
Abstract
In children at risk for type 1 diabetes, innate immune activity is detected before seroconversion. Enterovirus infections have been linked to diabetes development, and a polymorphism (A946T) in the innate immune sensor recognizing enterovirus RNA, interferon-induced with helicase C domain 1/melanoma differentiation-associated protein 5, predisposes to disease. We hypothesized that the strength of innate antienteroviral responses is affected in autoimmune type 1 diabetes patients and linked to the A946T polymorphism. We compared induction of interferon-stimulated genes (ISGs) in peripheral blood mononuclear cells (PBMCs) and dendritic cells (DCs) in healthy individuals and diabetes patients upon stimulation with enterovirus, enterovirus-antibody complexes, or ligands mimicking infection in relation to the A946T polymorphism. Overall, PBMCs of diabetes patients and healthy donors showed comparable ISG induction upon stimulation. No differences were observed in DCs. Interestingly, the data imply that the magnitude of responses to enterovirus and enterovirus-antibody complexes in PBMCs is critically influenced by the A946T polymorphism and elevated in heterozygotes compared to TT homozygous individuals in autoimmune diabetes patients, but not healthy controls. These data imply an intrinsic difference in the responses to enterovirus and enterovirus-antibody complexes in diabetes patients carrying a TT risk genotype compared to heterozygotes that may influence control of enterovirus clearance.
Introduction
T
Peripheral blood mononuclear cells (PBMCs) are a complex mixture of immune cells, including monocytes, natural killer cells, effector B and T cells, and professional antigen-presenting cells such as dendritic cells (DCs). DCs shape both innate and adaptive immune responses, including antiviral immune responses. They continuously sample their microenvironment through phagocytosis and are critical for efficient priming of (naive) T cells (2,27). Depending on the signals, a DC receives either an inflammatory response or more tolerogenic immune response is induced (17). These signals include pathogen- and danger-associated molecular patterns that bind to pattern recognition receptors (PRRs) on immune cells. PRRs that are involved in antiviral responses include Toll-like receptor (TLR) 3 and TLR7/8, which sense double-stranded and single-stranded RNA, respectively (17,22). TLR4 recognizes bacterial cell wall components and has also been shown to respond to CVB in vitro as well (41). In addition, intracellular retinoic acid-inducible gene I (RIG-I)-like receptors (RLRs) such as interferon induced with helicase C domain 1 (IFIH1)/melanoma differentiation-associated protein 5 (MDA5) are involved in sensing virus infection (17). Differences in DC numbers and DC function have been reported between type 1 diabetes patients and healthy donors (1,29,43,47), yet some data are contradictory and further research is required to elucidate possible differences. Furthermore, studies on possible differences in DC antiviral responses are warranted. DCs are naturally present in blood (9,24), but they can also be differentiated in vitro from monocytes yielding monocyte-derived DC (moDC) (32).
Type I interferons (IFNs) are key mediators of antiviral responses; however, they may also favor development of autoimmune diabetes (6,50). The RLRs, RIG-I and MDA5 (melanoma differentiation-associated gene 5), both sense cytoplasmic viral RNA resulting in innate immune activation and production of IFNs. IFNs subsequently induce interferon-stimulated genes (ISGs), including the RLRs, to ensure adequate responses. In addition, effector molecules such as protein kinase R and 2′-5′-oligoadenylate synthetase 1 (OAS1), which sense double-stranded RNA and, subsequently, activate RNaseL, result in degradation of (viral) RNA and induce cellular apoptosis (12,35), which are induced to halt the infection. Intriguingly, gene expression analysis revealed a type I IFN signature before seroconversion to islet autoimmunity (11,16,31) supporting the role of viral infections in development of autoimmune diabetes. In addition, polymorphisms in the MDA5 protein that recognizes viral RNA, including enterovirus-derived RNA (10), have been associated with risk for type 1 diabetes development (36,40). rs1990760 is a common type 1 diabetes-associated single nucleotide polymorphism (SNP) in the IFIH1 gene causing an alanine to threonine substitution at amino acid position 946 in the MDA5 protein. How this SNP influences antiviral responses is currently not fully clarified. Type 1 diabetes-associated IFIH1 polymorphisms were reported to have no or only minor influence on the occurrence, quantity, or duration of enterovirus infection in the gut (46); however, in blood, higher frequencies of enterovirus RNA were detected in heterozygotes (4). Murine studies revealed functional impact of the A946T polymorphism (13), but how precisely it influences antiviral responses in human tissues such as immune cells or islets of Langerhans remains to be determined.
We hypothesize that differences in the innate antienteroviral responses exist between type 1 diabetes patients and healthy subjects resulting in altered antiviral responses and that the common A946T SNP in MDA5 modulates these responses. Therefore, we studied the expression and induction of ISGs in PBMCs of healthy donors and type 1 diabetes patients in response to enterovirus, virus-antibody complexes, or ligands mimicking viral infection and studied whether the magnitude of ISG expression in unstimulated and stimulated conditions correlates with the A946T genotype.
Methods
Sample collection and storage
Peripheral blood of healthy blood donors or type 1 diabetes patients was obtained following institutional guidelines and after written consent. Inclusion criteria were as follows: patients diagnosed with type 1 diabetes having stable disease who, besides insulin, were not taking additional medication that might influence the immune system such as glucocorticoids. Patients were recruited from the Outpatient Clinic of the Radboud University Medical Center (UMC). Healthy donors were volunteers employed at the Radboud UMC and were matched for day of sampling and blood cell isolation to exclude effects due to material handling and/or storage. Characteristics of patients and healthy individuals are shown in Table 1. PBMCs were isolated using Ficoll separation and washed in phosphate-buffered saline (PBS)/1% human serum albumin/2 mM ethylenediaminetetraaceticacid, and a portion was frozen in 10% dimethyl sulfoxide (DMSO) in liquid nitrogen until use in experiments. moDCs were generated through adherence of monocytes to culture flasks using 2% human serum for 1 h, followed by washes with PBS to remove unbound lymphocytes and subsequent differentiation using interleukin (IL)-4 and granulocyte macrophage-colony stimulating factor as described (20). At day 3–5 of differentiation, moDCs were harvested using cold PBS and frozen in 10% DMSO until use in experiments. moDCs were thawed and cultured for an additional 1–3 days in IL-4 and GM-CSF to obtain “day 6 immature moDCs,” which were used for experiments.
BMI, body mass index; NS, not significant.
Stimulation of cells
PBMCs were stimulated with recombinant IFN-α2 (Roferon, Roche; 500 U/mL), TLR-ligand polyinosinic:polycytidylic acid (poly I:C) (Enzo Life Sciences; 20 μg/mL), R848 (Enzo Life Sciences, 4 μg/mL), or lipopolysaccharide (LPS; Sigma; 100 ng/mL) or were exposed to CVB3 at a multiplicity of infection (MOI) 100, CVB/antibody complexes equaling MOI 100, or exposed to the same equivalent antibodies without virus. CVB3 was prepared as described (20). Human normal immunoglobulin (Nanogam, intravenous immunoglobulins, IVIg) was from Sanquin. Virus-antibody complexes were prepared by mixing virus and antibody solutions 1:1 followed by incubation at 37°C for 1 h. moDCs were stimulated with TLR ligands as for PBMCs.
mRNA isolation and quantitative polymerase chain reaction
Six (moDCs) and twenty-four (PBMCs) hours after stimulation were chosen as optimal time points for mRNA analysis based on kinetic studies using cells from healthy volunteers. At these time points, a robust induction of ISGs was observed (data not shown). mRNA isolation, cDNA synthesis, and quantitative polymerase chain reaction (qPCR) were performed as de,scribed (33). Expression of MDA5, RIG-I, and OAS1 was analyzed relative to the housekeeping gene Porphobilinogen deaminase as described (33). Briefly, RNA was treated with DNase I (amplification grade; Invitrogen) and reverse transcribed into cDNA using random hexamers and Moloney murine leukemia virus reverse transcriptase (Invitrogen). To exclude genomic DNA contamination, we included a “–RT” control, in which the reverse transcriptase was replaced with RNase-free water. The “–RT” control was taken along in the qPCR analysis. cDNA was stored at −20°C until further use. mRNA levels for the genes of interest were determined by qPCR with a Bio-Rad CFX apparatus (Bio-Rad) with SYBR Green (Applied Biosystems). Analysis was done using Bio-Rad CFX-1.6 software; mRNA levels of the genes of interest were normalized to mRNA levels of the housekeeping gene glyceraldehyde 3-phosphate dehydrogenase and were calculated according to the cycle threshold method. Primers are available upon request.
Polymorphism determination
Genotyping of SNP rs1990760 (A946T) was performed using Applied Biosystems SNP Genotyping Assay according to manufacturer's instructions.
Statistical analysis
Statistical analysis was performed using two-way analysis of variance, Mann–Whitney U-test, Student's t-test, or z-test for two population proportions as indicated. A p-value <0.05 was considered significantly different.
Results
Patient characteristics
We included 12 type 1 diabetes patients (seven male, five female, mean age 37.2 years, range 21–52 years) and 12 healthy donors (six male, six female, mean age 35.8, range 26–60 years). Patient characteristics are described in Table 1.
Immune cells from healthy donors and type 1 diabetes patients induce similar levels of MDA5, RIG-I, and OAS1 upon stimulation
First, we set out to determine ISG expression in resting and activated PBMC and moDC from diabetes patients and healthy controls. The innate immune sensors, MDA5, RIG-I, and OAS1, were selected as they are differentially expressed between individuals during seroconversion and healthy control subjects (16). Furthermore, they are part of the IFN signature that is preceding autoimmunity in children genetically at risk for type 1 diabetes (11). To monitor the induction of antiviral responses, PBMCs were stimulated with recombinant IFN-α, TLR ligands, or enterovirus. ISG induction was determined as a measure for the strength of the antiviral response. Steady-state expression levels of MDA5, RIG-I, and OAS1 in unstimulated PBMCs were largely similar in type 1 diabetes patients compared to healthy donors (maximum two-fold average difference in diabetes patients compared to healthy donors) (data not shown). DCs are known to be important in clearance of viral infections and express multiple ISGs. Analysis of unstimulated moDCs revealed that no differences between healthy donors and diabetes patients exist (data not shown).
PBMCs from patients and healthy donors significantly induced ISG expression upon stimulation with IFN-α and poly I:C, a synthetic TLR3 and MDA5 ligand, mimicking double-stranded viral RNA (Fig. 1A; Supplementary Fig. S1A; Supplementary Data are available online at
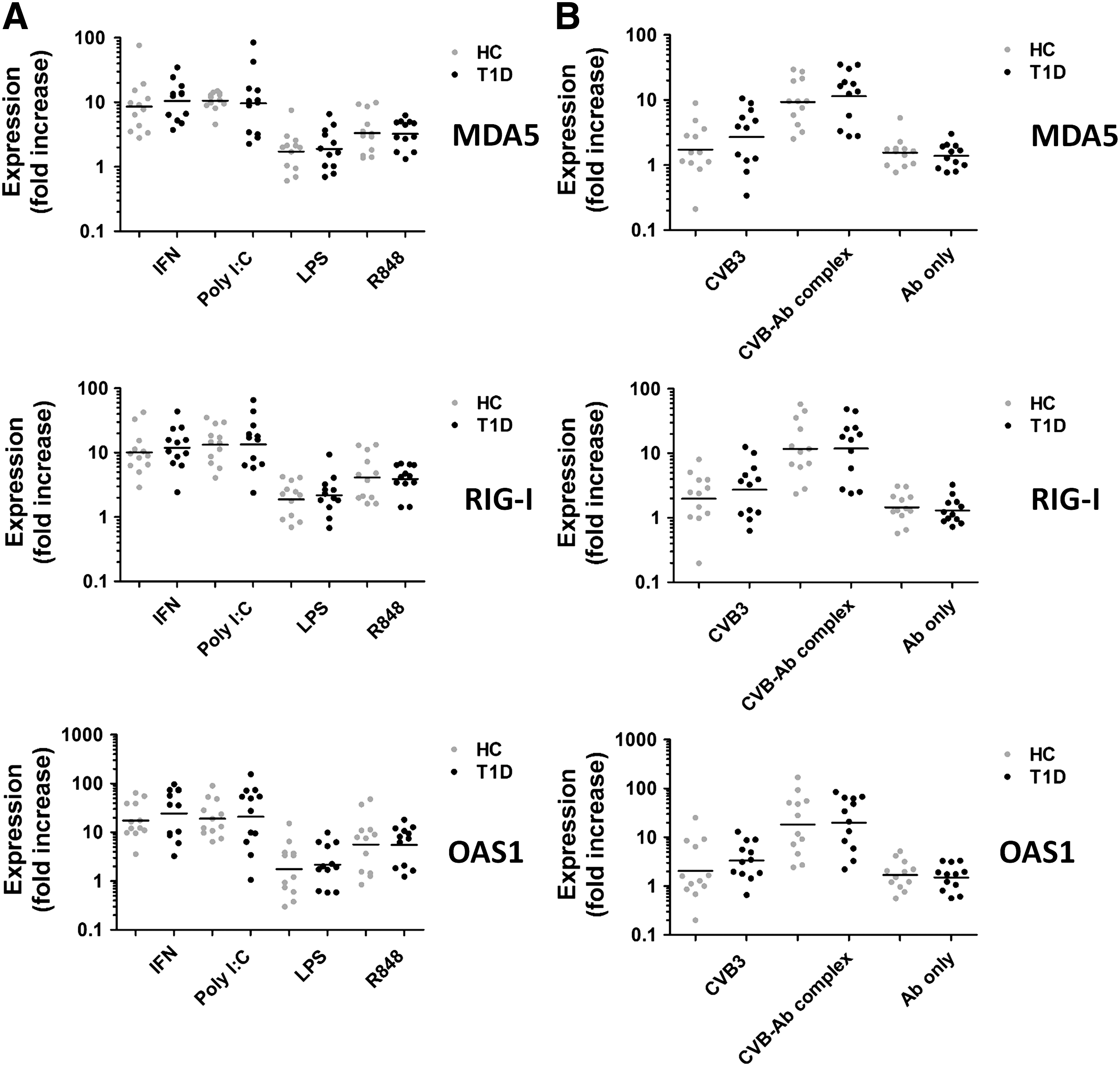
No differences in induction of ISGs in PBMCs of healthy donors and type 1 diabetes patients upon stimulation.
Stimulation with synthetic TLR ligands is rather different than encountering infectious virus. Therefore, we also determined the induction of ISGs in PBMCs in a more physiological encounter with CVB or CVB-antibody complexes (14,19,44). Of these pathological stimuli, CVB-antibody complexes induced the strongest responses (Fig. 1B; Supplementary Fig. S1B), reaching gene expression levels similar to exposure to the strong synthetic stimulus poly I:C. No significant differences between healthy donors and diabetes patients were observed. As a control, PBMCs were exposed to the antibodies without CVB. Exposure of PBMCs to the antibodies without CVB had no effect compared to medium alone, while exposure to CVB alone resulted in very heterogeneous expression of all ISGs tested in both the healthy control group, as well as the type 1 diabetes group (Fig. 1B; Supplementary Fig. S1B). No induction of ISGs was observed upon stimulation of moDCs from healthy donors with virus-antibody complexes (data not shown).
Elevated ISG induction in diabetes patients heterozygous for the MDA5 SNP rs1990760 (A946T) upon encounter of CVB and CVB-antibody complexes
For our SNP analysis, we included data from two additional healthy donors that were used for initial kinetic studies (see “Methods” section). Genotyping of the individuals included in our cohort revealed that the diabetes group contained more individuals with the TT risk genotype (healthy donors 6/14, type 1 diabetes patients 7/12), whereas in the healthy control group, slightly more heterozygotes were present (healthy donors 7/14, type 1 diabetes patients 4/12) (Table 2). Despite a relatively small sample size, this trend matches observations by Smyth et al. that the TT risk genotype is overrepresented in autoimmune diabetes patients compared to healthy individuals (36). Due to the low number of CC genotypes (one in each group), these samples were excluded from the analysis and discussion. In the total group, the basal expression levels of MDA5, RIG-I, and OAS1 in PBMCs were similar in the TT group compared to heterozygotes (data not shown). Similarly, in moDCs, no differences in basal expression were observed between the genotypes (data not shown).
Responses to stimulation were first analyzed for combined type 1 diabetes patients and healthy donors and subsequently for both groups separately. In the total group, no significant differences were present in poly I:C- or IFN-α-stimulated PBMCs in the CT heterozygotes compared to the TT (risk) group (Fig. 2A, B). In addition, when analyzing type 1 diabetes patients and healthy controls in separate groups, no significant differences were observed (Supplementary Fig. S3A, B). In PBMCs stimulated with CVB, heterozygotes showed a trend to higher induction (Fig. 2C). Strikingly, upon stimulation with the physiologically relevant enterovirus-antibody complexes, the induction of MDA5 was significantly higher (p = 0.0239) in the CT group compared to TT homozygotes (Fig. 2D). No differences were present in control stimulation with antibodies alone (Supplementary Fig. S4). Analyzing type 1 diabetes patients and healthy donors separately revealed that the differences between TT and CT genotypes in healthy donors were rather small and not significantly different (Fig. 3A, B, lower panels). In contrast, in the diabetes patients, the observed differences upon enterovirus stimulation were significant for all three ISGs tested (Fig. 3A). For MDA5 also, ISG induction upon stimulation with virus-antibody complexes was significantly higher in heterozygotes compared to TT homozygotes in the T1D-group (p = 0.0242) (Fig. 3B). These data demonstrate that diabetes patients heterozygous for A946T have an enhanced response to enterovirus and enterovirus-antibody complexes compared to TT homozygote diabetes patients.
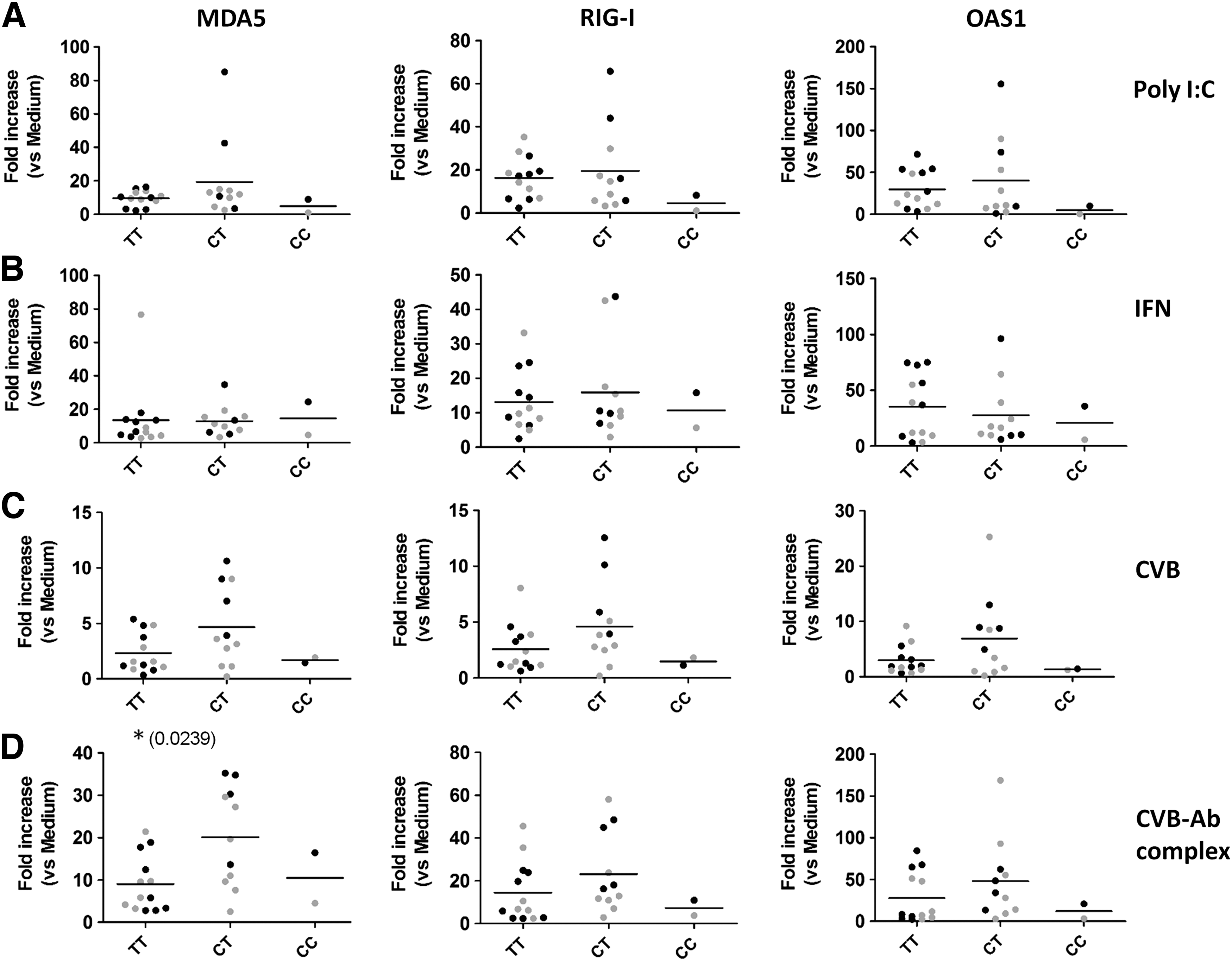
Enhanced ISG induction in heterozygotes for the A946T (rs1990760) polymorphism upon stimulation with CVB-antibody complexes compared to the TT risk genotype. ISG expression for PBMCs stimulated as in Figure 1 is shown per genotype for the rs1990760 SNP. Data for healthy donors (gray) and type 1 diabetes patients (black) combined are shown for the following stimulation:
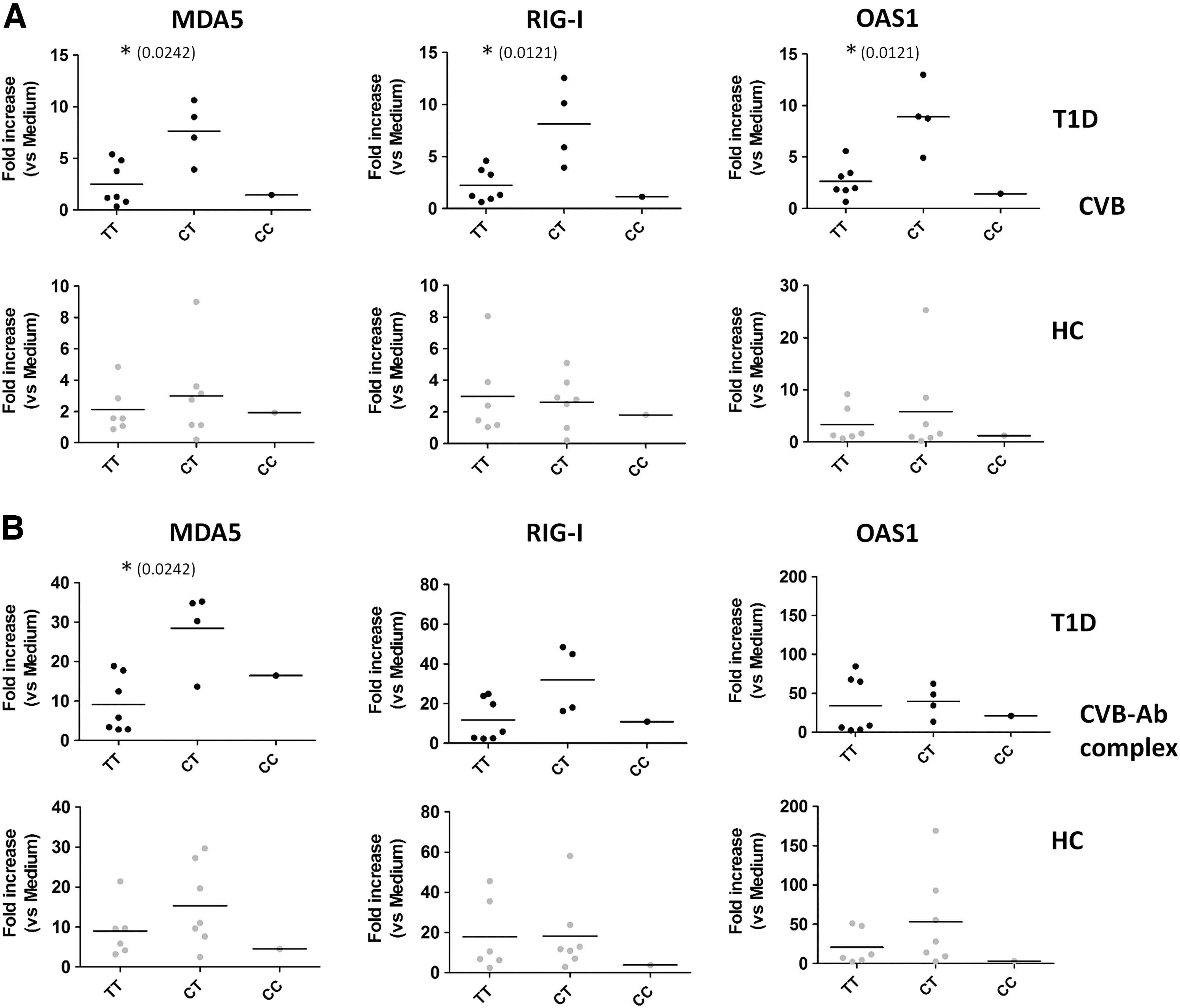
Enhanced ISG induction in type 1 diabetes patients, but not healthy donors, heterozygous for the A946T (rs1990760) polymorphism upon stimulation with CVB and CVB-antibody complexes. ISG expression for PBMCs stimulated with CVB
Discussion
Viruses in general and enteroviruses in particular have been associated with the development of type 1 diabetes (15,48). Yet, only limited data are available regarding the response of type 1 diabetes patients to (entero) viruses in comparison to healthy individuals, and the impact of the type 1 diabetes-associated A946T polymorphism in the innate immune sensor MDA5 herein remains elusive. Our data now show that specifically in diabetes patients heterozygous for the MDA5 A946T polymorphism, encounter with CVB and CVB-antibody complexes resulted in elevated expression of ISGs compared to the TT risk genotype. This difference was not observed in healthy controls. These data imply an intrinsic difference in the response pathway to enteroviruses in diabetes patients carrying a TT risk genotype compared to TC heterozygotes. The number of patients carrying the CC phenotype was too low to draw any conclusion.
Previous studies have shown that an IFN signature in unstimulated blood or PBMCs is correlated with seroconversion/type 1 diabetes development (11,16,31). However, whether differences exist in, for example, antiviral responses between individuals at risk for diabetes or manifest diabetes patients compared to healthy donors cannot be derived from these data. We studied functional antiviral responses in individuals with clinical type 1 diabetes. Our data in a limited number of patients show that the differences observed for the MDA5 A946T polymorphism are inherent to diabetes patients. The small sample size is a limitation of the current study. Therefore, it is important to reproduce the data in a larger independent cohort, and in addition, future studies should assess PBMCs from individuals at risk, recent onset type 1 diabetes patients, and patients with established diabetes to further substantiate these differences and the possible impact in the different patient groups. In the current study, we selected CVB3 and an MOI of 100 for stimulation based on previous data published by Wang et al. (44), who reported dose-dependent induction of type I IFN upon exposure to CVB-antibody complexes. In future studies, an important addition could be comparison between the response to different CVB serotypes, as well as a more in-depth analysis of the dose–response to CVB and CVB-antibody complexes for healthy individuals and type 1 diabetes patients.
For moDC, we did not observe any differences between healthy donors and type 1 diabetes patients in basal expression nor upon stimulation. In addition, no response to enterovirus-antibody complexes was observed in moDC. An explanation for this observation might be that activating Fc receptor (FcR) expression is downregulated by IL-4 during the 1 week culture period (30,38) needed to generate moDC.
To understand the impact of the common A946T SNP in MDA5 and its interactions with other signaling cascades and pathways, we studied the responses to virus or conditions mimicking virus infection in correlation with the rs1990760 SNP. Strikingly, the most significant differences were observed between the TT risk genotype and heterozygotes when using enterovirus and virus-antibody complexes, underscoring that differences may not be readily apparent in unstimulated conditions. Moreover, our data now show that the detection of differences in innate responses may not simply depend on stimulation per se, but is also greatly affected by the type of stimulation. Notably, depending on particular stimulations (e.g., recombinant IFN-α, poly I:C, or virus-antibody complexes), individuals were found to be high responders or average responders implying that different expression levels of the receptors involved (e.g., IFN-α/β receptor, TLR-3, and FcR) have a major influence on the outcome. Our data show that exposure to enterovirus-antibody complexes yields a strong innate immune response and that its magnitude is critically influenced by the A946T SNP in autoimmune diabetes patients, but not in healthy controls. FcR mediate the recognition of enterovirus-antibody complexes at the cell surface (44), and synergy between FcR and PRRs such as TLRs has recently been shown to influence the magnitude of responses (5,42). Our data imply that probing the type I IFN response pathway with a physiologically relevant stimulus, like enterovirus or enterovirus-antibody complexes, is most informative and supports a role for MDA5 and possibly FcR in this process. Furthermore, our data revealed this effect using PBMCs, but not in vitro generated moDC. PBMCs contain multiple different immune cells, including different DC subsets, allowing extensive immune cell cross talk. Although challenging due to limited cell numbers, it will be interesting to define the role of the different immune cells in the observed effects. Plasmacytoid DCs (pDCs) can present exogenous peptides derived from protein-antibody complexes to CD4+ T cells using Fcγ-receptor mediated uptake and are known to express high levels of MDA5 (3). In addition, they enhance presentation of soluble islet tyrosine phosphatase IA-2 in the presence of IA-2+ autoantibody patient serum (1) and augment Th1 responses in type 1 diabetes through IFN-α production (47). For the response to CVB-antibody complexes, pDCs may thus play a significant role (19,44), although other FcR bearing myeloid cell types such as monocytes (14) or myeloid DCs may be involved as well.
How the A946T SNP precisely exerts its influence on the various pathways stimulated is currently unknown. It is known that initial virus–host interactions involving RLRs and/or TLRs can dictate the course of virus infection (37). Previous studies have suggested that rare loss-of-function mutations in MDA5 provide protection against type 1 diabetes (26,34) implying that MDA5 is involved in the risk of autoimmunity. Recent mouse studies support the hypothesis that enhanced MDA5 activity promotes autoimmune disease, since mice carrying a mutant form of MDA5, which is constitutively active, develop a systemic lupus erythematosus-like autoimmune disease (13), and mice deficient for one MDA5 allele (MDA5+/−) are protected from diabetes development upon CVB encounter (21). Interestingly, however, children heterozygous for the A946T SNP were found to have higher frequency of enterovirus RNA in their blood relative to Ala/Ala and Thr/Thr genotypes (4). Previous studies analyzing MDA5 expression levels using human PBMCs showed contradicting results with respect to the consequence of A946T SNP on MDA5 expression. Marinou et al. and Panarina et al. found no difference in MDA5 expression in PBMCs between genotypes (25,28), whereas others found that haplotypes containing the Thr946 (risk) allele produced more IFIH1 mRNA than the Ala946 haplotype (8,23). Our functional studies looking at the consequences of virus exposure rather than analyzing steady-state expression levels suggest that the induction of MDA5, as well as two other ISGs, is higher in TC heterozygotes compared to TT (risk) homozygotes. These findings imply that Thr946 homozygotes have a lower response to CVB and CVB-immune complexes that may result in suboptimal or delayed virus clearance. How this could favor induction of autoimmunity is unknown, but one possibility is that the delayed virus clearance leads to more exposure causing prolonged activation of the type I IFN pathway. Such prolonged exposure to type I IFNs as a consequence of smoldering infections may promote autoimmune responses in genetically susceptible individuals. Moreover, delayed virus clearance may increase the chance of enterovirus infecting β-cells in islets of Langerhans. Strikingly, the difference in ISG induction upon CVB and CVB-Ab complexes between TT and CT was significantly different in the type 1 diabetes group, but not in healthy individuals. This raises the intriguing possibility that this response may be a predictive readout in prediabetic subjects.
In summary, our data show that the common MDA5 SNP rs1990760 influences antiviral responses in type 1 diabetes patients, particularly upon encounter of physiologically relevant stimulation with CVB and CVB-antibody complexes. These findings imply that within the genetic background of type 1 diabetes, this MDA SNP significantly impacts responses to enterovirus, consistent with type 1 diabetes being a multifactorial disease.
Footnotes
Acknowledgments
The authors thank all professionals involved in collection of human material and all individuals who agreed to participate in this study. This work was supported by grant 24-2008-949 from the Juvenile Diabetes Research Foundation (to G.J.A.), Ruby grant 2011.80.1427 from the Dutch Diabetes Research Foundation (to B.M.S and G.J.A.), and the European Union Seventh Framework Programme [FP7/2007–2013] under grant agreement no. 261441/PEVNET (to M.F-T.). Funding agencies had no role in conducting research and/or preparation of the article.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.