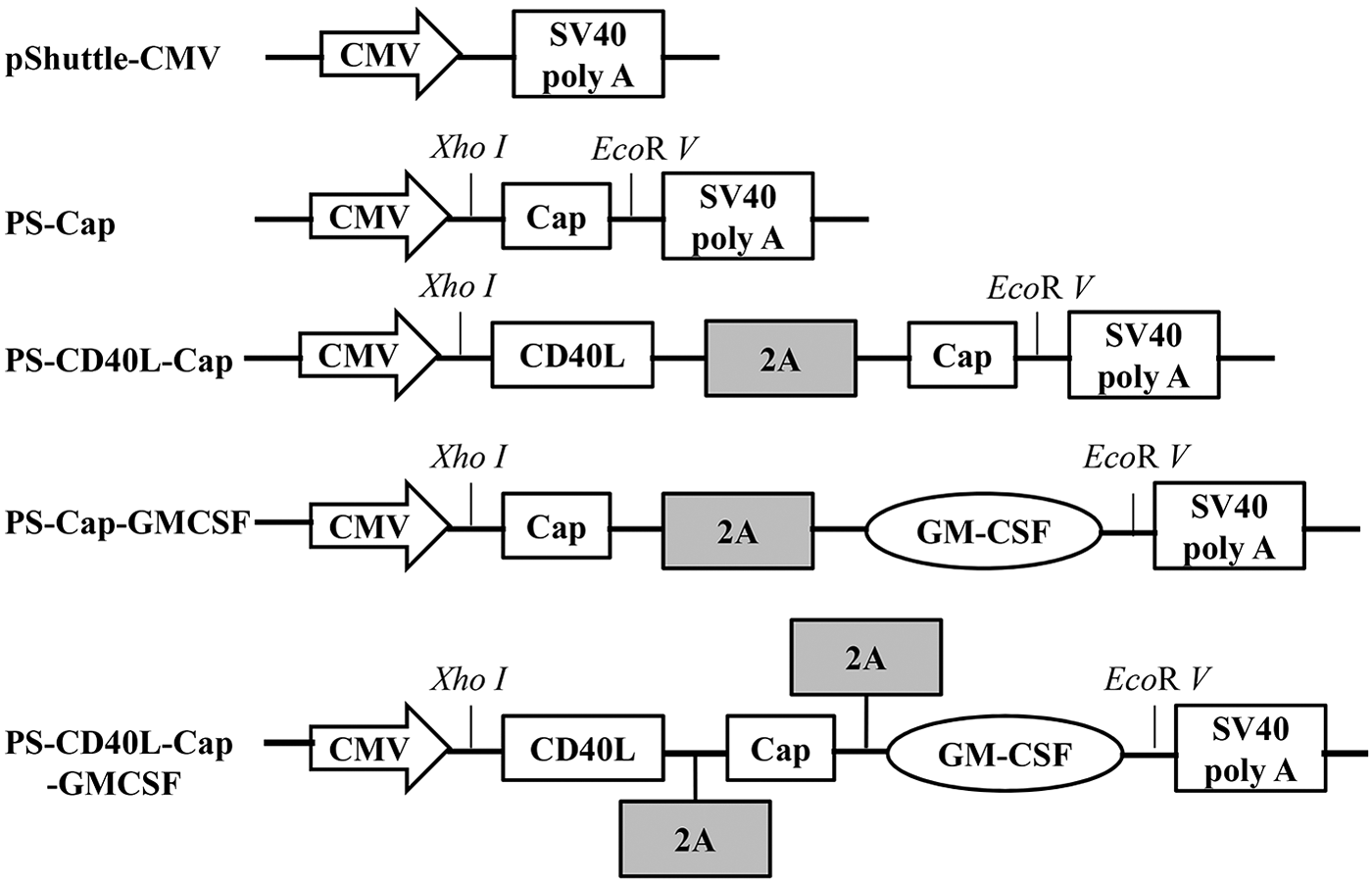
Schematic diagrams of recombinant transfer vectors. The CD40 ligand (CD40L), Cap, and granulocyte-macrophage colony-stimulating factor (GMCSF) gene were cloned into pShuttle-CMV. XhoI was located in the downstream of CMV promoter, and EcoRV was in the upstream of SV40 poly A. 2A gene of foot-and-mouth disease virus was used as a linker to connect PCV2 Cap, CD40L, or GMCSF.
The restriction sites are underlined; the sequences in bold show the 2A gene of foot-and-mouth disease virus used as a linker to connect PCV2 Cap and CD40L or GMCSF.
CD40L, CD40 ligand; GMCSF, granulocyte-macrophage colony-stimulating factor; PCV2, porcine circovirus 2.
Transfection and isolation of recombinant adenoviruses
HEK293 cells were seeded in a 24-well plate overnight before being transfected with 2 μg per well of recombinant plasmids (rAd-Cap, rAd-CD40L-Cap, rAd-Cap-GMCSF, rAd-CD40L-Cap-GMCSF, and rAd-wild) digested with enzyme PacI by applying Lipofectamine 2000 Transfection Reagent (Invitrogen). The viruses were collected 10 days later when a cytopathic effect (CPE) appeared. Then the recombinant adenoviruses were propagated in HEK293 cells and purified thrice with plaque test. Titers of fifth generation virus were detected by the method of TCID50 according to the method of Reed and Muench (45).
Identification of expression of CD40L, GMCSF, and Cap
Western blot
HEK293 cells were cultured in six-well plates overnight and inoculated with recombinant adenoviruses at 20 multiplicity of infection (MOI) for 30 h. Lysates were isolated by 10% SDS-PAGE and transferred to a nitrocellulose membrane. The membrane was put in the blocking solution (5% fat-free milk in phosphate-buffered saline [PBS]) at 4°C overnight and incubated with rabbit anti-Cap, rabbit anti-CD40L, and rabbit anti-GMCSF (polyclonal antibodies prepared by the standard procedures in our laboratory [data not shown]) for 1 h at room temperature and then incubated with goat anti-rabbit IgG conjugated with horseradish peroxidase (HRP) (Boshide) for 1 h. Detection was conducted by chemiluminescence luminal reagents (Super Signal West Pico Trial Kit; Pierce).
Indirect IFA
30 hours after infection by recombinant adenoviruses, the culture plates were washed with 0.05% Tween-20 in PBS (PBS-T) thrice and fixed with cold absolute ethanol for 15 min on ice. After washing thrice, cells were incubated with rabbit anti-Cap, rabbit anti-CD40L, and rabbit anti-GMCSF at 37°C for 1 h. The cells were washed thrice and stained with fluorescein isothiocyanate (FITC)-labeled goat anti-rabbit IgG (Boshide) at 37°C for 1 h. The fluorescent focus was determined by a fluorescent microscope after washing thrice.
Mice immunization with recombinant adenoviruses
One hundred and twenty Kunming mice (6-week old) were purchased from the Animal Center, The Fourth Military Medical University, Xi'an, China. Animal experiments were supported by the Animal Ethics Committee of the North West Agriculture and Forestry University. Mice were randomly divided into six groups and every group contained 20 mice. Mice in group 1–5 were immunized subcutaneously with 108 TCID50 of Ad-Cap, Ad-CD40L-Cap, Ad-Cap-GMCSF, Ad-CD40L-Cap-GMCSF, and Ad-wild, respectively; mice in group 6 were vaccinated with PBS as control. Booster immunization was conducted 14 days later under the same condition. Sera of four mice from each group were randomly collected on day 0, 7, 14, 21, 28, 35, 42, 49, and 56 after the first vaccination for detecting specific antibody and neutralizing antibody, and the sera collection times of every mouse from each group were no more than two. On day 28 after the first immunization, the spleen of four mice in every group were excised and detected by lymphocyte proliferation assay and cytokine release assay. All groups were intraperitoneally challenged with 2 × 104 TCID50 PCV2 on day 28 after the first immunization. Four mice from each group were euthanized on day 28, 35, 42, 49, and 56 after first the immunization and lung samples were harvested for detection of PCV2 viral loads.
Indirect ELISA
PET-32a-ORF2, containing ORF2 gene without the nuclear localization signal, was prepared in our laboratory (data not shown). The recombinant His-Cap protein was purified by the His-Bind purification Kit (Novagen) and applied as indirect ELISA coating protein in 96-well plates with 5 μg/mL. The plates were blocked by 5% fat-free milk in PBS. Sera were diluted in twofold, beginning with 1:100. Then, the plates were incubated at 37°C for 1 h. After washing thrice, goat anti-mouse IgG conjugated with HRP (Boshide) was put into the wells and incubated at 37°C for 1 h. The plates were incubated with substrate solution tetramethyl benzidine at 37°C for 15 min and the reaction was stopped with 2 M H2SO4. The absorbance reading of each well at 490 nm was recorded from a spectrophotometer. Meanwhile, the mice sera inoculated with wild adenovirus or PBS were used as negative control. The results were determined by the ratio of optical density 490 nm (OD490nm) conducted by serum samples divided by the negative control. A ratio value higher than 2.1 was regarded as being positive. The titers were regarded as the highest dilution of antibody producing 2.1 ratio value.
Virus neutralization assay
The sera were diluted in RPMI-1640 (pH 7.2) in twofold, beginning with 1:2. PCV2 (2,000 TCID50/mL) was equally added to the diluent and incubated at 37°C for 1 h. Mixture was added into a 96-well plate with 50–70% confluent PK15 cells as the previous study (8). After 72 h, the cells were fixed with cold absolute ethanol for 15 min at 4°C, incubated with antibody to Cap, and stained with FITC-conjugated goat anti-rabbit IgG (Boshide). Titers were considered as the reciprocal of the last serum dilution at 80% or greater fluorescent focus reduction in inoculated cell cultures with a fluorescent microscope.
Lymphocyte proliferation assay
Splenocyte preparations were followed by a previous study (30). On day 28 after the first vaccination, lymphocytes were seperated aseptically from the spleens of four mice with the mouse lymphocyte separation medium (Dakewe) following the standard procedures. Lymphocytes were seeded into a 96-well plate and stimulated for 68 h with Concanavalin A (ConA; Sigma, 5 μg/mL) as positive control, Cap protein (20 μg/mL) as the stimulant, or RPMI-1640 as negative control. Then, 20 μL MTT (3-[4, 5-dimethylthiazol-2-y]-2, 5-diphenyltetrasodium bromide Tetrazolium; Sigma, 5 mg/mL in PBS) was added per well with a further incubation for 4 h. Dimethyl sulfoxide was added 100 μL per well to stop the reactions 4 h later. The OD 570nm was detected and the stimulation index (SI) was determined as SI = mean OD of Cap-stimulated cells/mean OD of unstimulated cells.
Cytokines release assay
Splenocytes were separated aseptically from the spleens of four mice from each group as mentioned above on day 28 after the first vaccination. Then, the lymphocytes were seeded into 96-well plates with 2 × 105 cells per well. Triplicate wells with RPMI-1640 alone and 2 μg Cap protein were added for 72 h, and supernatants were collected. Titers of IFN-γ and IL-4 were detected by mice IFN-γ and IL-4 ELISA Kits (ExCell Bio) following standard procedures. The concentrations of mice IFN-γ and IL-4 in the supernatants were tested by the standard curves.
PCV2 nucleic acid detection in lungs
Four mice of every group were euthanized on day 28, 35, 42, 49, and 56 after the first immunization and the lungs were collected for virus nucleic acids detection. Virus DNA was extracted and detected by a quantitative real-time PCR established previously (25). Briefly, PCV2 genomic DNA was extracted by a DNA extraction kit (Axygen). The concentrations of plasmid containing PCV2 genomic DNA were determined by Nanodrop 2000 Spectrophotometer (Thermo Scientific). The copy number was calculated using the following formula: amount (copies/mL) = 6.02 × 1023 (copies/mol) × concentration (g/μL)/MW (g/mol). The viral copy number in samples per mL was tested by real-time PCR with serially diluted plasmid from 103 to 109 copies/mL. Real-time PCR was performed with primers (forward 5′-AAGGGCTGGGTTATGGTATGT-3′ and reverse 5′-CGCTGGAGAAGGAAAAATGG-3′). Fluorescence data were recorded in real time with an IQ5 multicolor real-time PCR detection system (Bio-Rad). The viral loads were determined as the mean of the logarithmic DNA copy number divided by gram (Lg copies/gram).
Statistical analysis
All data were conducted as the mean ± standard deviation. Student's t-test and one-way ANOVA were used to assess the statistical differences among groups with SPSS 19.0 software (SPSS, Inc.). A value of p < 0.05 was regarded as significant.
Results
Identification of recombinant adenovirus vectors
In this study, pShuttle-CMV with PCV2 ORF2 gene was modified (Fig. 1). The constructed recombinant adenovirus plasmids were identified by PCR and digested with PacI. Four genes were successfully amplified with expected sizes. Two fragments of about 30 kb and 5.0 kb were observed after five recombinant plasmids were digested by PacI (Fig. 2). At 10 days posttransfection of recombinant plasmids (including the rAd-wild) into HEK293 cells, CPE were observed, indicating that the recombinant adenoviruses were packaged successfully. Then five kinds of recombinant adenoviruses were purified by plaque purification thrice and named as Ad-Cap, Ad-CD40L-Cap, Ad-Cap-GMCSF, Ad-CD40L-Cap-GMCSF, and Ad-wild. The viral titers of five kinds of recombinant adenoviruses were all about 1010 TCID50/mL.
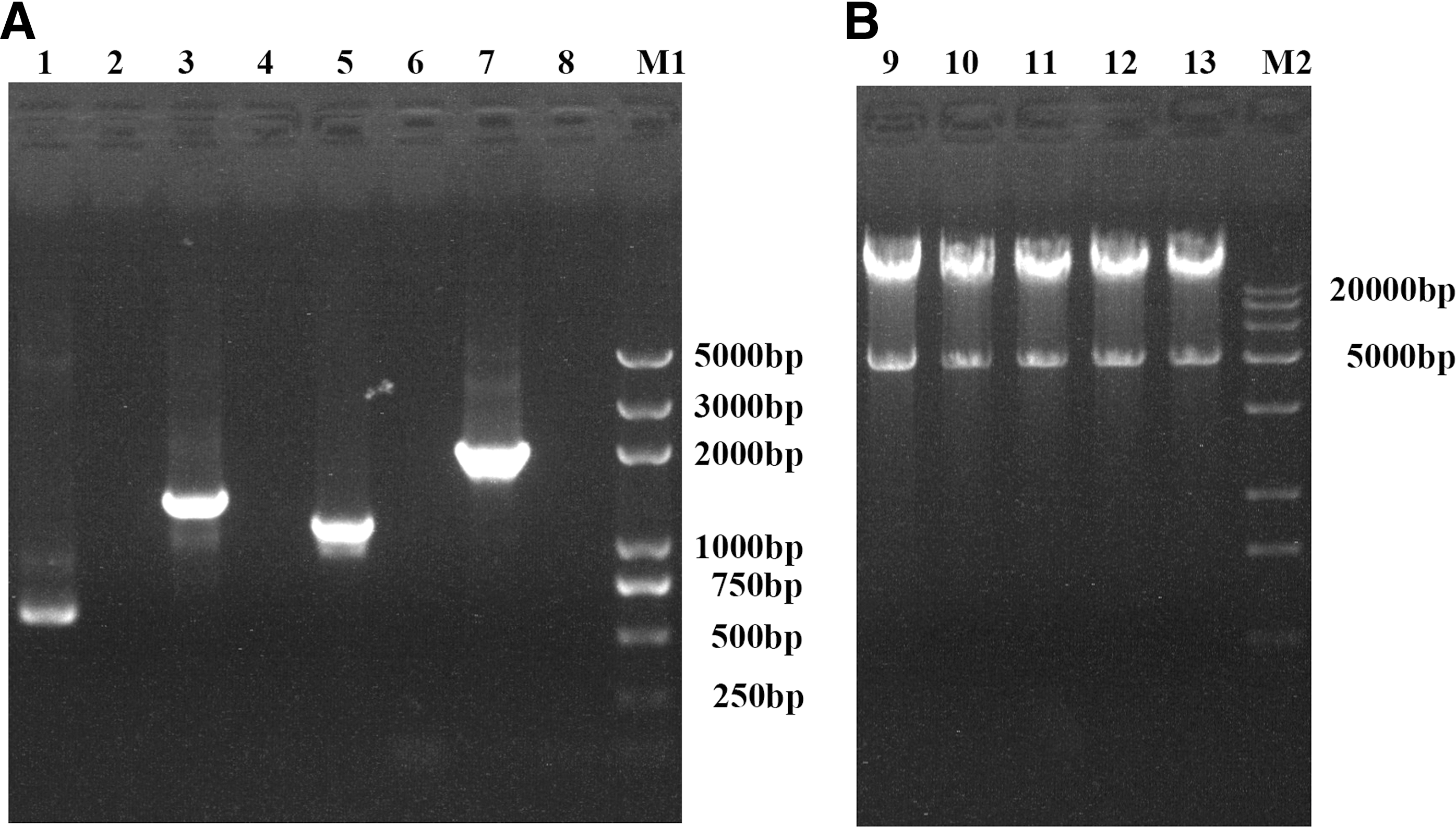
The recombinant plasmids were identified by PCR and digested with PacI enzyme.
Identification of Cap, CD40L, and GMCSF expression in recombinant adenovirus-infected cells
HEK293 cells were infected with Ad-Cap, Ad-CD40L-Cap, Ad-Cap-GMCSF, Ad-CD40L-Cap-GMCSF, and Ad-wild at 20 MOIs for 30 h. Western blot showed that Cap was detected in Ad-Cap-infected cells; CD40L and Cap were detected in Ad-CD40L-Cap-infected cells; GMCSF and Cap were detected in Ad-Cap-GMCSF-infected cells; CD40L, GMCSF, and Cap were detected in Ad-CD40L-Cap-GMCSF-infected cells, but no band appeared in Ad-wild infected or mock infection HEK293 cells (Fig. 3). Consistently, indirect IFA showed that recombinant adenovirus Ad-Cap-, Ad-CD40L-Cap-, Ad-Cap-GMCSF-, and Ad-CD40L-Cap-GMCSF-infected HEK293 cells could be stained with PCV2-specific antibody (Fig. 4A). CD40L could be detected in both Ad-CD40L-Cap- and Ad-CD40L-Cap-GMCSF-infected cells after staining with CD40L-specific antibody (Fig. 4B). GMCSF could be detected in both Ad-Cap-GMCSF- and Ad-CD40L-Cap-GMCSF-infected cells after staining with GMCSF-specific antibody (Fig. 4C). However, Cap, CD40L, and GMCSF could not be detected in Ad-wild-infected HEK293 cells or PBS-treated cells.
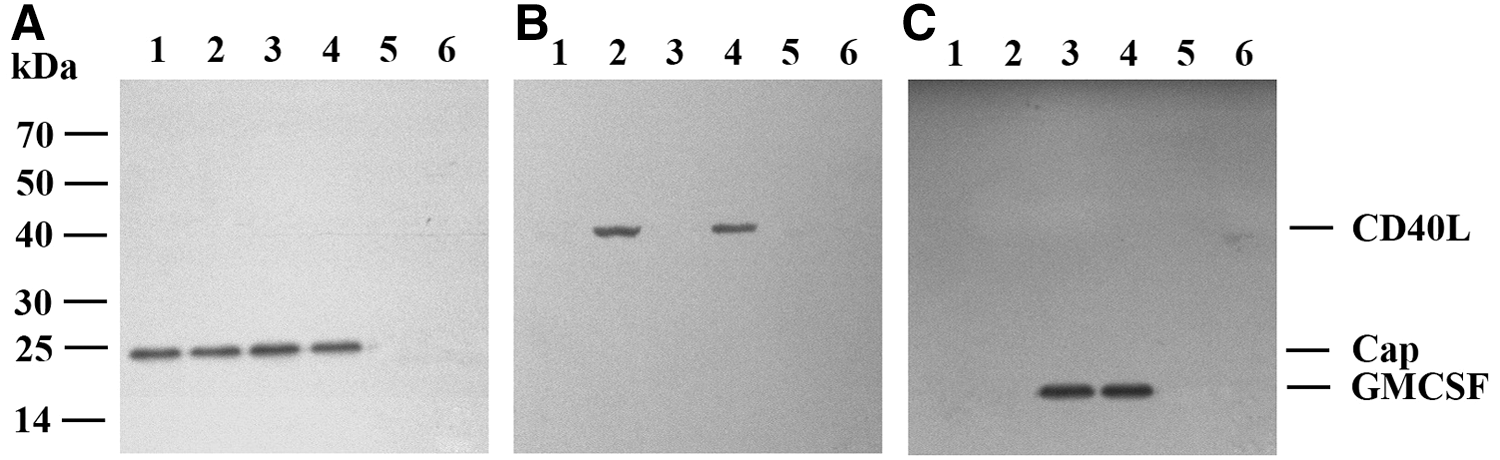
Identification of expression of Cap, CD40L, and GMCSF by Western blot. Western blot analysis of cell lysates infected with Ad-Cap (lane 1), Ad-CD40L-Cap (lane 2), Ad-Cap-GMCSF (lane 3), Ad-CD40L-Cap-GMCSF (lane 4), Ad-wild (lane 5), and phosphate-buffered saline (lane 6), respectively, by using rabbit anti-Cap serum
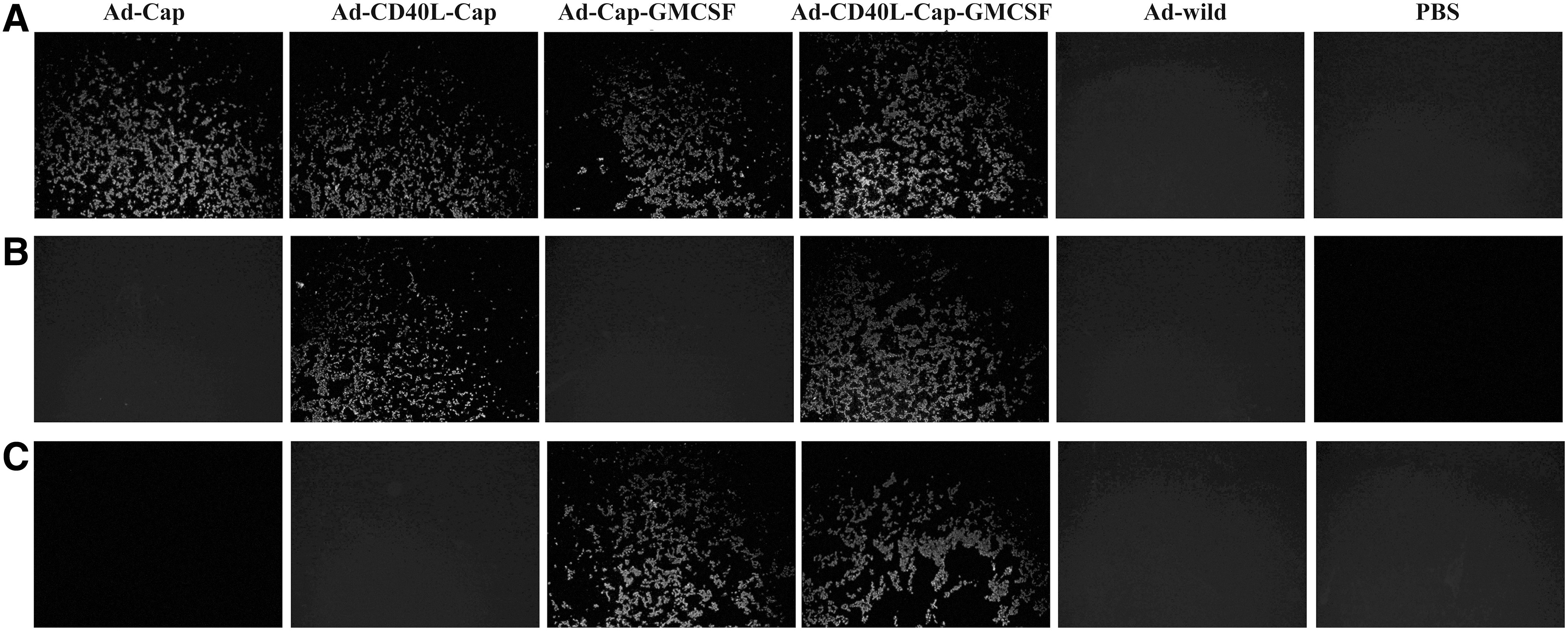
Identification of expression of Cap, CD40L, and GMCSF by immunofluorescence assay (IFA). IFA analysis of 293 cell monolayers infected with the recombinant adenoviruses by using rabbit anti-Cap serum
Humoral immune response induction by the recombinant adenoviruses
PCV2 Cap-specific antibody detection in mice
PCV2 Cap-specific antibody titers were measured in different recombinant adenovirus-vaccinated mice on 0, 7, 14, 28, 35, 42, 49, and 56 days after the first vaccination. The antibodies against PCV2 were initially detected on day 7 after the first vaccination and gradually increased in subsequent times in Ad-Cap-, Ad-CD40L-Cap-, Ad-Cap-GMCSF-, Ad-CD40L-Cap-GMCSF-vaccinated mice. Antibody titers of different immunized groups reached highest by day 35 after the first immunization, with 1:2,000, 1:4,000, 1:5,600, and 1:11,200 in mice from Ad-Cap-, Ad-CD40L-Cap-, Ad-Cap-GMCSF-, and Ad-CD40L-Cap-GMCSF-vaccinated groups, respectively. At all of detected time points after day 14, antibody titers of Ad-CD40L-Cap-GMCSF-vaccinated group were highest among these groups, and significantly higher compared with the Ad-CD40L-Cap-vaccinated group and Ad-Cap-GMCSF-vaccinated group (p < 0.05), Ad-Cap-GMCSF-vaccinated group was slightly higher than the Ad-CD40L-Cap-vaccinated group, but did not show significant difference (p > 0.05). Mice vaccinated with Ad-wild or PBS did not detect Cap-specific antibody throughout the whole experiments (Fig. 5). In addition, the antibody titers did not show significant difference among four mice per group at all indicated times.
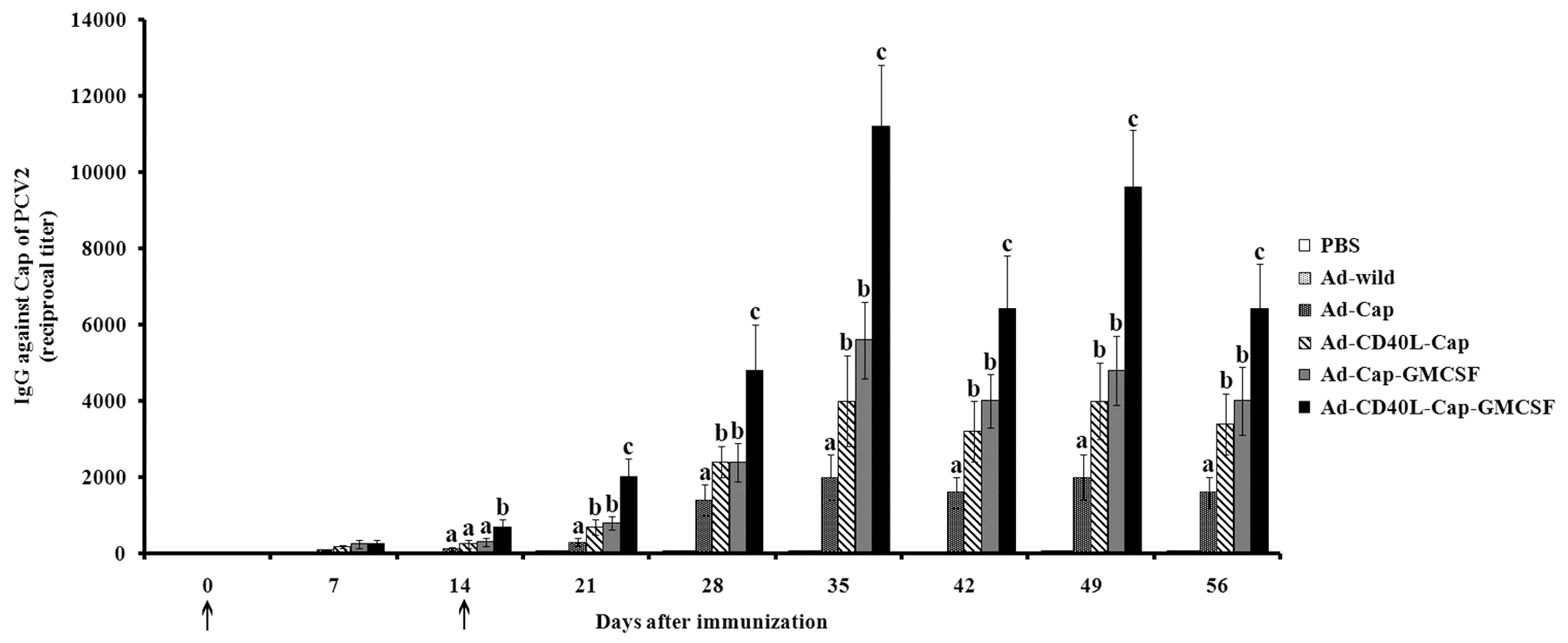
Indirect ELISA for porcine circovirus 2 (PCV2)-specific antibodies of sera from mice immunized with different adenoviruses. The results were expressed as the ratio of OD490nm produced by the serum samples compared to the negative control serum. Sera giving a ratio value higher than 2.1 were considered to be positive sera. The titers were expressed as the highest dilution of antibody producing a 2.1 ratio value. Data are presented as the mean ± standard deviation (SD). Treatments with different lowercase letters (a, b, and c) indicate statistically significant differences in specific antibody levels between the indicated groups (p < 0.05). Arrows (↑) indicate time of initial immunization and boost.
PCV2 Cap-neutralizing antibody detection in mice
Neutralizing antibody titers of sera from inoculated groups were measured in PK15 cells that were challenged with PCV2. The sera from mice vaccinated with recombinant adenoviruses could prevent PK15 cells from PCV2 infection. On day 35 after the first vaccination, neutralizing antibodies titers reached 1:40, 1:96, 1:112, and 1:224 in Ad-Cap-, Ad-CD40L-Cap-, Ad-Cap-GMCSF-, and Ad-CD40L-Cap-GMCSF-vaccinated groups, respectively, whereas sera from Ad-wild-vaccinated mice or PBS-treated mice did not show a neutralizing activity (Fig. 6). On day 35 after the first vaccination, the neutralizing antibody titer of Ad-CD40L-Cap-GMCSF-vaccinated group was highest among these groups (p < 0.05), whereas Ad-Cap-GMCSF-vaccinated group was not significantly higher than Ad-CD40L-Cap-vaccinated group (p > 0.05).
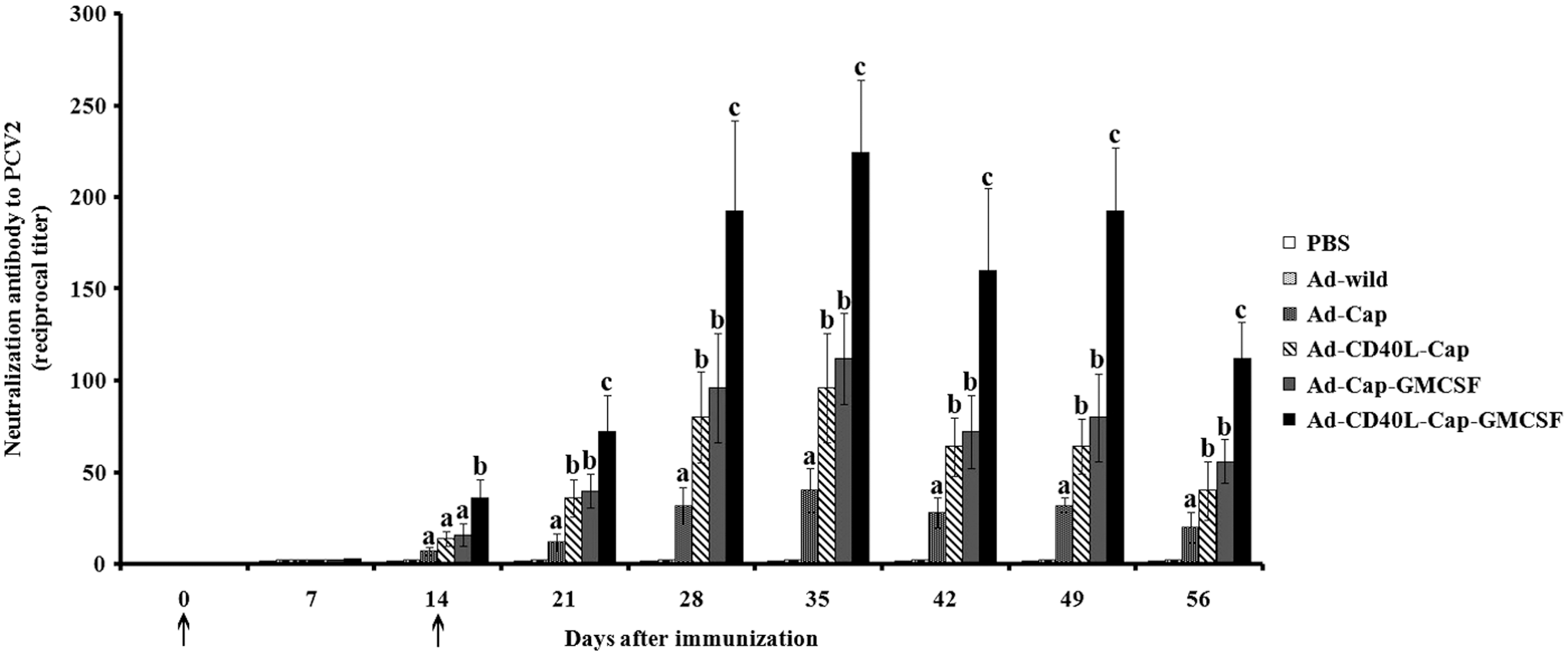
IFA assay for neutralization activity of sera from the mice immunized with different adenoviruses. Neutralizing titers are reciprocals of the highest dilution of serum, which yielded 80% reduction in virus infectivity. Data are presented as the mean ± SD. Treatments with different lowercase letters (a, b, and c) indicate statistically significant differences in neutralizing antibody levels between the indicated groups (p < 0.05). Arrows (↑) indicate time of initial immunization and boost.
Cellular immune response induction by the recombinant adenoviruses
Lymphocyte proliferative activity
Lymphocyte proliferation level is one of the most important indexes of cellular immune response, and lymphocyte proliferative activities of different vaccinated mice were measured on day 28 after the first immunization. As shown in Figure 7, the proliferative level of Ad-Cap-, Ad-CD40L-Cap-, Ad-Cap-GMCSF-, and Ad-CD40L-Cap-GMCSF-vaccinated mice were more compared with Ad-wild-vaccinated mice or PBS-treated mice. Ad-CD40L-Cap-GMCSF-vaccinated group showed higher proliferative level than the other groups (p < 0.05), the proliferative level of Ad-Cap-GMCSF-vaccinated group was not significantly higher compared with the Ad-CD40L-Cap-vaccinated group (p > 0.05), but both were higher than the Ad-Cap-vaccinated group (p < 0.05).

Cell proliferation levels of splenocytes after specific stimulation. After immunization with different recombinant adenoviruses, four mice of each group were sacrificed and splenocytes were pooled for the lymphocyte proliferation assay. Data are presented as the mean ± SD of three replicates of the same splenocyte pool. Treatments with different lowercase letters (a, b, c and d) indicate statistically significant differences in stimulation index between the indicated groups (p < 0.05).
Th1-type and Th2-type cytokine detection
Another important index of cellular immune response was cytokine secretion by Th1 and Th2 lymphocytes. On day 28 after the first immunization, the secretion of IFN-γ and IL-4 was detected in the stimulated lymphocytes to evaluate the Th1 and Th2 response of immunized mice. IFN-γ and IL-4 production of stimulated lymphocytes from Ad-Cap-, Ad-CD40L-Cap-, Ad-Cap-GMCSF-, and Ad-CD40L-Cap-GMCSF-vaccinated mice were higher compared with Ad-wild-vaccinated mice or PBS-treated mice. In recombinant adenovirus-vaccinated groups, mice inoculated with adenovirus Ad-CD40L-Cap-GMCSF showed significantly higher IFN-γ and IL-4 level than others (p < 0.05). Besides, cytokines levels of Ad-Cap-GMCSF-vaccinated group were also higher than the Ad-CD40L-Cap-vaccinated group (p > 0.05) and both were higher than the Ad-Cap vaccinated group (p < 0.05) (Fig. 8).
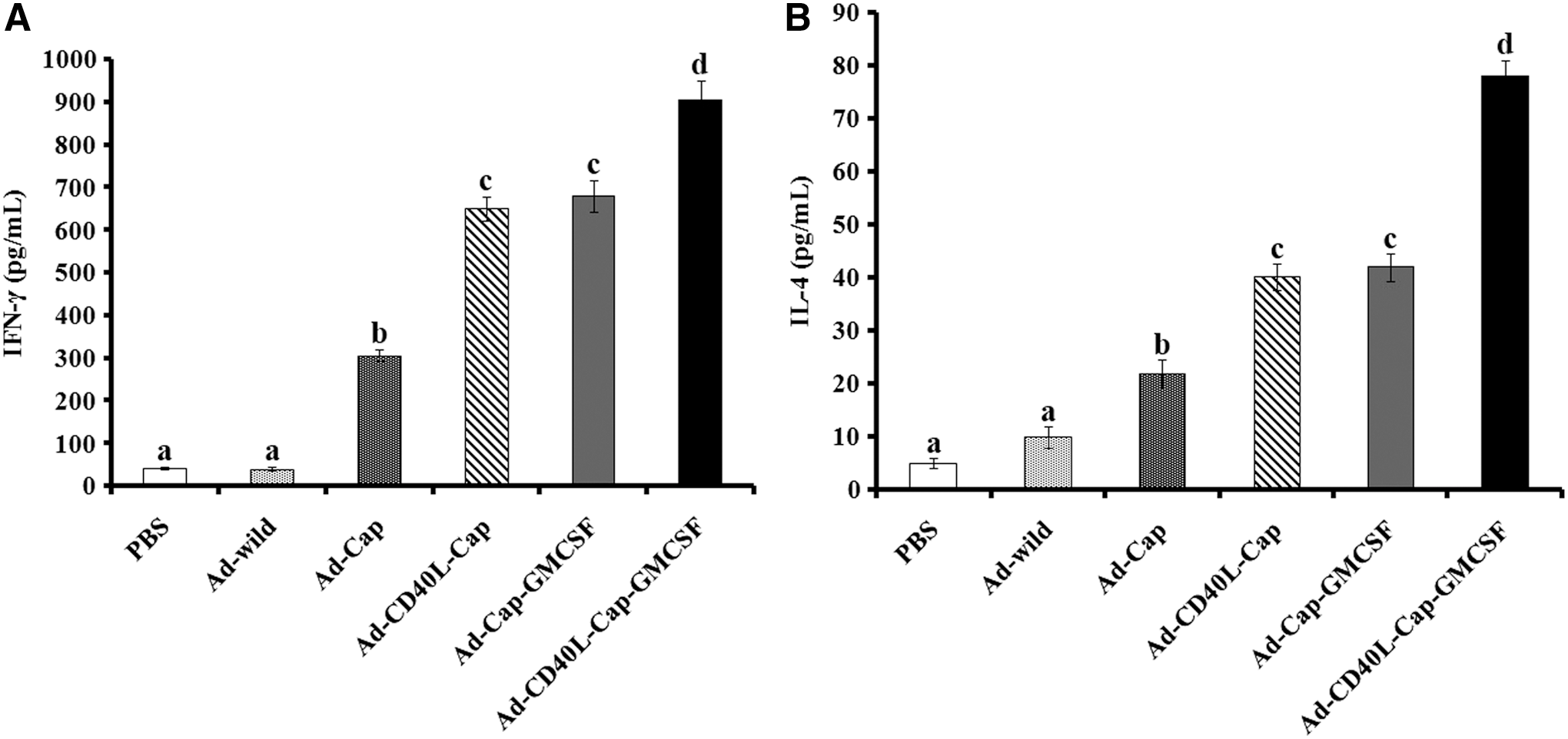
Levels of
Protective effects of the recombinant adenoviruses against PCV2 challenge
Virus DNA extracted from the lungs of six groups was detected by quantitative PCR on day 28, 35, 42, 49, and 56 after the first immunization. As shown in Figure 9, viral loads were undetectable in the lungs collected on day 28 after the first vaccination. After day 35, the viral loads in the lungs of Ad-wild or PBS-treated groups were significantly higher than modified recombinant adenovirus-vaccinated groups at all detected time points (p < 0.05). Notably, Ad-CD40L-Cap-GMCSF-vaccinated group showed significantly lower viral loads compared to the Ad-Cap groups (p < 0.05), Ad-CD40L-Cap and Ad-Cap-GMCSF groups, on day 35, 42, 49, and 56 after the first immunization (p < 0.05). The number of PCV2 nucleic acid of Ad-CD40L-Cap group was not significantly different from that of Ad-Cap-GMCSF group (p > 0.05), but both Ad-CD40L-Cap and Ad-Cap-GMCSF groups showed lower viral loads than the Ad-Cap group (p < 0.05) on day 35, 42, 49, and 56 after the first immunization.
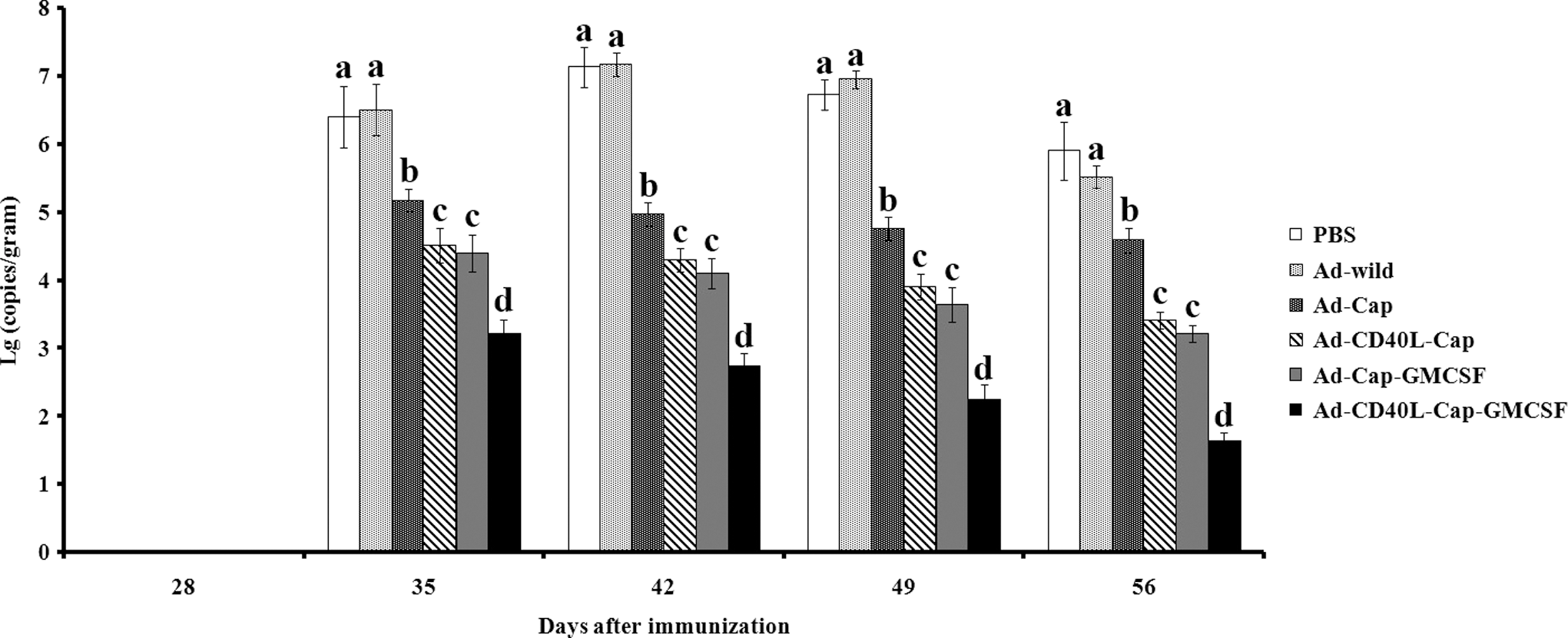
Quantification of PCV2 viral loads in lungs from challenged mice. Mice were challenged with 2 × 104 tissue culture infectious dose 50 (TCID50) PCV2 on day 28 after first immunization, and PCV2 viral loads were detected by quantitative PCR in lung samples collected from mice on day 28, 35, 42, 49, and 56 after first vaccination. Data are presented as the mean ± SD. Treatments with different lowercase letters (a, b, c, and d) indicate statistically significant differences in virus loads between the indicated groups (p < 0.05).
Discussion
Recombinant adenovirus (Ad) vector is one of the most widely used live virus vectors with many advantages such as high transduction efficiency and wide tropism (6,50). Adenovirus vector system has been used to express many proteins, including PCV2, porcine reproductive and respiratory syndrome virus, and porcine parvovirus (26,42,52). However, adenoviruses expressing these antigens have not been put into practice in field and no relevant adenovirus commercial vaccines have been available yet. The main reason is that many proteins of adenovirus itself could induce strong immune responses, resulting in poor efficacy (55). To solve this problem, several studies have been conducted, such as coexpressing cytokines (33,53). So in the present study, porcine CD40L and GMCSF were applied as immune adjuvants and inserted into the adenovirus to enhance the immune responses and protective efficacy of PCV2 adenovirus vaccine.
CD40L is mainly expressed by activated CD4+ T cells and the interaction of CD40L and its receptor, CD40, plays important roles in both humoral and cellular immune responses, such as the Th1-type response development (3,11), Ig type switching, antibody production (10,41), and interactions between T cell and DC or T cell and macrophage (56). Meanwhile, CD40L/CD40 interaction could induce activated B cells to upexpress CD95, making activated B cells more sensitive to apoptosis signal pathways (7). However, most importantly, CD40L has a powerful immunostimulatory effect on increasing humoral and cellular immune responses. Thus, it is an effective vaccination strategy applied in vaccine designs of some viruses, such as HIV (22), bovine herpesvirus-1 (35,36), and influenza virus (23). In the study, CD40L and Cap were inserted into the transfer vector pShuttle-CMV and constructed recombinant adenovirus, named as Ad-CD40L-Cap. The results showed that the Ad-CD40L-Cap-vaccinated group did induce stronger humoral and cellular immune responses in mice than Ad-Cap, indicating that CD40L could enhance the immunogenicity of PCV2 adenovirus vaccines in mice.
In addition to CD40L, GMCSF is another cytokine used to enhance immune responses in the vaccine development. GMCSF can stimulate neutrophil, monocyte/macrophage, eosinophil colony formation, and enhance antibody-dependent cell-mediated cytotoxicity (38,39). Although some studies suggest that GMCSF increase causes inflammatory response (16,40), and others suggest that GMCSF promotes immunological tolerance by acting as an immunoregulatory cytokine (31,43), it is still a very promising molecular adjuvant to increase vaccine efficacy (48). In this study, we constructed the recombinant adenovirus, Ad-Cap-GMCSF, and evaluated the role of GMCSF in increasing immunogenicity of adenovirus vaccines in mice. The results showed that Ad-Cap-GMCSF significantly increased humoral and cellular immune responses to PCV2 in mice compared to Ad-Cap.
DC is mainly responsible for ingestion, processing, and presentation of antigen, resulting in specific immune response activation (4). GMCSF can stimulate the proliferation and differentiation of DC, while CD40L can stimulate B-cell activation, differentiation, and promote humoral immune responses. Therefore, these two cytokines can work together theoretically to enhance the immune response of vaccines. In this study, mice experiments showed that Ad-CD40L-Cap-GMCSF elicited the highest immune responses compared to Ad-CD40L-Cap, Ad-Cap-GMCSF, or Ad-Cap, which was consistent with the predication, suggesting that these two cytokines, CD40L and GMCSF, not only increased immunogenicity of adenovirus vaccines individually but also could produce a synergistic effect in improving the efficacy of adenovirus system.
To confirm the protective immune response induced by different recombinant adenoviruses, the mice were challenged with PCV2. Virus loads in lung samples were detected to evaluate the protective efficiency of different recombinant adenoviruses. The results showed that Ad-CD40L-Cap-GMCSF, Ad-CD40L-Cap, and Ad-Cap-GMCSF could provide higher protective efficiency than Ad-Cap, and Ad-CD40L-Cap-GMCSF showed highest among the groups, indicated that better protection was developed by the Ad-CD40L-Cap-GMCSF group. Moreover, vaccine efficacy between the recombinant adenoviruses constructed in this study and commercial vaccines should be tested in pigs. Although no commercial adenovirus vaccines against PCV2 have been available so far, we wish that CD40L and GMCSF will be beneficial to improve immune responses and protective efficacy of adenovirus vaccines against PCV2.
In conclusion, CD40L and GMCSF could synergistically increase both humoral and cellular immune responses of PCV2 adenovirus vaccine, and could protect mice against the PCV2 challenge. Although there are great anatomical and physiological differences between mice and pigs, the results of mice experiment would be helpful for experiment design to test the vaccine efficacy of the constructed recombinant adenoviruses in pig studies. According to the results in this study, we have optimized the parameters, such as the dose and route of delivery and the procedure of vaccination, to improve the efficacy of PCV2 adenovirus vaccines in pigs. Now the relative works are under way in pigs.
Footnotes
Acknowledgments
This research was supported by Key project of Shaanxi province science and technology innovation team (2013KCT-28) and the Fundamental Research Funds for the Central Universities (ZD2013009, Z111021103).
Author Disclosure Statement
No competing financial interests exist.