
Abstract
Noroviruses (NoVs) account for the majority of diagnosed cases of viral acute gastroenteritis worldwide. Virus-like particle (VLP)-based vaccines against NoV are currently under development. Serum antibodies that block the binding of NoV VLPs to histo-blood group antigens, the putative receptors for NoV, correlate with protection against NoV infection. The role of functional mucosal antibodies in protection is largely unknown, even though the intestinal mucosa is the entry port for NoV. Balb/c mice were immunized intramuscularly (IM) or intranasally (IN) with NoV GII.4 VLPs, and systemic and mucosal blocking antibody responses were studied. IN immunization elicited NoV-specific serum and mucosal IgG and IgA antibodies, whereas IM immunized animals completely lacked IgA. Both immunization routes induced similar blocking activity in serum but only IN route generated blocking antibodies in mucosa. The level of IgA in the mucosal (nasal) lavages strongly correlated (r = 0.841) with the blocking activity, suggesting that IgA, but not IgG, is the major NoV blocking antibody on mucosal surfaces. The results indicate that only mucosal immunization route induces the development of functional anti-NoV IgA on mucosal surface.
Introduction
N
Histo-blood group antigens (HBGAs) are complex carbohydrates expressed on red blood cells and, in individuals with a functional fucosyltransferase 2 (FUT 2) gene, also on mucosal surfaces and body secretions (23). HBGAs likely serve as attachment factors or receptors for NoVs in a genotype-specific manner (9,24,31). An established correlate of protection to NoV infection is the ability of serum antibodies to block the binding of NoV VLPs to HBGAs (3,26,30). High levels of NoV-specific serum IgG antibodies alone were not protective in adults challenged with NoV (3,12,20), but prechallenge NoV-specific memory IgG B cells have been recently identified as a new correlate of protection against NoV gastroenteritis (GE) (29). Concurrently, salivary IgA (18,29) and serum IgA antibody levels (3) have been associated with protection from NoV GE. Only a study by Lindesmith et al. (19) addressed the functionality of NoV-specific serum IgA antibodies, namely the blocking activity. They showed that human IgA, purified from NoV GI.1-infected volunteers' sera, blocked the binding of strain-specific and heterotypic VLPs to HBGAs (19).
However, studies determining the role of mucosal immune response in protection from NoV infection are still limited. This is of a particular importance as mucosa is an entry port for NoV. We have previously shown that mucosal and systemic immunizations of mice with NoV VLPs induce qualitatively different B- and T-cell responses in mice (22). Here, we studied whether the immunization route and the isotype of induced NoV-specific antibodies play a role in the blocking activity on mucosal surface.
Materials and Methods
Mice immunizations and collection of samples
Female Balb/c mice were immunized at study weeks 0 and 3 with 10 μg NoV GII.4 (1999) VLPs via intramuscular (IM) or intranasal (IN) route and terminated at study week 5 as described in detail earlier (22). Baculovirus–insect cell expression system production, purification, and characterization of the GII.4 VLPs are described elsewhere (5,10). Mice receiving carrier only (sterile phosphate-buffered saline [PBS]; Lonza) were used as negative controls. Mice were terminated by cutting the axillary artery under termination anesthesia obtained with ketamine (Ketalar®; Pfizer Ltd.) and medetomidine (Dorbene®; Syva). Whole blood (serum) was collected and processed as previously described (5). Nasal washes (NWs) were collected by injecting sterile PBS (2X 125 μL) via the trachea into the nasopharynx, and the lavages were collected from the nostrils and centrifuged (12,000 × g for 10 min at +4°C), and the clear supernatants were stored at −20°C. Before terminations, stool samples were collected as described previously (5). All procedures were conducted according to the regulations of the Finnish Animal Experimental Board.
Immunoassays
Sera and NWs of immunized mice were tested in enzyme-linked immunosorbent assay (ELISA) for the presence of NoV GII.4-specific IgG and IgA antibodies as described in detail elsewhere (22,33). Briefly, GII.4 VLPs were coated (1 μg/mL) on 96-well polystyrene plates (Corning, Inc.). Sera diluted serially from 1:200 (IgG ELISA) or 1:20 (IgA ELISA) or NWs (diluted 1:5) were added on the plates, and the bound antibodies were detected with horseradish peroxidase (HRP)-conjugated IgG or IgA anti-mouse antibody and OPD substrate (all from Sigma-Aldrich). Optical density (OD) values at 490 nm were measured by a microplate reader (Victor2 1420; PerkinElmer). A sample was considered positive if the net OD was above the cutoff value: mean OD +3 × standard deviation of the control mice and OD ≥0.100. Endpoint titer was determined as the final dilution, giving an OD above the set cutoff value. If the first dilution tested was below the set cutoff limit, the sample was assigned a value half of the detection limit of the assay.
For the detection of NoV blocking antibodies in sera and NWs, a surrogate neutralization assay for NoV utilizing saliva as a source of HBGAs was used (5,22). Briefly, type A saliva (1:3,000 dilution) was coated on 96-well microplate (Corning, Inc.). Serial dilutions of sera (1:50–1:800), NW samples (1:5), or 10% stool suspensions (diluted serially from 1:5) were preincubated with GII.4 VLPs (0.1 μg/mL) for 1 h at +37°C and added to the plate. The saliva-bound VLPs were detected from the plate using human anti-NoV immune serum and HRP-conjugated anti-human IgG antibody (Novex; Thermo Fisher Scientific) reacting with OPD substrate. The maximum binding of GII.4 VLPs (OD ≥1.600) was obtained from the wells without serum preincubation. The blocking index (%) was calculated as follows: 100% − (OD wells with VLP–serum mix/OD wells without serum; maximum binding) × 100%.
Statistical analyses
Fisher's exact test was used to assess the intergroup differences in the IgG and IgA endpoint titers. Mann–Whitney U-test was used to compare the differences in the NW antibody levels. Spearman rank correlation coefficient was used to examine the correlation between the blocking index (%) and the antibody magnitude. Statistical significance was defined as p < 0.05. All hypothesis testings were two tailed.
Results and Discussion
The role of mucosal antibodies in protection is largely unknown but cannot be neglected. To address this, we immunized mice IM or IN with NoV GII.4 VLPs and assessed the ability of sera and NWs (used as a proxy for mucosal samples) to block the binding of GII.4 VLPs to HBGAs. To our knowledge, this is the first study describing the correlation between mucosal NoV-specific IgA antibodies and HBGA blocking activity.
Both immunization routes elicited similarly high NoV-specific serum IgG antibodies (p > 0.05) with blocking activity (data not shown), as earlier reported by our group (22). Only IN immunization elicited detectable NoV-specific IgA antibodies in serum (GMT 120, 95% CI = 80–180), which did not have an increasing effect on the total serum blocking ability (data not shown). However, when we tested the blocking activity of the mucosal samples (NWs) from IM and IN immunized mice, a remarkable difference was observed (Fig. 1A, B). IN immunized mice had superior blocking activity, even though the mean levels of NoV GII.4-specific IgG were similar between IM and IN immunized mice (p > 0.05) (Table 1). Importantly, only IN immunized mice had anti-GII.4 IgA antibodies detected in the NW samples (Table 1), suggesting that IgA could be the major blocking antibody isotype in the mucosa. Indeed, correlation analyses showed a strong positive correlation between NW IgA levels and the blocking activity (r = 0.841, p < 0.001, Fig. 1C). In contrast, IgG level did not correlate with the blocking activity (r = −0.223, p > 0.05, Fig. 1D). Studies to determine a direct blocking activity of purified IgA from the mucosal lavages are warranted. The matter is complicated by the fact that no reliable methodology for mouse IgA purification exists.
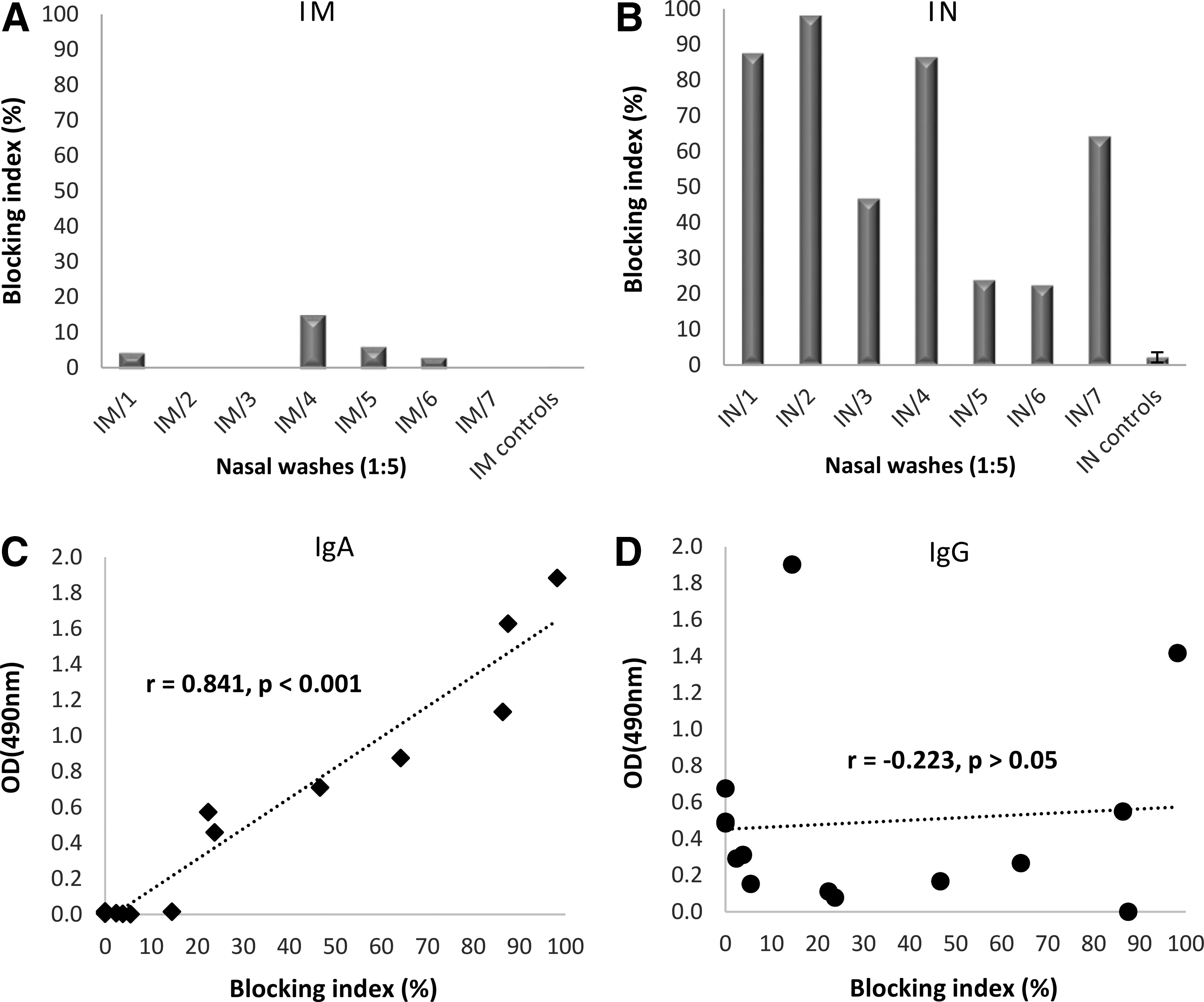
Norovirus GII.4-specific mucosal antibody responses. Balb/c mice were immunized IM or IN two times with 10 μg GII.4 VLPs, and NWs were collected at the time of termination as described in the “Materials and Methods” section. Mice receiving carrier buffer (PBS) only were used as negative controls (ctrls). A surrogate neutralization assay was used to detect NW (diluted 1:5) ability to block GII.4 VLP binding to HBGAs in human type A saliva. Shown are blocking indexes of individual mice immunized IM
Nasal washes were tested at 1:5 dilutions in ELISA.
Group mean OD values with standard errors of the mean (±SEM).
ctrls, Control mice; ELISA, enzyme-linked immunosorbent assay; Ig, immunoglobulin; IM, intramuscular; IN, intranasal; ND, not determined; NoV, norovirus; OD, optical density.
Our results confirm a significant NoV blocking activity of serum anti-NoV IgG (5,8,21,22) and suggest that mucosal antibodies may have the capability to block NoV infection as well. The blocking activity correlated with NoV-specific mucosal IgA but not IgG. Purified human serum anti-NoV GI.1 IgA was recently shown to have a good HBGA blocking activity, indicating that serum IgA antibodies are functional (19). Mucosal IgA is actively secreted in dimeric or polymeric form, namely secretory IgA (sIgA), to the luminal surface of epithelium, where it is known to be the major player against microbial infections (7). In serum, IgA is mostly present in the monomeric form lacking the secretory component (14). Presumably, anti-NoV sIgA with high valences of antigen-binding sites has more blocking potential on the mucosal surface in contrast to the low level of monomeric IgG exuded from serum. The blocking effect of serum exuded monomeric IgA in mucosal secretions is probably negligible as it is shown that <0.5% of antigen-specific monomeric IgA is transported from serum to nasal secretions of immunized mice (25). For human immunodeficiency virus (HIV), it has been shown that dimeric form of anti-HIV IgA inhibits virus mucosal transmission more effectively than its monomeric counterpart or monomeric IgG (11). Mucosally delivered vaccines are known to induce sIgA responses at the site of antigen delivery as well as at the distant mucosal sites (1,17,37). As the intestine is the NoV infection route, we first performed blocking assays with mouse stool suspensions, which also contain NoV-specific IgG and IgA (22). However, the stool suspensions, even from control naive mice, caused high unspecific blocking (data not shown) for unknown reasons; therefore, NWs were used as a proxy for mucosal samples.
Contradictory findings exist for the significance of NoV-specific mucosal immunity. Fecal IgA has not been shown to correlate with the protection (27,29), but it has been associated with reduced viral load and shorter duration of virus shedding (29). In contrast, preexisting salivary IgA (3) and the early (<5 days) elevation of the salivary IgA postinfection (18) have been associated with protection. Recent adult human trials with NoV VLPs delivered IM have indicated that NoV protective immunity seems to be based on rapid recall response by IgA biased NoV-specific memory B cells generated by previous exposure to NoV in natural infections (29,32). It may be that routes of immunization, which induce mucosal IgA, should be favored in NoV vaccine development. Future trials in pediatric population are needed to determine the optimal immunization route for this naive population. It is possible that to naive children, the vaccine should be delivered mucosally, while for adults and preexposed individuals, IM administration may be sufficient. Our results further underline the importance of mucosal immunity, specifically IgA antibodies, in NoV protection.
Footnotes
Acknowledgments
The laboratory personnel Marianne Karlsberg, Eeva Jokela, Sanne Kaven, Nina Koivisto, and Marjo Salminen and researchers Suvi Lappalainen, Leena Huhti, and Hanni Uusi-Kerttula are warmly thanked for their valuable technical assistance and help during the study.
Author Disclosure Statement
No competing financial interests exist.