
Abstract
Intestinal epithelial cells, which serve as the first physical barrier to protect intestinal tract from external antigens, have an important role in the local innate immunity. Screening of reference genes that have stable expression levels after viral infection in porcine intestinal epithelial cells is critical for ensuring the reliability of the expression analysis on anti-infection genes in porcine intestinal epithelial cells. In this study, nine common reference genes in pigs, including ACTB, B2M, GAPDH, HMBS, SDHA, HPRT1, TBP, YWHAZ, and RPL32, were chosen as the candidate reference genes. Porcine sapelovirus (PSV) was used as a model virus to infect porcine intestinal epithelial cell line (IPEC-J2). The expression stability of the nine genes was assessed by the geNorm, NormFinder, and BestKeeper software. Moreover, RefFinder program was used to evaluate the analytical results of above three softwares, and a relative expression experiment of selected target gene was used to verify the analysis results. The comprehensive results indicated that the gene combination of TBP and RPL32 has the most stable expression, which could be considered as an appropriate reference gene for research on gene expression after PSV infection in IPEC-J2cells. The results provided essential data for expression analysis of anti-infection genes in porcine intestinal epithelial cells.
Introduction
I
Pigs are economically important animals, which serve as a major source of animal proteins for humans. However, infectious diseases caused by porcine intestinal viruses are a grave threat to the healthy development of the pig industry and lead to significant economic losses. The porcine sapelovirus (PSV), also called porcine enterovirus 8, can induce viral syndromes in multiple systems, including porcine diarrhea, pneumonia, severe or mild nerve diseases, and dysgenesis. In recent years, PSV has received increasing attention because of the high infection ratio in pig herds farmed intensively; PSV has also been considered as a viral model for research on the antiviral innate immunity of the intestinal mucosa (18).
The porcine intestinal epithelial cell line (IPEC-J2) is an epithelial cell line that has been developed in recent years (3). This cell line originates from nontransformed and nononcogenic intestinal epithelial cells in the jejunum of newborn piglets, which had not yet been breast-fed. The cells retain the differentiation characteristic of primary intestinal epithelial cells and are very similar to primitive intestinal epithelial cells. IPEC-J2 has been extensively applied in studies on the interaction of intestinal bacteria, probiotics, and enteroviruses with the intestinal mucosa (3). Previous research has estimated the stability of reference genes in various porcine tissues, but mostly focused on specific tissues, such as in the liver (14), longissimus dorsi (11), stomach (27), fat (33), alveolar macrophages (5), embryo (21), and mesenchymal stem cells (24). However, the selection of ideal reference genes has not been reported in IPEC-J2, which is a satisfactory in-vitro model for studying the local immunity of intestinal mucosa (30). Therefore, we used the geNorm, NormFinder, and BestKeeper software to assess the expression stability of nine reference genes, including ACTB, B2M, GAPDH, HMBS, SDHA, HPRT1, TBP, YWHAZ, and RPL32, after PSV infection in IPEC-J2cells. Moreover, the web-based analysis tool RefFinder was used to evaluate the analytical results of above three softwares and an experimental analysis of relative expression of selected target gene was used to conduct a general verification of the reference gene candidates. This study provides a foundation for research on gene expression levels after viral infection in porcine intestinal epithelial cells.
Materials and Methods
Experimental cells, virus strains, and sample collection
The IPEC-J2 cells and PSV strains were preserved and provided by our laboratory (33). The IPEC-J2 cell suspension was inoculated on six-well cell culture plates at a rate of 2.5 mL/well and cultured in an incubator with 5% CO2 at 37°C. When the cell confluence reached 80–90%, PSVs were inoculated at the rate of 2 mL/well (multiplicity of infection [MOI] = 2), and the cell culture was continued in the 5% CO2 incubator at 37°C for 1 h. Three wells in each plate were inoculated with viruses, whereas the other three blank controls were inoculated with the same volume of the maintenance medium containing 1% fetal calf serum. After the inocula were aspirated out of each well, the RPMI-1640 culture medium containing 1% fetal calf serum was added at the rate of 2.5 mL/well and cell culture continued in the 5% CO2 incubator at 37°C. The appearance of the cells in each plate was observed, and the culture plate was collected at 12, 36, 60, and 84 h after the PSV infection. To each well of the collected plate, 800 μL of TRIzol was added and then preserved at −80°C for use.
Extraction of total RNA and complementary DNA synthesis
The RNA of cell lysate collected at each infected time point was extracted using TRIzol reagent (Life Technology) according to the manufacturer's instruction. The complementary DNA (cDNA) synthesis was conducted according to the instructions of the Reverse Transcription Kit (Takara), and the products were preserved at −20°C for use.
Selection of reference genes and preparation of positive standards
According to the literature, nine common reference genes in pigs, including ACTB, B2M, GAPDH, HMBS, SDHA, HPRT1, TBP, YWHAZ, and RPL32, were chosen as reference gene candidates in this research. The primers for the nine common reference genes were previously reported in literature (23), as listed in Table 1.
The preparation of positive standards of the reference genes is as below: with the cDNA synthesized in the previous section as the template, conventional polymerase chain reaction (PCR) was conducted with the abovementioned primers. The 25 μL amplification system was composed of 12.5 μL of 2× Taq Master Mix, 9.5 μL of dH2O, 1 μL of forward primer, 1 μL of reverse primer, and 1 μL of template. The reaction procedure was as follows: predenaturation at 94°C for 5 min, 30 cycles of denaturation at 94°C for 30 sec, annealing at 60°C for 30 sec, and extension at 72°C for 45 sec; and final extension at 72°C for 5 min. The PCR product was identified by 2% agarose gel electrophoresis, recycled, and purified with the Axygen DNA Gel Extraction Kit (Axygen). The PCR product and pMD19-T vector were ligated, transformed into DH5α competent cells, and coated on Amp+ agar plates. After cultivation at 37°C, single colonies were selected for bacterial liquid PCR identification. The recombinant plasmids were sent to Invitrogen for sequencing, and the plasmids contained qualified sequences used as positive standards.
Quantitative real time reverse transcription PCR
Reactions were conducted on the BIO-RAD CFX96 real-time PCR detection system (Bio-Rad). The 15 μL PCR reaction system was composed of 7.5 μL of SYBR Premix Ex Taq™ II, 0.5 μL of forward primer, 0.5 μL of reverse primer, 1 μL of template, and 5.5 μL of dH2O. The PCR amplification procedure was as follows: predenaturation at 95°C for 30 sec; 39 cycles of denaturation at 95°C for 5 sec, annealing at 60°C for 25 sec, and extension at 72°C for 20 sec; and melting curve analysis. Three replicates were arranged for each sample, and a negative control with dH2O as the template was added.
Stability evaluation of the reference genes
The stability of all the chosen reference genes was analyzed with geNorm, Normfinder, and BestKeeper three softwares. Moreover, comprehensive evaluation of the analytical results of above three software programs and verification of the reference gene candidates were conducted with the RefFinder program (
Experiment verification of the screened reference gene
To ensure the reliability of the analysis results, B2M, YWHAZ, TBP, and RPL32 were, respectively, used as the standardized index to correct quantitative data. The 2−ΔΔCt method was adopted to calculate the relative expression of NLRC5 (NLR family, CARD domain containing 5) to verify the stability of reference genes. NLRC5 is a newly found member of NLRs family (7), which has been verified to have significant regulatory effect on the Type I IFN production pathway and an important role in antiviral innate immunity (6,16,20,25,32). Previous research found that after PSV infection in porcine intestinal epithelial cells, the expression of NLRC5 significantly increased, and NLRC5 participated in the innate immune response of porcine intestinal epithelial cells to PSV (18). Therefore, NLRC5 was selected as the target gene in this research.
Results
PCR electrophoretic detection and melting curve analysis of reference genes
With the IPEC-J2 cDNA as the template, the target fragments of reference gene candidates were, respectively, amplified. The electrophoretic detection result showed that the target band was single, and the fragment size was consistent with the expected result.
Primer specificity could be determined by the analysis of the real-time PCR melting curve and amplification curve. As shown in Supplementary Figure S1 (Supplementary Data are available online at
Expression stability of reference genes
The expression stability and pairwise variation of each candidate reference gene analyzed by three softwares is shown in Figure 1. The analytical results of geNorm (Fig. 1A) indicated that the reference genes with the most stable expression were ACTB and TBP, followed by the RPL32 and GAPDH, whereas B2M was the most unstable gene, whose M value exceeded 1.5. The NormFinder analysis results (Fig. 1B) indicated that the expression of GAPDH was most stable, the expression levels of RPL32 and HPRT1 were relatively stable, and B2M was the most unstable one. According to BestKeeper analysis (Fig. 1C), TBP was the reference gene with the most stable expression, the expression levels of ACTB, RPL32, and HMBS were relatively stable, and B2M was still the most unstable one. In addition, in the pairwise variation analysis by geNorm software (Fig. 1D), the Vn/n + 1 value was much larger than the default 0.15, thereby indicating that the expression stability of single reference gene was not strong. According to the geNorm software's operation manual, two to three of the most stable reference genes can be selected according to tendency of V value. As shown in Figure 1D, V3/4 = 0.255 > V2/3 = 0.198, thereby suggesting that the addition of the third gene did not increase the stability of the combination. Therefore, the reference gene combination with two genes was the most stable.
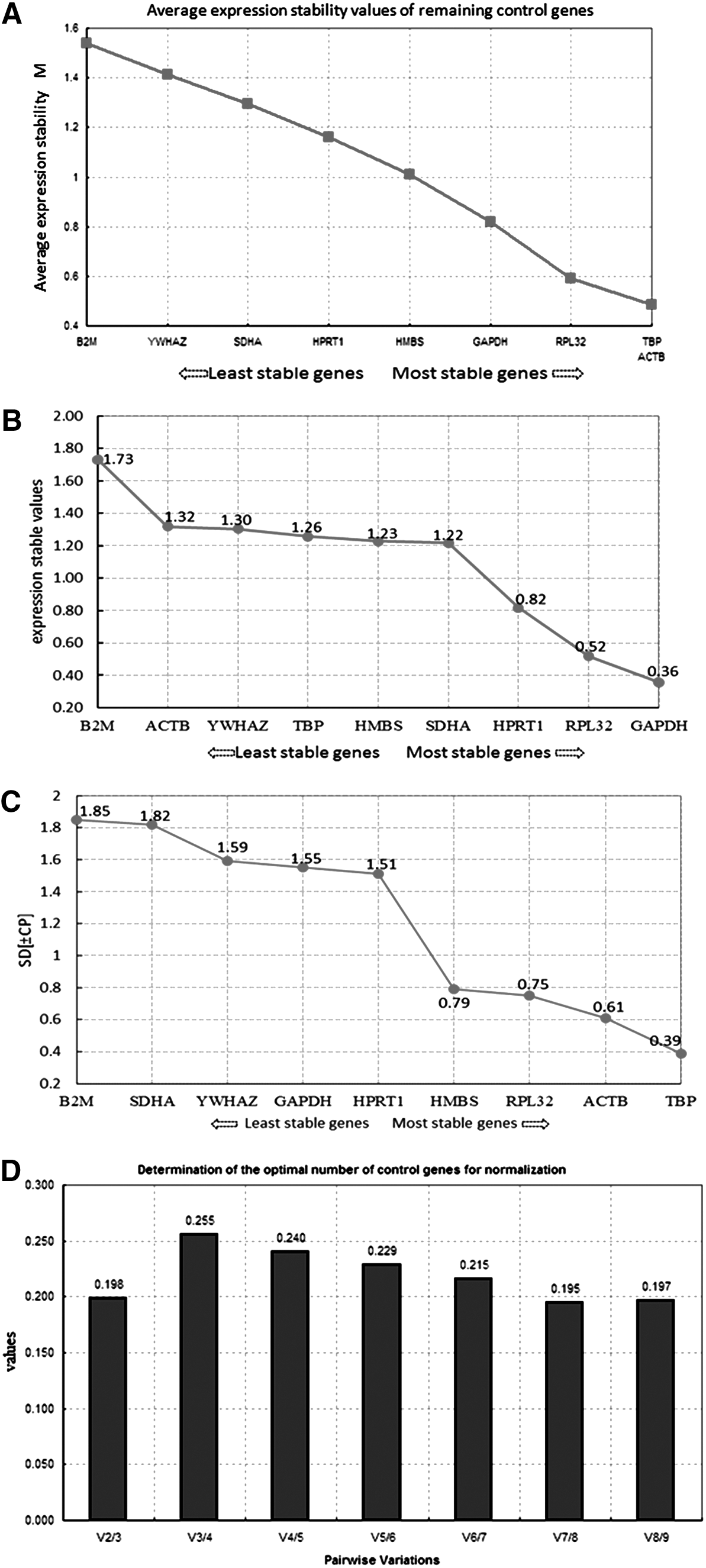
Expression stability and pairwise variation coefficient of each reference gene.
Furthermore, the analytical results of above three software programs were further evaluated comprehensively by the RefFinder program. The evaluation result showed that the analytical results obtained from RefFinder (Table 2) were generally consistent with the separately calculated results of the three softwares. Therefore, the analytical results of the three kinds of software were correct. The comprehensive ranking showed that the reference genes with the most stable expression were TBP and RPL32, followed by GAPDH, HPRT1, HMBS, and SDHA. The expression levels of YWHAZ and B2M were the most unstable. By taking the pairwise variation result into consideration, TBP and RPL32 were combined as the reference gene in the analysis of gene expression after PSV infection in IPEC-J2 cells.
Experiment verification of the selected reference gene
The relative expression levels of NLRC5 were calculated with YWHAZ, B2M, TBP, and RPL32 as the reference genes (Fig. 2). For the relative expression of NLRC5 at 12, 36, 72, and 84 h, the individual results of YWHAZ or B2M were lower than those with TBP and RPL32 when simultaneously used as reference genes. A significantly higher NLRC5 expression at 36 h was obtained in the analysis by TBP and RPL32 than by YWHAZ and B2M. In addition, the express curve calculated by TBP and RPL32 showed an express trend close to the previous microarray analysis results (28), but the express curve calculated by YWHAZ and B2M was messy, especially the express curve of B2M was almost like a straight line, thereby suggesting that the TBP and RPL32 had better stability. The results provided an experiment evidence to prove that TBP and RPL32 were the appropriate reference genes that could be used in the analysis of gene expression after PSV infection in IPEC-J2 cells.
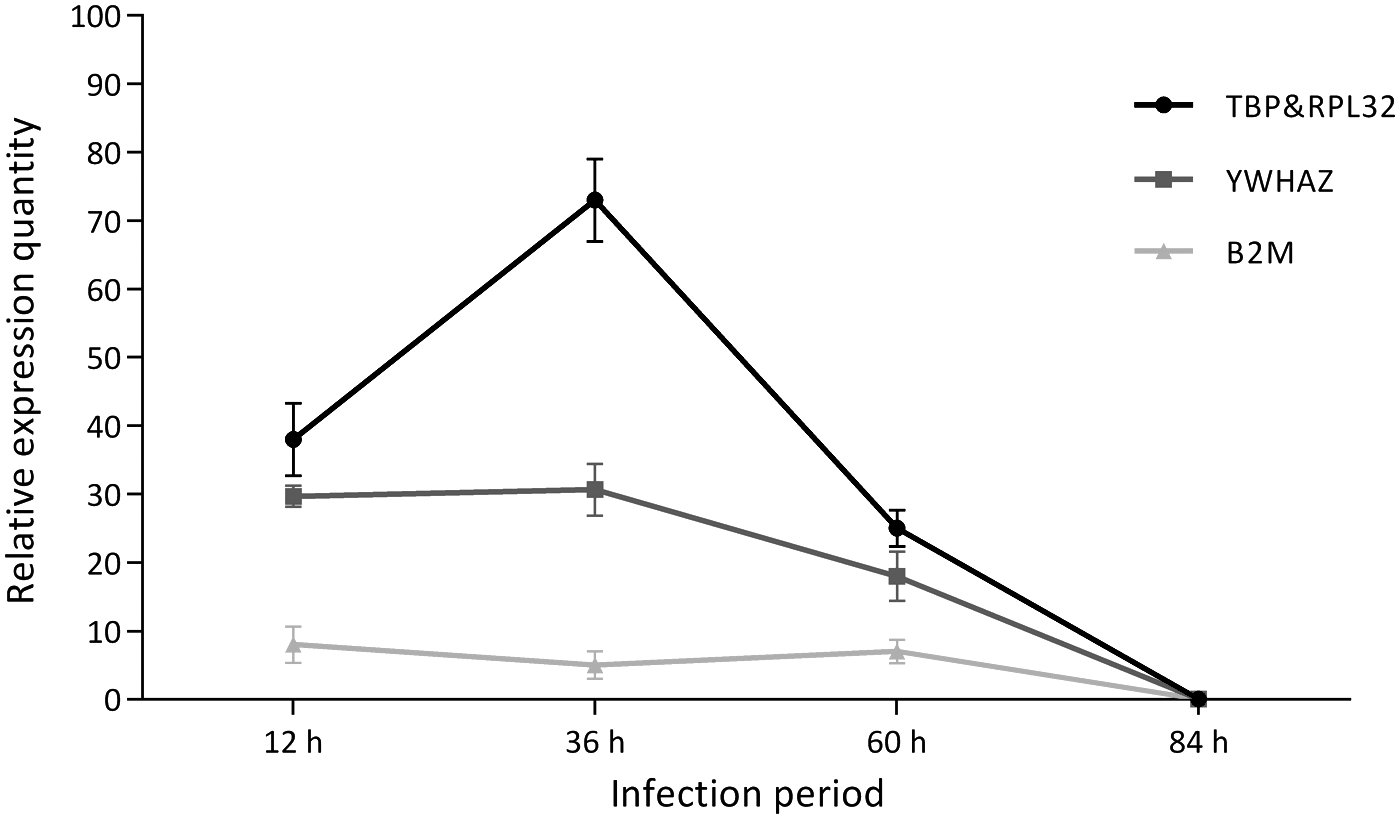
Relative expression of NLRC5.
Discussion
In research on gene expression in various tissues and cells, ideal reference genes are expected to exhibit stable expression and not be affected by experimental conditions, such as the feeding condition, external treatment, development phase, or physiological status. However, several studies have indicated the absence of the reference gene of universal expression stability in various experimental conditions (4,17). According to existing literature, GAPDH and ACTB are two commonly used reference genes in pig research (12). However, the stability of GAPDH and ACTB significantly vary; in some cases, the use of these genes is inappropriate for the correction of gene expression analysis (1,15,28,29). The random selection of reference genes without verification is inadvisable. In this study, we first screened the ideal reference genes in IPEC-J2 cells after PSV infection, which provided a basis for further expression analysis of anti-infection genes in porcine intestinal epithelial cells.
Among the analytical results of three softwares, the results analyzed by geNorm and NormFinder software were basically identical, which indicated that the expression levels of ACTB and TBP were the most stable, while that of B2M was the most unstable after PSV infection in IPEC-J2 cells. This result is partially consistent with the report by Nygard et al. (26), wherein the expression levels of ACTB, RPL4, TBP, and HPRT1 were the most stable in various porcine tissues. Martinez-Giner et al. (23) screened the reference genes of uterine and ovarian tissues at different gestation stages of sows, and their results also indicated that TBP had the most stable expression; HMBS and B2M had the most unstable expressions in uterine tissues, and ACTB in ovarian tissues. This trend is partially consistent with the results of our research. In addition, Erkens et al. (8) also reported that the reference gene combination with the most stable expression in porcine longissimus dorsi was composed of ACTB, TBP, and TOP2β. However, an analysis of the expression stability of reference genes in various porcine tissues by Tai Yulei et al. (31) indicated that the expression of ACTB was the most unstable. When screening for reference genes in piglet peripheral blood, Wang et al. (34) also found that ACTB was the gene with the most unstable expression, whereas B2M exhibited the most stable expression. These results were completely opposite to the results of the present study, which suggest that the expression stability of reference genes is very different in different tissues and conditions.
Interestingly, NormFinder analysis showed that GAPDH was the reference gene with the most stable expression, which was distinctly different from the analytical results of above two softwares. The respective stability results of geNorm, Normfinder, and BestKeeper software were not completely identical, which might be attributed to the different calculation methods. GeNorm ranks the genes according to the similarities obtained in the pairwise comparisons of all reference genes. NormFinder is based on intraclass variance and interclass variance. The calculation principle of BestKeeper is similar to that of geNorm, but its pairwise correlation analysis directly relies on the Cq value, which is the most direct reflection of original data. To increase the reliability of analysis results, more than one algorithm should be applied in the evaluation of reference genes because no perfect algorithm exists. Therefore, RefFinder program is recommended to comprehensively evaluate the analytical results of above three softwares and verify the reference gene candidates, thereby avoiding the possible one-sidedness caused by the use of a single method while providing quick and simple operation.
In addition, the pairwise variation result of GeNorm analysis indicated that the Vn/n + 1 values were far larger than the default 0.15. According to the geNorm software manual, V7/8 with the lowest V value should be selected, that is, seven reference genes are combined to adjust the quantitative expression analysis. However, the use of so many reference genes in an experimental process is impractical. Besides, several reports have suggested that a combination of three genes with the most stable expression is most appropriate to adjust data. However, V3/4 > V2/3 in the present study, which means that the stability of the combination would not increase after a third gene was added. Therefore, a combination of two reference genes is sufficient for the analysis of gene expression in this study. According to the RefFinder comprehensive ranking, TBP, RPL32, and GAPDH were the reference genes with the most stable expression. However, GAPDH cannot be used as an appropriate reference gene in gene expression analysis, because its standard deviation value was larger than 1 in the BestKeeper analysis. Meanwhile, when YWHAZ, B2M, TBP, and RPL32 were separately used as reference genes in the experimental analysis of relative expression of NLRC5 to verify the analysis results, TBP and RPL32 were again identified as the most stable reference genes. Therefore, TBP and RPL32 are recommended as the standardized index for adjusting quantitative data of gene expression analysis after PSV infection in IPEC-J2 cells.
In conclusion, this study successfully screened a reference gene combination, including TBP and RPL32 in IPEC-J2 cells after PSV infection, and provided essential data for the further study of gene expression after viral infection in porcine intestinal epithelial cells.
Footnotes
Acknowledgment
This study was funded by Natural Science Foundation of China (NSFC; Grant No. 31302074).
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.