
Abstract
Regulatory T cells (Tregs) and interleukin-17-producing T helper (Th17) cells were mutually antagonistic in the pathogenesis of chronic hepatitis C virus (HCV) infection. However, the regulation of imbalance between Tregs and Th17 cells was poorly understood in HCV infection. A recent report revealed the immunomodulatory role of Toll-like receptor (TLR) 2 in regulating the balance of Tregs/Th17 functions in multiple sclerosis. Thus, the aim of the current study was to assess the effect of TLR2 stimulation on the suppressive function of Tregs and Th17 differentiation in chronic hepatitis C. A total of 65 patients with chronic hepatitis C receiving pegylated interferon-a2a and ribavirin therapy for 48 weeks, as well as 20 of normal controls (NCs) were enrolled. Cellular proliferation and cytokine production was tested in purified CD4+CD25+CD127dim/− Tregs in response to the stimulation of Pam3Csk4, an agonist of TLR2. In treatment-naive patients, Tregs, but not Th17 cells, from chronic hepatitis C patients expressed higher levels of TLR2 compared with NCs. Stimulation with Pam3Csk4 enhanced the suppressive function of Tregs and production of IL-10 in chronic hepatitis C more than in NCs. However, TLR2 stimulation did not promote Th17 differentiation of Tregs in chronic hepatitis C patients. Moreover, effective anti-HCV therapy resulted in the induction of IL-17-secreting phenotypic shift of Tregs without loss of inhibitive function upon TLR2 stimulation. These data provided a novel mechanism underlying modulating the balance of Tregs/Th17 cells in chronic hepatitis C. HCV infection shifted Tregs/Th17 cells through TLR2 stimulation by inducing Tregs to produce IL-10 and enhancing inhibitive function of effector T cells, resulting in viral persistence.
Introduction
H
HCV has been demonstrated to be a noncytopathic pathogen, which induces liver injury by stimulation of the immune system. Therefore, the interaction between virus and host immune response may influence the outcome of chronic hepatitis C (11). Furthermore, effective clearance of HCV requires robust and specific antiviral T cell response (18,30). However, patients often exhibit impairment of HCV-specific T cell activity and generated insufficient T-cell response against the virus during chronic HCV infection (8,12). The precise mechanism associated with T cell tolerance is still not fully elucidated, and identification of immunological factors associated with clinical outcomes in response to antiviral therapy is also important to improve clinical management.
Naive CD4+ T cells can differentiate upon activation into different lineages of T helper (Th) cells, which differ in the cytokine productions and their functions. CD4+CD25+ regulatory T cells (Tregs) and interleukin (IL)-17-producing CD4+ T (Th17) cells are two additional Th cell subsets with reciprocal relationship (3,5). Recent studies revealed that chronic hepatitis C patients have elevated frequencies of Tregs in peripheral blood (2,32), liver (28,29), and even in their colon (15). The increased proportions of Tregs, which were associated with HCV viral loads, suppressed the proliferation of viral-specific CD8+ T cells in persistent HCV infections (7,25). Effective anti-HCV therapy led to the decrease of Tregs (1), and in vitro study also suggested that anti-HCV agents reversed HCV-specific Tregs-mediated suppression of T effector cells (16). Th17 cells were also significantly increased in chronic hepatitis C patients, which were considered to be vital factors of triggering liver inflammation (9,27). Importantly, Tregs and Th17 cells, which may be generated from the same precursor T cells, were closely associated with each other (4). Transforming growth factor-β and IL-6 were essential for the differentiation of both Tregs and Th17 cells in a concentration-dependent manner (36).
More recently, Hao et al. revealed that the imbalance of Tregs/Th17 cells contributed to the persistent HCV infection (13). However, there were few reports on the regulatory factors which modulated the balance of Tregs/Th17 cells in chronic hepatitis C. Toll-like receptor (TLR) 2, which is a pattern recognition receptor and plays a central role in the initiation of innate immune response against invading pathogens, was demonstrated to regulate the balance between Tregs and Th17 functions in multiple sclerosis (22). Hence, we hypothesized that TLR2 also plays a regulatory role in the balance between Tregs and Th17 cells in chronic HCV infection. To test this possibility, we investigated the TLR2 expression on Tregs and Th17 cells, and assessed the effect of TLR2 stimulation on the suppressive function of Tregs and Th17 differentiation in healthy individuals and patients with chronic hepatitis C.
Patients, Materials, and Methods
Subjects
A total of 65 patients with chronic HCV infection were enrolled in this study. All patients were hospitalized or followed-up in the Affiliated Hospital to Changchun University of Chinese Medicine from July 2013 to February 2015. Twenty healthy individuals who matched for mean age and sex ratio were also enrolled for normal controls (NCs). The subjects who were coinfected with HIV or other hepatitis viruses were excluded. No subjects received antiviral or immunomodulatory treatments within 1 year of baseline sampling. All patients were diagnosed exclusion of liver cirrhosis by ultrasound and Fibroscan test. Some patients also took abdominal CT scan. The baseline characteristics of the enrolled subjects are presented in Table 1. All patients with chronic hepatitis C received pegylated interferon α2a (peg-IFN-α2a, 40KD, Roche, Shanghai, China) plus ribavirin combination therapy for 48 weeks. Blood sampling were made on baseline and 48 weeks of the therapy. Those 65 patients were divided into responders (53 patients with undetectable HCV RNA after 48-week therapy) and nonresponders (12 patients with detectable HCV replication at the end of standard therapy) (10). All patients were in good adherence to medication during therapy. This study was approved by the ethics committee of the Affiliated Hospital to Changchun University of Chinese Medicine, and written informed consent was obtained from each subject.
Data are shown as median and range.
HCV, hepatitis C virus.
Virological and biochemical assessments
Anti-HCV antibody was measured using the commercial Enzyme Immunoassay Kits (Jinhao Biotech, Beijing, China). HCV RNA was quantified using the commercial Real-Time PCR Kits (PG Biotech, Shenzhen, China) with detection limit of 2 log10 copies/mL. HCV genotyping was performed by a second-generation line probe assay (INNO-LiPA II; Innogenetics, Zwijnaarde, Belgium). Biochemical assessments were tested by Hitachi 7600 automatic analyzer (Hitachi Ltd., Tokyo, Japan).
Peripheral blood mononuclear cell isolation and CD4+CD25+CD127dim/ − Tregs purification
Peripheral blood mononuclear cells (PBMCs) were isolated by Ficoll-Hypaque (Sigma-Aldrich, St. Louis, MO) density gradient centrifugation. The isolated PBMCs were then counted and checked for viability. CD4+CD25+CD127dim/− Tregs were purified using the Human CD4+CD25+CD127dim/− Regulatory T Cell Isolation Kit II (Miltenyi Biotec GmbH, Bergisch Gladbach, Germany) according to the instruction of the manufacturer. The purity of enriched cells for CD4+CD25+CD127dim/− T cells (>90%) was determined by flow cytometry. Cells were incubated in RPMI 1640 supplemented with 10% of heat-inactivated FBS at 37°C in a 5% CO2 environment.
Flow cytometry
PBMCs were stimulated with PMA (50 ng/mL) and ionomycin (1 μg/mL), supplemented with Brefeldin A (10 μg/mL) for 6 h for measurement of Th17 cells. PBMCs were then transferred into FACS tubes, and anti-CD3-PerCP (BD Bioscience, San Jose, CA), anti-CD4-APC (eBioscience, San Diego, CA), anti-CD25-PE Cy7 (eBioscience), and anti-TLR2-PE (eBioscience) were added and incubated at 4°C in the dark for 30 min. Cells were then fixed by 100 μL of Fixation & Permeabilization Medium A (Caltag Laboratories, Invitrogen, Carlsbad, CA) for 15 min incubation, and then were resuspended in 100 μL of Fixation & Permeabilization Medium B (Caltag Laboratories) containing anti-IL-17A-FITC (eBioscience) for 20 min incubation at room temperature. In some experiments, cells were only stained with anti-CCR4-PerCP (BD Biosciences) and anti-CCR6-PE (BD Biosciences). Samples were analyzed with a BD FACSAria II analyzer (BD Biosciences). Acquisitions were performed with CellQuest Pro software (BD Biosciences) and analyses were performed with FlowJo version 8.7.2 for Windows (Tree Star, Inc., Ashland, OR).
Cellular proliferation assay
To evaluate the suppression activity, isolated CD4+CD25+CD127dim/− Tregs was cultured alone (2.5 × 104) or together with autologous CD4+CD25− responder T cells (Tresps) at ratio of 1:1, 1:4, or1:8, respectively, and stimulated with 1 μg/mL of anti-CD3 (eBiosciences) and anti-CD28 (eBiosciences). Before mixture, Pam3Csk4 (final concentration 3 μg/mL; InvivoGen, San Diego, CA) was added to the culture medium of Tregs for 24 h stimulation. For proliferation assay, cells were cultured for 5 days, with replacement of fresh medium containing 20 U/mL recombinant human IL-2 (Sigma-Aldrich) at day 3. Cellular proliferation was measured by the Cell Counting Kit-8 (CCK-8; Alexis Biochemicals, San Diego) according to the instruction of the manufacturer. Briefly, for the last 4-h culture, 20 μL of CCK-8 solution was added to each well. Wells that contained a known number of viable PBMCs was also prepared for creating a calibration curve. The absorbance of the samples was measured at 450 nm using Model 680 Microplate Reader (Bio-Rad, Hercules, CA) (34).
Enzyme-linked immunosorbent assay
Concentrations of IL-10, IL-17, IL-22, and IL-23 were measured using the commercial Enzyme-Linked Immunosorbent Assay (ELISA) Kits (eBioscience) according to the instruction of the manufacturer.
Statistical analysis
Data were analyzed using GraphPad Prism version 5.0 (GraphPad Software, La Jolla, CA). The Kruskal–Wallis H test was used for comparison between groups. All tests were two-tailed and p < 0.05 was considered to indicate a significant difference.
Results
Elevation of TLR2 expression on Tregs in chronic hepatitis C patients
Sixty-five chronic hepatitis C patients and 20 healthy individuals were enrolled in the current study. We measured the mean fluorescence intensity (MFI) values corresponding to TLR2 on either CD3+CD4+CD25+CD127dim/− T cells (Tregs) or CD3+CD4+IL-17A+ T cells (Th17 cells) by flow cytometry. All subjects clearly display the two CD4+ T cell subsets, and representative samples are shown in Figure 1A (Tregs) and Figure 1B (Th17 cells). The level of TLR2 was examined in this cell population, and typical flow histogram of TLR2 expression within Tregs (Fig. 1A) or Th17 cells (Fig. 1B) is shown. There was a remarkably higher TLR2 expression on Tregs of chronic hepatitis C patients (MFI: 26.23 ± 7.49) when compared with NCs (21.33 ± 6.07, p = 0.0081, Fig. 1C). However, we did not find a notable difference of MFI of TLR2 on Th17 cells when comparing NCs (36.46 ± 12.98) and chronic hepatitis C (40.63 ± 16.03, p = 0.465, Fig. 1C). Among the 65 chronic hepatitis C individuals, 39 patients were HCV genotype 1b and other 26 were genotype 2a. We further analyzed the TLR2 expression on Tregs and Th17 cells between the two different genotypes. There was no significant difference for TLR2 expression on either Tregs or Th17 cells between the two groups (Fig. 1D).
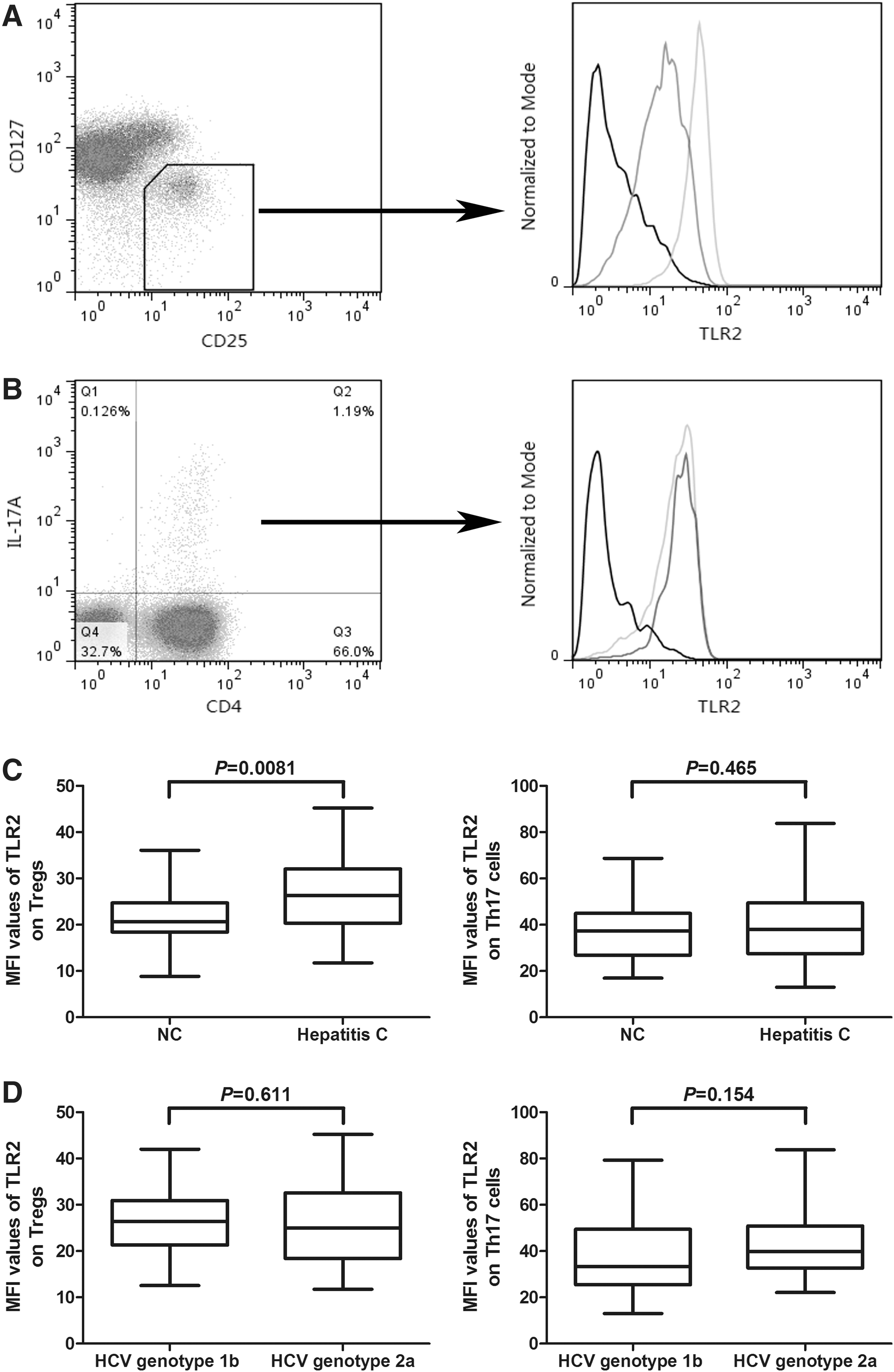
Toll-like receptor (TLR) 2 was highly expressed on CD4+CD25+CD127dim/− regulatory T cells (Tregs) in patients with chronic hepatitis C than in normal controls (NCs). The expression of TLR2 did not show significant difference between NCs and chronic hepatitis C.
TLR2 stimulation enhanced the suppressive function of Tregs in chronic hepatitis C
We then compared the effect of TLR2 stimulation on the suppressive functions of CD3+CD4+CD25+CD127dim/− Tregs between NCs and chronic hepatitis C. We sorted highly pure CD4+CD25+CD127dim/− Tregs and CD4+CD25− Tresps subsets from NCs (n = 6) and chronic hepatitis C (n = 6). Tregs did not proliferate in response to anti-CD3/CD28 with or without Pam3Csk4 in either NCs or chronic hepatitis C (Fig. 2A). In contrast, Tresps from both NCs and chronic hepatitis C revealed proliferation in response to anti-CD3/CD28 (Fig. 2B). Moreover, Pam3Csk4 did not remarkably increase the proliferation in either group (Fig. 2B). There were no significant differences in the suppressive activities of Tregs between NCs and chronic hepatitis C without the stimulation of Pam3Csk4 (Fig. 2C–E). The presence of Pam3Csk4 significantly enhanced the suppressive capacity of Tregs in both groups (Fig. 2C–E). Moreover, this enhancement effect was more potent in chronic hepatitis C patients when compared with NCs at the tested Tregs:Tresps ratios of 1:4 (p = 0.041, Fig. 2D) and 1:8 (p = 0.0087, Fig. 2E). Furthermore, we also measured IL-10, IL-17, IL-22, and IL-23 productions in the supernatants of cultured cells at the Tregs:Tresps ratios of 1:4. Secretion of IL-10 was elevated in chronic hepatitis C (22.19 ± 8.33 pg/mL) when compared with NCs (13.49 ± 5.47 pg/mL) without the stimulation of Pam3Csk4 (p = 0.026, Fig. 3A). Treatment with Pam3Csk4 increased the production of IL-10 in both NCs and chronic hepatitis C (Fig. 3A). The effect in chronic hepatitis C (80.66 ± 34.35 pg/mL) was stronger than in NCs (43.19 ± 21.53 pg/mL) (p = 0.045, Fig. 3A). However, there were trends of elevation in IL-17, IL-22, and IL-23 expressions in NCs and chronic hepatitis C in response to Pam3Csk4 stimulation, and these differences failed to achieve significances (p > 0.05, Fig. 3B–D). The expression profile of these cytokines revealed similar trends in the supernatants of cultured cells at the Tregs:Tresps ratios of 1:1 or 1:8 (data not shown).
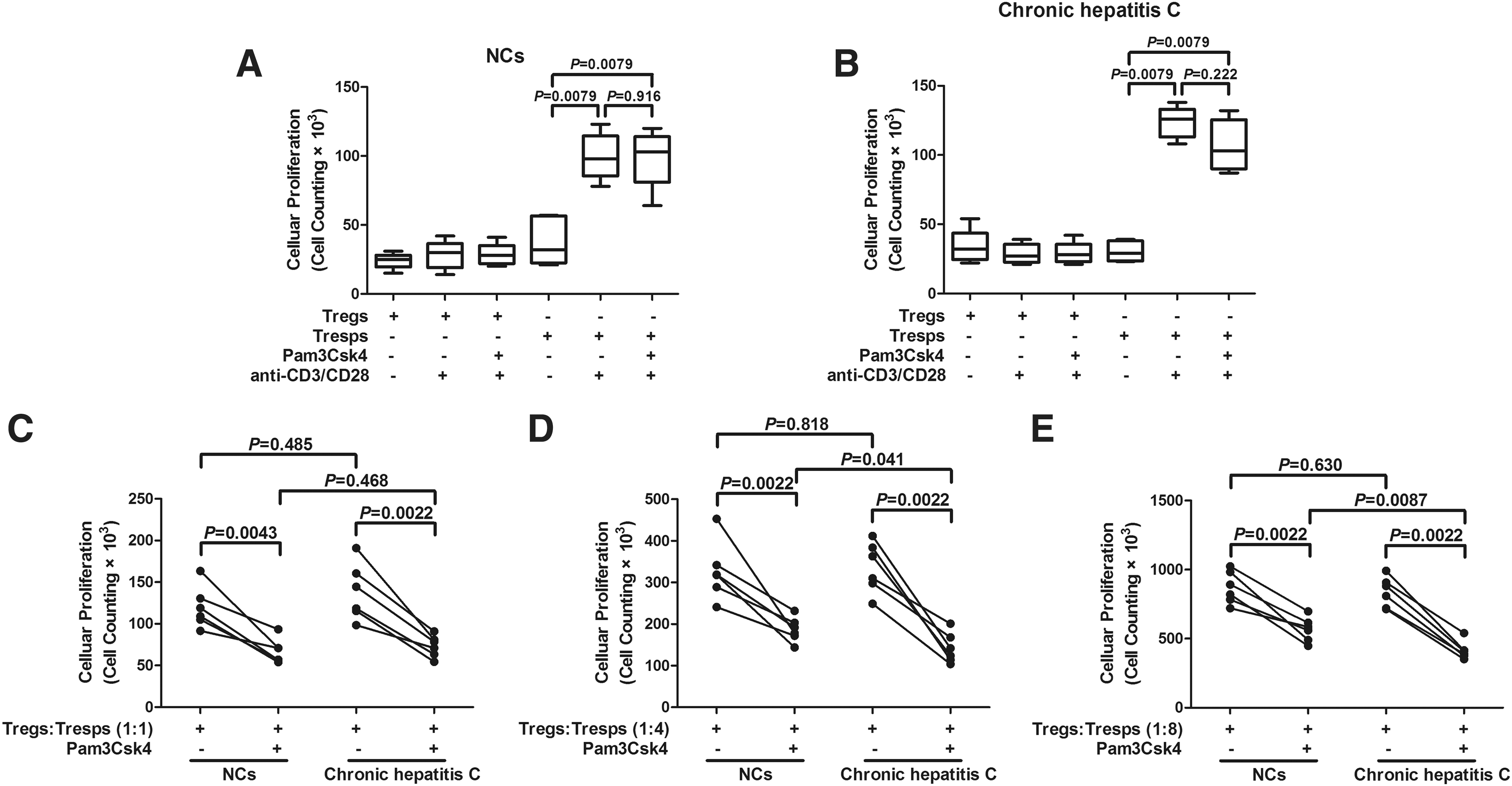
TLR2 stimulation enhanced the suppressive function of CD4+CD25+CD127dim/− regulatory T cells (Tregs) from both NCs and chronic hepatitis C patients. Purified CD4+CD25+CD127dim/− Tregs and CD4+CD25+ responder T cells (Tresps) from NCs

Levels of regulatory T cells (Tregs) and Th17-related cytokines in the supernatant of CD4+CD25+CD127dim/− Tregs and CD4+CD25− responder T cells (Tresps) cocultured system at the ratio of 1:4 from NCs (n = 6) and chronic hepatitis C (n = 6) in response to Pam3Csk4 stimulation.
TLR2 stimulation did not promote Th17 differentiation of Tregs from chronic hepatitis C patients
Th17 cells were demonstrated to play a pathogenic role in chronic viral infectious diseases (20,33), including hepatitis C (9,27). Previous study also revealed that TLR2 stimulation increased IL-17 production and enhanced a Th17 shift from Tregs in human (23). Thus, we analyzed the effect of TLR2 stimulation on the differentiation of CD4+CD25+CD127dim/− Tregs in chronic hepatitis C. Purified enriched CD4+CD25+CD127dim/− Tregs from both NCs (n = 7) and chronic hepatitis C (n = 11) were cultured for 4 days with or without Pam3Csk4 stimulation in a complete medium containing anti-CD3/CD28. Stimulation with Pam3Csk4 slightly enhanced the expressions of CCR4 and CCR6 on Tregs isolated from NCs (p = 0.018 and p = 0.011, respectively; Fig. 4A, B). However, Tregs, which isolated from chronic hepatitis C, did not elevate CCR4 and CCR6 expressions in response to TLR2 stimulation (p = 0.065 and p = 0.101, respectively; Fig. 4A, B). Moreover, the secretions of IL-17, IL-22, and IL-23 were also measured in the supernatants. IL-23 concentrations were under the limitation of detection in most cases. There were no notable differences in IL-17 and IL-22 expressions in response to TLR2 stimulation in either NCs or chronic hepatitis C (p > 0.05, Fig. 4C, D).
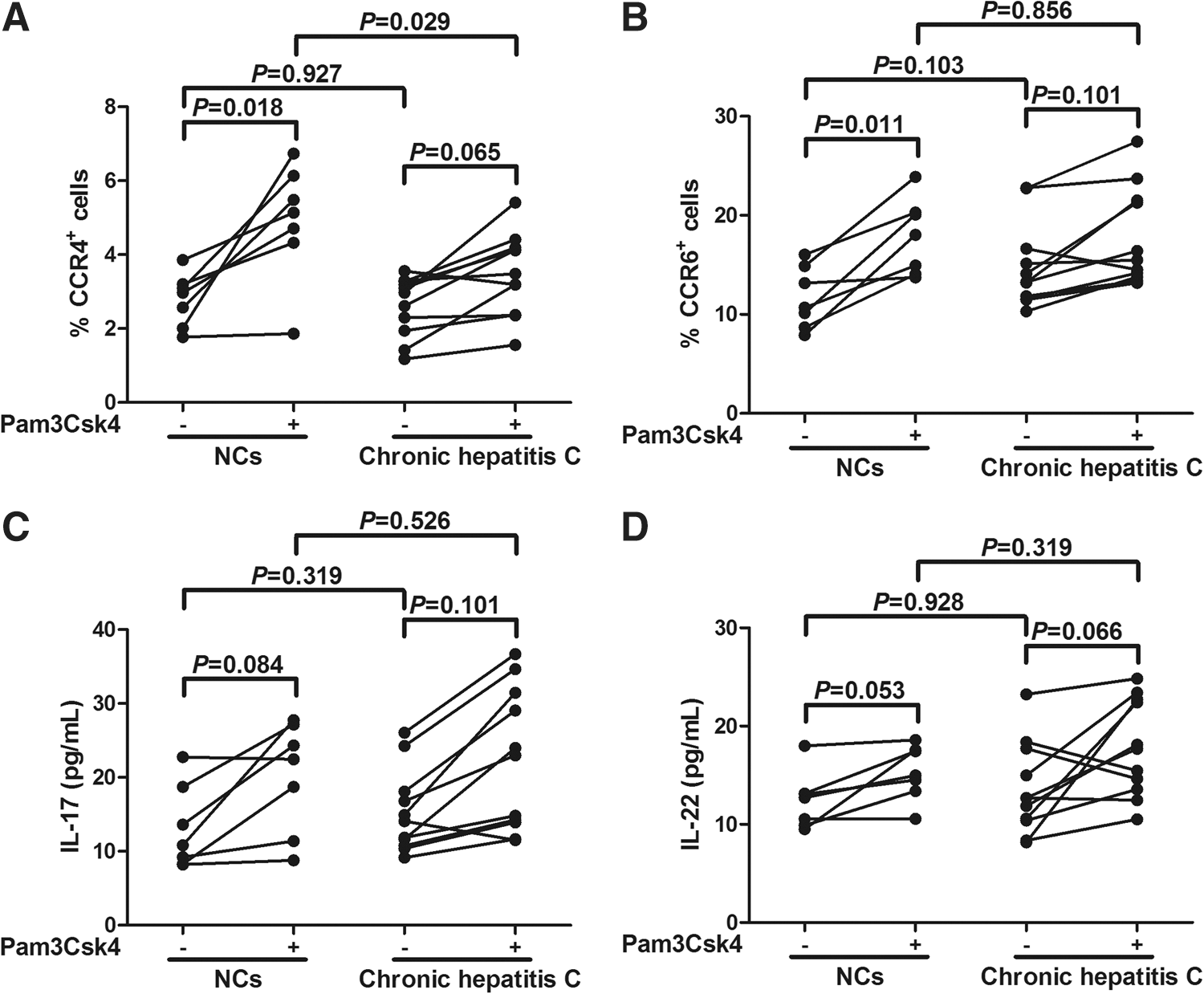
TLR2 stimulation did not enhance the expressions of CCR4 and CCR6 and productions of interleukin (IL)-17 and IL-22 by CD4+CD25+CD127dim/− regulatory T cells (Tregs) from NCs (n = 7) and chronic hepatitis C (n = 11). Purified CD4+CD25+CD127dim/− Tregs from NCs and chronic hepatitis C patients were cultured in the absence or presence of Pam3Csk4. Cultures were incubated for 4 days, and the cells were harvested and stained for the expression of CCR4
Effective anti-HCV therapy did not influence the TLR2-induced suppressive function of Tregs, but enhanced TLR2-mediated Th17 differentiation
All the 65 chronic hepatitis C patients received peg-IFN-α2a plus ribavirin therapy. HCV viral loads were determined at baseline and end of therapy. Fifty-three of the patients were responders to the therapy, and 12 were nonresponders. To assess the effect of anti-HCV therapy on the TLR2-mediated functions of Tregs and Th17 cells, PBMCs were isolated from 24 patients (including 12 responders and 12 nonresponders) at 48 weeks posttreatment. CD4+CD25+CD127dim/− Tregs were purified, and cocultured with CD4+CD25− Tresps at the ratio of1:4. There were no remarkable differences in the suppressive function of Tregs between responders and nonresponders. Pam3Csk4 stimulation significantly increased the suppressive activities of Tregs, however, the elevation effect did not reveal notable difference in either group (Fig. 5A). The secretions of related cytokines were also measured in the supernatants. IL-10 production was reduced in chronic hepatitis C patients with responders to therapy (18.13 ± 8.02 pg/mL) when compared with nonresponders (34.72 ± 21.37 pg/mL) (p = 0.031, Fig. 5B). Treatment with Pam3Csk4 enhanced the expression of IL-10 in both groups. However, there was no significant difference in IL-10 level in response to Pam3Csk4 stimulation between responders and nonresponders (p = 0.436, Fig. 5B). There were also elevations in IL-17, IL-22, and IL-23 in patients with responder therapy in response to Pam3Csk4 stimulation (p = 0.0024, p = 0.0032, and p = 0.0005, respectively; Fig. 5C–E). However, nonresponders did not show dramatic changes with Pam3Csk4 stimulation of these Th17 cytokines (p > 0.05, Fig. 5C–E).
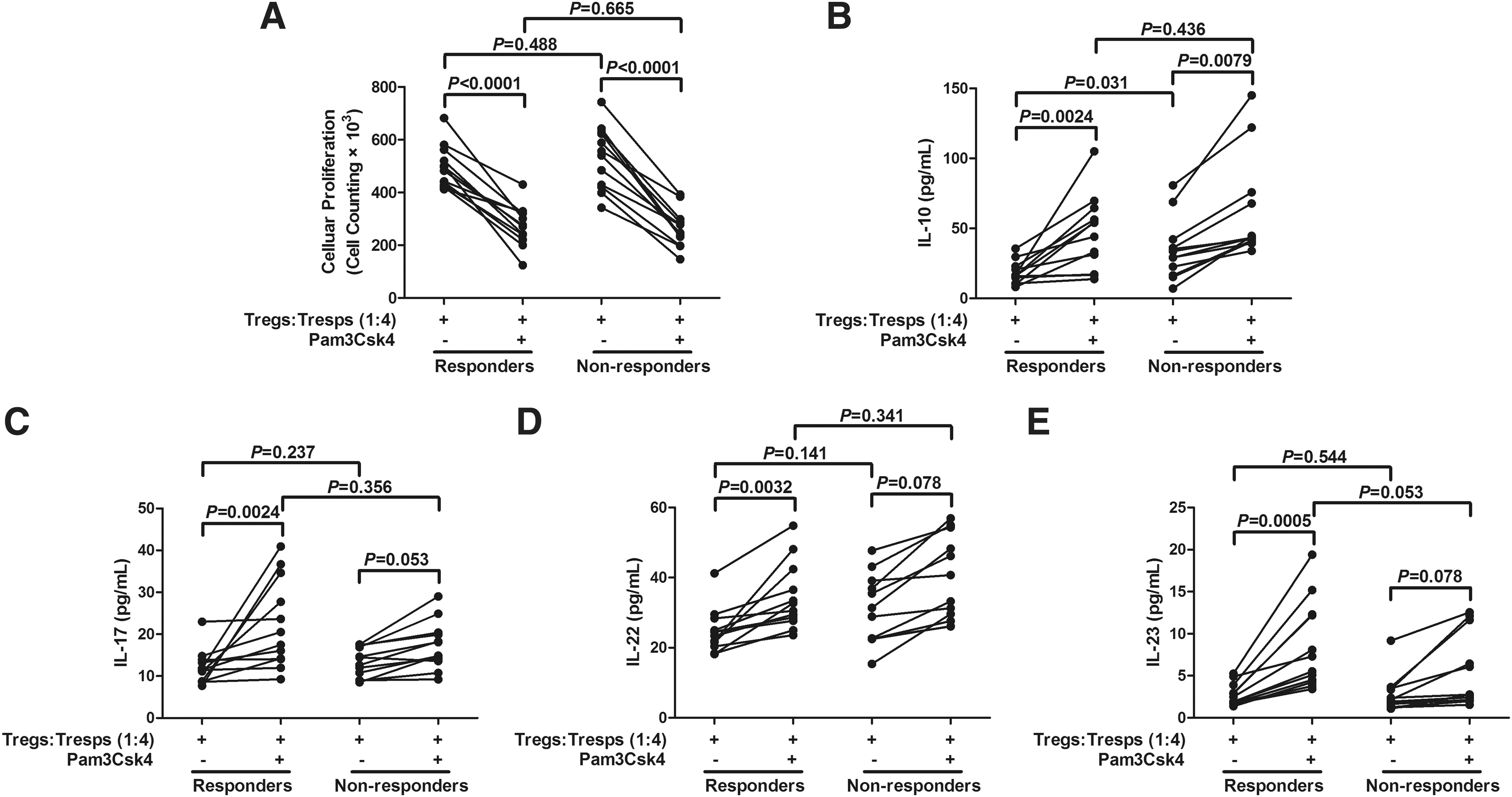
TLR2 stimulation enhanced the suppressive function of CD4+CD25+CD127dim/− regulatory T cells (Tregs) and Th17-related cytokine productions from responders to antiviral therapy (n = 12) and nonresponders (n = 12).
CD4+CD25+CD127dim/− Tregs purified from responders and nonresponders were also cultured alone with or without Pam3Csk4 stimulation. Interestingly, CCR4 and CCR6 expressions on Tregs were remarkably increased in response to TLR2 stimulation in responders (p = 0.001 and p = 0.069, respectively; Fig. 6A, B). However, nonresponders did not reveal significant CCR4 and CCR6 elevation on Tregs (p = 0.069 and p = 0.194, respectively; Fig. 6A, B). Similarly, IL-17 and IL-22 secretion were also notably increased in responders (p = 0.043 and p = 0.0002, respectively; Fig. 6C, D), but not in nonresponders (p = 0.078 and p = 0.056, respectively; Fig. 6C, D) in response to TLR2 stimulation.
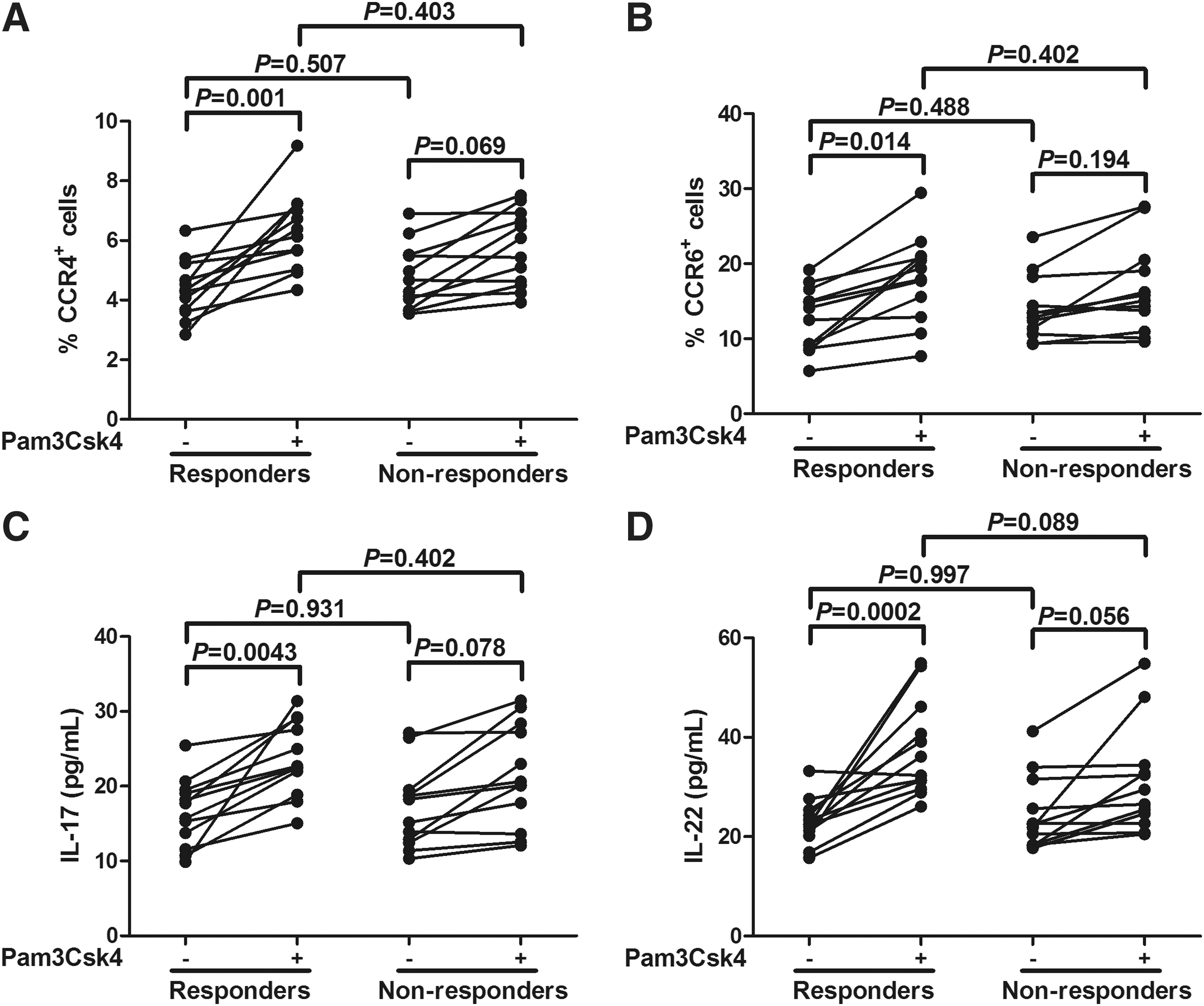
TLR2 stimulation enhanced the expressions of CCR4 and CCR6 and productions of interleukin (IL)-17 and IL-22 by CD4+CD25+CD127dim/− regulatory T cells (Tregs) from responders to antiviral therapy (n = 12) when compared with nonresponders (n = 12). Purified CD4+CD25+CD127dim/− Tregs from responders and nonresponders were cultured in the absence or presence of Pam3Csk4. Cultures were incubated for 4 days, and the cells were harvested and stained for the expression of CCR4
Discussion
In the current study, TLR2 was expressed in both Tregs and Th17 cells from NCs and patients with chronic HCV infection, revealing that these cells may be directly regulated by microbial molecular patterns, thus, linking innate signals and adaptive immune response. Furthermore, TLR2-induced enhancement of Tregs activity was found in both NCs and chronic hepatitis C. In contrast, TLR2 did not remarkably increase Th17 cell differentiation in both groups, however, effective anti-HCV treatment notably elevated TLR2-induced shift of Tregs toward Th17-like phenotype and function without loss of suppressive function in chronic hepatitis C. The regulation of adaptive immune cells by innate stimuli in chronic HCV infection may partly explain how systemic infections influence phases of immune activation.
TLR2 was expressed at higher levels by Tregs of chronic hepatitis C patients than Tregs of NCs. Consistent with this observation, purified CD4+CD25+CD127dim/− Tregs from chronic hepatitis C patients were more susceptible to Pam3Csk4 induction. However, controversy remained as to the effect of TLR2 stimulation in regulation of Tregs function. Nyirenda et al. revealed that stimulation of TLR2 in patients with relapsing-remitting multiple sclerosis reduced Tregs suppressive function, and TLR2 induced loss of suppressive function, which was more prominent in Tregs isolated from patients compared with NCs (22). In contrast, Nawijn et al. showed that Pam3Cys treatment induced an expansion of Tregs in lungs, which resulted in long-term protection against allergic asthma in mice model (21). Furthermore, Zhang et al. demonstrated that exposure of Tregs to Pam3Csk4 enhanced the immunosuppressive function and decreased cellular proliferation in patients with chronic hepatitis B (34). The present results showed that TLR2 stimulation increased CD4+CD25+CD127dim/− Tregs suppressive function by reduction of cellular proliferation and elevation of IL-10 production. The Pam3Csk4-induced enhancement of Tregs suppressive function was more highlighted in chronic hepatitis C patients than NCs. Although coculture of Tregs:Tresps at 1:1 did not make a significant difference in the proliferation of NCs and chronic hepatitis C in response to TLR2 stimulation, this is partly due to the high percentage (50%) of Tregs in the culture that played an important nonspecific immunosuppressive role. Moreover, effective anti-HCV therapy did not influence the TLR2-induced suppressive function of Tregs since responders to peg-IFN-α2a plus ribavirin combination therapy did not reveal higher cellular proliferation and less IL-10 secretion than nonresponders upon Pam3Csk4 stimulation. These findings demonstrated that CD4+CD25+CD127dim/− Tregs responded directly to TLR2 signaling, a mechanism that was likely to contribute to viral persistence and chronic HCV infection.
Regulation of Th17 cell differentiation in HCV infection was not fully elucidated. Previous studies on autoimmune and infectious diseases demonstrated a direct immunomodulatory role of TLR2 in Th17 cell differentiation. He et al. revealed that high-mobility group box 1 promoted Th17 cell differentiation through upregulation of TLR2 on CD14+ monocytes in patients with rheumatoid arthritis (14). Zhao et al. also showed that Pam3Csk3 directly activated Th17 cell response without antigen stimulation in HBV infection (35). We found that TLR2 expression in Th17 cells did not reveal a significant difference between NCs and chronic hepatitis C. Pam3CSK4 treatment did not enhance the productions of Th17-secreting cytokines and Th17 differentiation in either group. Interestingly, effective anti-HCV therapy remarkably enhanced the TLR2-mediated Th17 cell differentiation, which manifested by secretion of higher levels of IL-17, IL-22, and IL-23 in responders to peg-IFN-α2a and ribavirin treatment. The increased susceptibility of Th17 cells might be induced by peg-IFN-α2a therapy since recent study on HCV/HIV coinfection indicated that Th17-secreting IL-23 promoted IFN-α responsiveness, which potentially led to activation of Th17 cells (24).
Tregs secreted little or no IL-17 without antigen stimulations or pathogen infections, such as thymus in humans or in naive mice under specific pathogen-free conditions (31). Voo et al. reported a large number of IL-17-producing CD4+CCR6+FoxP3+ Tregs in human peripheral blood and lymphoid tissue. The CD4+IL-17+FoxP3+ T cells exhibited strong suppressive function through a cell-to-cell contact mechanism (31). Further study revealed that Tregs produced IL-17 during oropharyngeal candidiasis and inflammatory bowel disease in a TLR2/MyD88 signaling-dependent manner. Thus, stimulation of TLR2 promoted IL-17 secretion in Tregs, even absence of T cells receptor signals in vitro (6). However, Nyirenda et al. showed that TLR2 stimulation promoted the differentiation of human Tregs into a Th17-like phenotype by enhancing IL-17 and IL-6 production, elevating RORc, and reducing FoxP3 expression (23). This resulted in the loss of inhibitive function in Tregs (23). We found that Tregs isolated from chronic hepatitis C produced similar levels of IL-17 and IL-22 in response to Pam3Csk4 stimulation. Moreover, effective antiviral therapy induced more IL-17 and IL-22 secretion and higher expression of CCR4 and CCR6 by both Tregs and enriched CD4+ T cells upon TLR2 stimulation. Importantly, the shift phenotype toward Th17 did not reduce the suppressive function of Tregs. This is consistent with the previous findings that Tregs expanded with TLR2 agonists were capable of suppressing CD4+ T cells, even upon IL-6 withdrawal (6). This implied that the HCV infection may have inhibited the function of IL-17 production by Tregs in response to TLR2 stimulation, but exhibited more suppressive function, which in turn contributed to persistent HCV infection.
Conclusion
In summary, we demonstrated that stimulation of TLR2 modulated the Tregs/Th17 cell balance in patients with chronic hepatitis C. HCV infection regulated Tregs/Th17 cells through TLR2 stimulation by inducing Tregs to produce IL-10 and enhancing cell-to-cell contact inhibition of effector T cells, resulting in viral persistence. Inhibition of HCV replication by effective antiviral therapy led to the promotion of IL-17-secreting phenotypic shift upon TLR2 stimulation. Such mechanisms would enable deliberate manipulation targeting Tregs/Th17 differentiation pathway to treat chronic HCV infection.
Footnotes
Acknowledgment
The authors thank all the volunteers who generously participated in this study.
Author Disclosure Statement
No competing financial interests exist.