
Abstract
Continued evolution of highly pathogenic H5N1 viruses causing high mortality in humans obviates need for broadly cross-reactive vaccines. For this, hemagglutinin (HA) inducing specific protective antibodies, highly conserved nucleoprotein (NP), and ectodomain of matrix (M2e) protein, either singly or in combination, were evaluated in BALB/c mice. Recombinant HA and NP (baculovirus system) and M2e (synthetic peptide) and 3 adjuvants, that is, liposomes, Mw (heat killed Mycobacterium w), and alum were utilized for the homologous virus challenge. Additional immunogens included liposome-encapsulated HA/NP proteins and corresponding DNAs. Mice groups received two doses of respective formulations given at 3-week intervals and challenged intranasally with 100LD50 of H5N1 virus strain. Dynamics of weight loss, lung viral load, titres of IgG-anti-HA, NP, and M2e antibodies (ELISA), and IgG-subtype analysis was done. Two doses of all the formulations led to 100% seroconversion against the immunogens evaluated (100% seroconversion after the first dose in majority). Antibody titres against the components were dependent on the adjuvant and combination. HA-driven Th2 response with all the adjuvants, balanced Th1/Th2 response to NP protein, and Th2-bias with alum were noted. Low anti-M2e antibody titres did not allow subtype analysis. On challenge, complete protection was observed with Mw-HA, alum-HA+NP, Lipo-HA+NP+M2e, alum-HA+NP+M2e, and HA-DP formulations with 12-fold, 8-fold, 720-fold, 17-fold, and no reduction, respectively, in lung viral load. In conclusion, the results identify several adjuvant-immunogen combinations conferring 100% protection in mice that need further evaluation in higher animals.
Introduction
T
Based on hemagglutinin (HA) gene sequence analyses, H5N1 viruses are classified in several clades, the number increasing with the emergence of new variants. On account of the observed drug resistance in avian influenza strains (5,16), vaccination remains the most effective public health intervention for influenza epidemics, as well as pandemic. India experienced the first H5N1 outbreak in poultry in 2006 (24). In 2011, a new clade emerged (2.3.2.1) that was distinct from the previously circulating clade 2.2 virus (20). Although specific anti-HA antibodies are protective, evolution of unique strains of unknown HA specificity demands use of broadly reactive vaccines for viruses without prior immunity in humans. For this, the highly conserved nucleoprotein (NP) and M2 proteins represent the ideal candidates. Immunization with NP was shown to produce CTL responses, while the anti-NP antibodies were associated with viral clearance (15). M2e (ectodomain of M2 protein) has been targeted for universal vaccine development because of accessibility to antibodies and remarkably conserved nature (12). Monoclonal antibodies recognizing the SLLTEVET epitope could inhibit influenza A virus replication in MDCK cells (31), while passive transfer of M2e-specific human monoclonal antibodies inhibited virus replication in the lungs of mice challenged with H5N1 virus (30).
Different DNA-based and vectored vaccines were evaluated as broadly cross-protective vaccines (7,8,23,26). Moderate protection was noted against low-dose H5N1 virus challenge in ferrets with DNA/rAd5 vaccines encoding NP, M2, and NP+M2 (26). NP+M2 DNA vaccine with Vaxfectin adjuvant could just prolong appearance of symptoms and death in ferrets (14).
Previous reports of clinical studies indicate that low immunogenicity of H5N1 in humans can be compensated by the addition of adjuvants to vaccine formulations (3,17,18) and is probably essential to improve cross-clade immunity in humans. We report comparative evaluation of liposome, Mw, and alum adjuvants in generating protective immune response in mice immunized with various combinations of recombinant HA and NP proteins and synthetic M2e.
Material and Methods
Viruses
The H5N1 influenza A virus used in this study was isolated at the National Institute of Virology (H5N1-Navapur-A/chicken/India/33487/2006 influenza virus, clade 2.2).
Construction of recombinant plasmids and expression/purification of proteins
HA and NP genes were amplified from viral RNA and cloned in pVAX1 and pFASTBac1 expression vectors (Invitrogen) according to manufacturer's instructions. The primers used for HA gene: forward primer (5′-CAT CGG ATC CCA ATC TGT CAA AAT GGA GAA AAT AGT GCT TCT TCT TG-3′) and reverse primer (5′-CAT CCT CGA GAA CTA CAA TCT GAA CTC ACA AAT TTA AAT GCA AAT TCT G-3′) and for NP gene, forward primer (5′-CAT CGG ATC CGA CAT CAA CAT CAT GGC GTC TC-3′) and reverse primer (5′-CAT CCT CGA GTT AAT TGT CAT ACT CCT CTG CAT TG-3′). pFASTBac1-HA and NP DNA were transformed into maximum efficiency DH-10Bac competent cells (Invitrogen). rBac was transfected in Spodoptera frugiperda (SF9) cells according to the standard protocol, and 72 h post transfection, pellets and supernatants were harvested, and the presence of the proteins was confirmed by ELISA and immunoblotting. Protein stocks of HA and NP were produced by harvesting the infected SF9 cells at 120 and 72 h postinfection, respectively.
Recombinant HA and NP proteins were purified from infected SF9 cells using lentil lectin affinity chromatography (11) and cation exchange chromatography, respectively, and assayed by ELISA/western blot using specific mouse monoclonal antibodies (HyTest Ltd.). The biological activity of the purified protein was confirmed by hemagglutination (HA) test using 0.5% turkey RBCs. For NP, the lyzed cell pellets were centrifuged and supernatants were subjected to cation-exchange chromatography using Mono S 10/100 GL column (GE healthcare). The elution was done by MES buffer containing 1M NaCl. Endotoxin levels in the purified HA and NP protein preparations were measured using LAL Chromogenic Endotoxin Quantitation Kit (Thermo Scientific).
Endotoxin-free bulk plasmid preparations
HA and NP cloned-DNAs were purified using EndoFree Plasmid megaprep columns (Qiagen). Agarose gel and spectrophotometric (NanoDrop, Thermo Scientific) analyses were done to check the quality, purity, and quantity of the plasmids.
M2e
Synthetic M2 peptide (M2e) was obtained from INBIOS S.r.l, Italy. The sequence of this peptide was “SLLTEVETPTRNEWECRCSDSSD”.
Adjuvants
Mw was purchased from Cadila Pharmaceuticals, India. Formulations containing Mw were prepared by adding adjuvant and immunogens (1:1 v/v ratio) to make a total of 100 μL dose per mouse. Thus, 100 μL of each vaccine formulation per mouse contained 50 μL of Mw adjuvant (2.5 × 108 cells) and 50 μL of immunogens (volume made up with phosphate buffer saline). Formulations containing Al(OH)3 adjuvant were prepared by adsorbing 1 μg of the antigen on 3.25 μg of Al(OH)3 (Sigma chemicals) by vortexing at a low speed for 1 h at RT followed by centrifugation at 5,000 rpm for 5 min at RT. The pellet was resuspended in 0.01M PBS, pH 7.2 and diluted to the required dose at the time of immunization. Liposomes were prepared as described previously (27). Immunogens were mixed with the liposome suspension in the mass ratio of 1:200.
Experiments in mice
The study was approved by the Institutional Animal Ethics Committee and conducted as per the guidelines. Experiments requiring infection of mice with H5N1 viruses were done at the BSL 3+ biosafety containment facility of the institute. The work in BSL3+ laboratory was approved by the Institutional Biosafety Committee.
Mice immunizations
Female inbred BALB/c mice aged 6–8 weeks (n = 10 per group) were immunized with two doses of respective formulations given at 3-week intervals (Table 1). Five microgram each of recombinant proteins HA, NP, M2e peptide, and naked DNA (HA/NP) were used as per the vaccine formulations. Groups of mice were immunized intramuscularly with 100 μL of respective formulations, 50 μL per quadriceps muscle. Mice were bled before immunization, 2 and 3 weeks postimmunization, and 10 days post second dose. Sera were stored at −20°C until tested. Control mice received PBS.
IgG titers against HA, NP, and M2e in mice immunized with two doses of different formulations at 3-week intervals.
The immunized groups where 100% seroconversion was achieved after second dose.
DPΦ, DNA plus corresponding protein; HA, hemagglutinin; NP, nucleoprotein; ND, not done.
Virus challenge
Ten days post second dose, immunized and control mice (n = 10/group) were challenged intranasally with 50 μL of 100LD50 homologous virus challenge. Mice were weighed daily for 14 days and analyzed for disease (i.e., weight loss, ruffling fur, and inactivity).
Lung viral load
Forty-eight hours post challenge, three mice/group were sacrificed and harvested lungs were stored in RNAlater solution at −80°C until tested. RNA was extracted from lung homogenate using QIAamp Viral RNA Mini Kit (Qiagen) according to the manufacturer's instructions. Viral RNA copy number was determined by VLA TaqMan Influenza A/H5/H7 Detection Kit (Applied Biosystems).
Serologic assays
For the detection (seroconversion) and titration of IgG-anti-HA antibodies, recombinant protein-based ELISA was used as described earlier (2). Same protocol was used for the detection and titration of anti-NP and anti-M2e antibodies with following modifications. For NP, 1 μg/well protein was used for coating. For M2e, 250 ng/well was used for coating at 4°C for 16 h and the incubation periods were increased to 60 min (sample) and 45 min (conjugate). The cutoff values for IgG positivity for all ELISAs were: 3 × mean OD values of preimmune sera (negative controls). Samples showing OD values ≥ cutoff value were considered positive for anti-HA, anti-NP, and anti-M2e IgG antibodies and taken as evidence of seroconversion with the respective antigens. IgG subtype analysis was done as described previously (6).
Statistical analysis
Student's t-test and ANOVA with Tukey's post hoc test were performed using SPSS 20 to compare antibody titers among groups of mice. Kaplan–Meier survival analysis was performed in GraphPad Prism software. The differences at p < 0.05 were considered significant.
Results
PCR amplification and cloning of HA and NP gene
The sequences of complete HA and NP genes cloned in the pVAX1 and pFASTBac1 expression vectors were 100% identical with the sequence of the isolate, GenBank accession no. EF362418.1 (HA) and EF362421.1 (NP). Figure 1 depicts purified HA and NP proteins and immunoreactivity with specific monoclonal antibodies. Endotoxin levels in the purified HA and NP preparations were less than 10EU/mg.
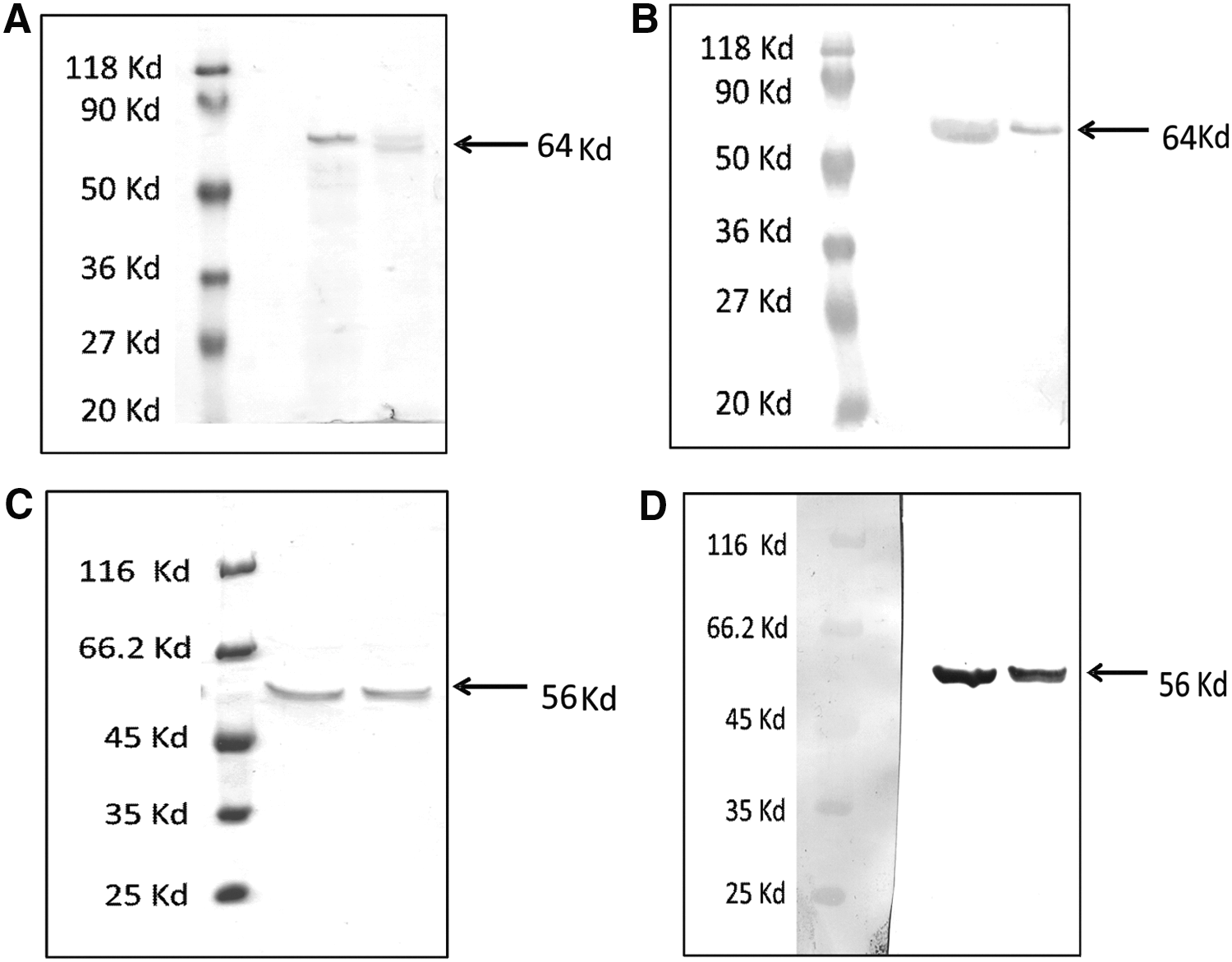
SDS PAGE and immunoblot of HA and NP proteins. Recombinant HA and NP proteins were expressed in Sf9 cells, and purified proteins were analyzed by SDS PAGE (
Antibody response in mice
Seroconversion rates
Two doses of all the formulations led to 100% seroconversion (ELISA) against all the formulations evaluated (Table 1). After the first dose, except for 90% seroconversion recorded in the mice receiving lipo-HA-NP-M2e, all the other formulations showed 100% seroconversion to anti-HA antibodies. For NP, the addition of M2e to HA+NP combination enhanced seroconversion from 80% to 100%, irrespective of the adjuvant type. When all the three immunogens were used in combination, 100% seroconversion was recorded after first dose with all the three adjuvants. Mice immunized with protein and corresponding DNA encapsulated in liposomes showed 100% seroconversion after first dose. Anti-M2e antibodies were detected only after the second dose except when all the immunogens were administered with alum.
HA adjuvanted with Mw, liposome, and alum
Mw-HA formulation yielded highest IgG-anti-HA titers (13,718 ± 2,375) that were comparable with liposome-HA (8,444 ± 2,664). Alum-induced antibody titres were significantly lower (6,400 ± 746) than the Mw formulation (p < 0.005) and comparable with liposome-HA. All the formulations induced significantly higher anti-HA-IgG1 than IgG2a, suggestive of predominant Th2 response (Fig. 2).
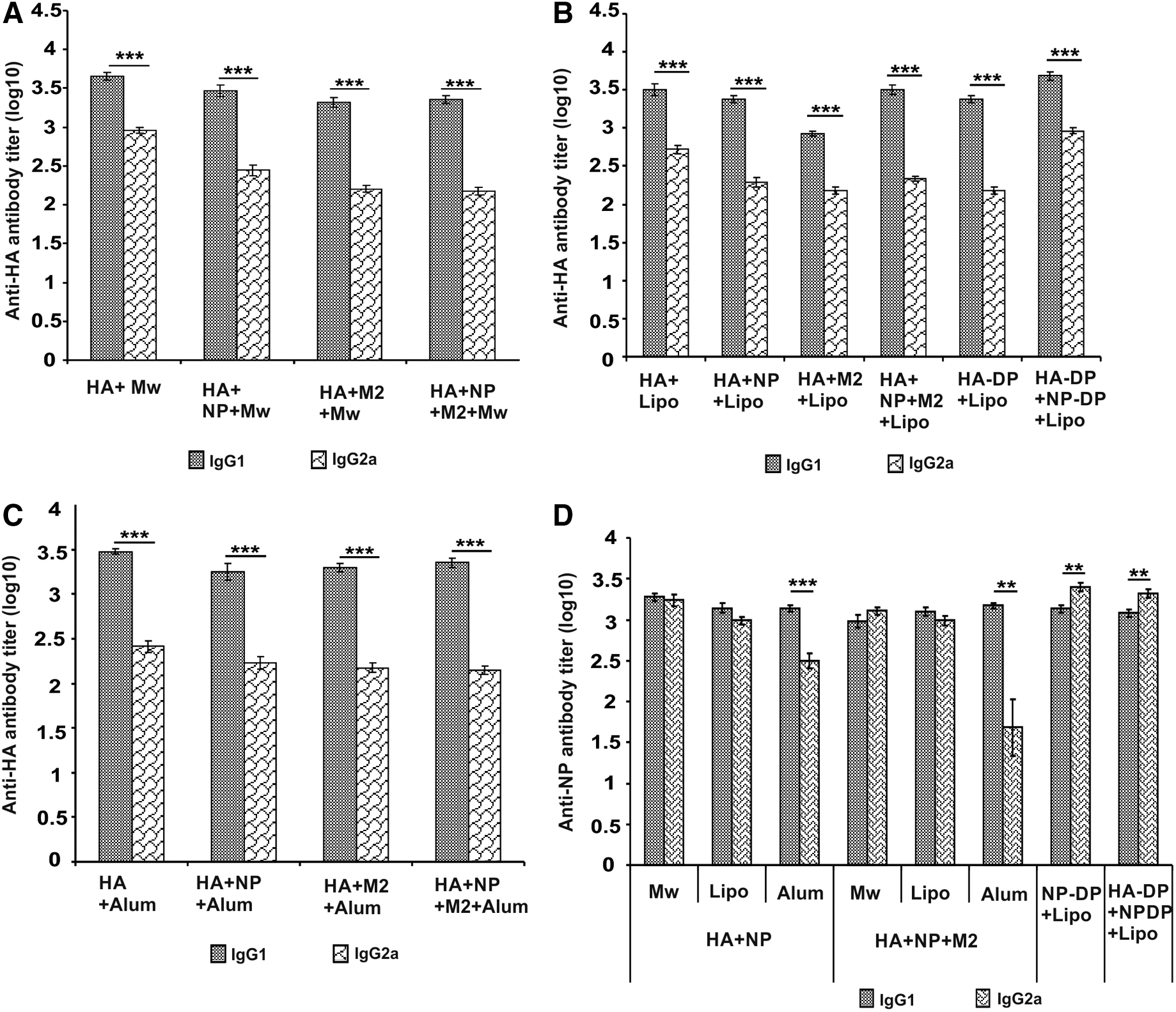
Anti-HA and anti-NP IgG isotype titers at prechallenge time point. Geometric mean HA-specific
HA + NP adjuvanted with Mw, liposome, and alum
The pattern of anti-HA-IgG titres did not change after the addition of NP (Table 1). Both Mw and liposome induced comparable high anti-HA titres (9,050 ± 2,579 and 7,879 ± 2,742, respectively), while significantly lower titres were induced with alum (4,525 ± 1,013) than Mw (p < 0.05). IgG-anti-NP titers were significantly higher with Mw (4,850 ± 522) than liposome (3,200 ± 582) and alum formulations (2,599 ± 477) (p < 0.05). Liposome and alum elicited equivalent titers.
HA + M2e adjuvanted with Mw, liposome, and alum
Addition of M2e to HA led to reduction in anti-HA titres (Table 1). With this formulation, the anti-HA titres were comparable with Mw and alum, liposome-induced titres reducing significantly (p < 0.001). A significant reduction (p < 0.05) in IgG-anti-HA titres was noted when M2e peptide was added to alum-HA. Overall, all the three adjuvants induced comparable low levels of IgG-anti-M2e titres.
HA+NP+M2e with Mw, liposome, and alum
Liposome formulations yielded highest HA-specific IgG antibody titers compared to Mw and alum (p < 0.05) (Table 1). IgG-anti-HA titers induced by liposome and alum were significantly improved by the addition of NP to the respective HA+M2e preparations (p < 0.05). IgG-anti-HA titers elicited by Mw were comparable with alum. IgG-anti-NP titers produced by all the three adjuvants were comparable. The addition of M2e to alum-HA+NP increased IgG-anti-NP (p < 0.05). IgG-anti-M2e titres with Mw-adjuvant were comparable with alum and significantly lower than the liposome group (p < 0.05). Liposome and alum groups induced comparable antibody titres.
Liposome encapsulated HA/NP DNA+Protein (DP) combinations
The anti-HA titres recorded for all the formulations were comparable to lipo-HA group (Table 1). However, significantly higher titres were induced when HA-DP+NP-DP formulation was administered compared to HA-DP immunization (p < 0.005).
As far as anti-NP antibodies are concerned, both NP-DP and HA-DP+NP-DP formulations induced higher titers than with combination of Lipo-HA+NP proteins (p < 0.005). No difference was noted when liposome-adjuvanted NP-DP and HA-DP+NP-DP groups were compared.
Isotyping
In all the groups receiving HA as one of the components of vaccine formulation with all the three adjuvants, anti-HA-IgG1 isotype titers were significantly higher than IgG2a, suggestive of predominant Th2 response (Fig. 2A–C). Mw and liposome-adjuvanted HA+NP and HA+NP+M2e groups exhibited equivalent titers of NP-specific IgG1 and IgG2a antibodies, indicative of balanced Th1/Th2 response to NP protein (Fig. 2D). With alum, IgG1 titers were higher than IgG2a (Th2 bias). Liposome-encapsulated NP+DNA formulations elicited strong Th1 response. Due to the low anti-M2e-IgG titers, isotype analysis was not done.
Protection offered by different formulations against H5N1 virus challenge
HA alone
Mice immunized with Mw-adjuvanted formulation were completely protected (100% survival) with no appreciable weight loss, whereas alum-HA led to 86% survival (12.9% ± 3.2% weight loss) (Fig. 3). With liposome, 57% protection was recorded (12% ± 4.1% weight loss).

Morbidity (body weight loss) and Percent Survival kinetics. At day 10 post dose-2, mice vaccinated with HA + adjuvant formulations
Addition of NP to HA
Protection offered by this combination differed for all the adjuvants compared to HA-alone formulations. In this category, 100% protection (17.3% ± 1.3% weight loss) was provided by alum while an increased survival from 57% to 83% was recorded with liposome. However, 100% protection offered by HA-Mw was reduced to 71% with HA-NP-Mw.
Addition of M2e to HA
This formulation did not yield 100% protection with any of the adjuvants. In fact, for both Mw and alum adjuvants, addition of M2e reduced survival from 100% and 86% to 43% and 57%, respectively, while no added advantage was noted with liposome.
Combination of HA, NP, and M2e
Liposome and alum-adjuvanted formulations provided 100% protection. However, with Mw formulation, survival was reduced to 43%.
DNA+Protein combinations
Complete protection was conferred by HA-DP combination while NP-DP could confer 29% survival. Surprisingly, when both the formulations were combined, 86% mice were protected.
Viral load in the lungs of infected mice
At 48 h postinfection, the control mice exhibited 9 × 104 RNA copies/mL lung suspension, whereas no viral RNA could be detected in the mice group receiving lipo-HA+NP formulation (Table 2). Same combination with Mw and alum resulted in 708- and 8-fold reduction in the viral load. In the Mw-HA group, viral load decreased by 12-fold and all the mice survived with 5.6% ± 2.7% weight loss. An 8- and 17-fold reduction in viral load and complete protection were noted with alum-HA+NP and alum-HA+NP+M2e formulation, respectively. Interestingly, liposome-HA+DNA group exhibited complete protection in the presence of high viral load (9 × 104). Liposome-HA+NP+M2e providing complete protection showed 720-fold reduction.
DP*, DNA plus corresponding protein.
Among the partially protected groups, lipo-HA-DP+NP-DP group showed 7-fold reduction. In the lipo-HA and alum-HA groups, partial protection was associated with 10- and 14-fold reduction in viral RNA, respectively.
Discussion
We evaluated three adjuvants in generating protective immune response when immunized with HA alone or in combination with NP and M2e, the two conserved viral proteins known to confer broad cross-reactive immunity against infection with different clades of H5N1. Being an agent with pandemic threat, various strategies are being evaluated to develop universal cross-protective H5N1 vaccines requiring minimum manufacturing time. Use of recombinant proteins is one such effort. In this respect, the HA protein is of special importance, anti-HA antibodies being protective against the viral infection (2). It was, therefore, decided to evaluate strain-matched HA, singly or in combination with NP and M2e, as possible vaccine candidates. Success with hepatitis E virus (HEV) (27) prompted us to assess liposome-encapsulated HA and NP formulations containing protein and corresponding DNA.
Mw is approved for use as an adjuvant (Immuvac/Cadi-05) in leprosy treatment (25) and was shown to induce potent Th1 immune response (1). On the contrary, we documented a predominant Th2 response with liposome-encapsulated recombinant proteins of HEV, Hepatitis B virus, Hepatitis C virus, and chikungunya virus (10,13,27). We selected adjuvants predominantly eliciting Th1 or Th2 responses and alum as the gold standard. Incidentally, this is the first report of use of Mw for influenza vaccine development.
On account of the induction of virus-specific neutralizing response, HA has been the choice for vaccine development (19,22). It is, therefore, logical to expect optimum response with this protein. Immunogenicity of nonadjuvanted HA was evident by 100% seroconversion post dose-1, with anti-HA-IgG titers of 1131.4 ± 151.2 that was differentially enhanced by different adjuvants. Overall, efficacy of the formulations was adjuvant dependent, Mw-HA conferring complete protection, whereas liposome-HA and alum-HA formulations provided 57% and 86% protection, respectively. Interestingly, addition of HA-DNA to the liposome formulation led to 100% protection reconfirming efficacy of this approach (9).
We further compared anti-HA-IgG titres and degree of protection obtained with different HA formulations. With Mw adjuvant, highest antibody titres correlated with 100% protection. Despite comparable anti-HA antibody titers, degree of protection varied drastically with liposome. Lipo-HA and lipo-HA+DNA groups exhibited 57% and 100% protection, respectively. The observed complete protection may be the result of cellular immune response induced by the DNA component of the vaccine. Studies have shown that despite low HI antibodies, DNA vaccine encoding H5 HA induced protection in mice suggesting crucial role of cell-mediated immune response elicited by DNA vaccination (14). Although Mw was evaluated as a Th1 inducer, a consistent Th2 response with the HA-formulations suggests antigen-driven modulation of immune response by HA, aTh2-inducing protein. The effect on anti-NP and anti-M2e titres could not be determined as we did not examine NP or M2e as distinct immunogens.
To enhance the efficacy of HA formulations, we added the conserved NP and/or M2e. No universal additive effect was seen and the efficacy of combination formulations was variable and adjuvant driven. Addition of NP was beneficial for alum (86% to 100% protection) and liposome (57% to 86%), whereas a distinct reduction (100% to 71%) was exhibited by Mw. Overall, Mw with higher anti-HA and anti-NP titres was not protective, whereas liposome with comparable anti-HA titres and lower anti-NP titres showed improved efficacy while alum with lower anti-HA and NP titres provided complete protection. These results suggest that the protection was not solely dependent on antibody response to both HA and NP. As NP is shown to act through CTL response (7), differential modulation of such responses by the adjuvants may have contributed to the observed differences. Although we did not determine HI titres, it may be noted that the ELISA used during the study correlated well with HI (2).
None of the formulations containing HA and M2e led to 100% protection. With all adjuvants, addition of M2e alone to HA led to decline in anti-HA-IgG titers (p < 0.001), reduced virus clearance in the lungs of mice at 48 h post challenge (Table 2), and efficacy. Importantly, we used M2e without any carrier protein. A weakened protection by the addition of M2 peptide to oil in water emulsion adjuvanted split H3N2 virus is noteworthy (33).
Among the liposome formulations containing DNA and corresponding proteins, HA was protective, while NP could protect ∼1/3rd mice suggesting inferiority of NP alone. These results are at variance with an earlier study reporting complete protection of mice immunized with liposome encapsulated 10 μg NP or M2 or 5 μg each of (NP+M2) against heterosubtypic H5N1 virus challenge (28).
Alum, the gold standard, did emerge as an excellent adjuvant for recombinant antigens as both HA+NP and HA+NP+M2e formulations provided complete protection. Protection offered by HA (86%) was enhanced by the addition of NP and NP+M2e (100%). NP appeared to be a better antigen as addition of M2e to HA led to lowering of IgG-anti-HA titres and concomitant reduction in survival (57%).
Taken together, these findings demonstrate that protective response against H5N1 vaccines varies with adjuvant and the viral proteins. Understanding immunologic basis for these differences will be the key to delineate the mechanism(s) of adjuvant action and formulating strategies to modulate the immune response in favor of the host.
In conclusion, our study provides comparative efficacy of HA used singly or in combination with NP and M2e adjuvanted with Mw, liposome, or alum adjuvants and identifies Mw-HA, Lipo-HA+NP+M2e, HA-DP, alum-HA+NP, and alum-HA+NP+M2e as formulations yielding complete protection against lethal homologous challenge. These results need to be extended to heterologous H5N1 challenge and confirmed by studies in higher animals.
Footnotes
Acknowledgments
This work was funded by the Department of Biotechnology, Govt. of India. The authors thank the Director, National Institute of Virology, Pune for all the facilities. The authors sincerely thank Dr. S.D. Pawar for providing virus stocks for challenge experiments. The authors appreciate the assistance of Mr. Prakash Sarje, Prakash Jawalkar, and Prasad Babar in BSL-3 work. The authors also thank Mr. Shirish Vaidya and Mr. Satish Ranwade for technical assistance.
Authors’ Contributions
N.B.I: Planning and execution of experiments, including mice immunizations and experiments in BSL3+ facility, data analysis, and article preparation. R.G.V.: Planning and execution of experiments, including mice immunizations. K.A.: Expression of recombinant proteins. K.S.S.: Animal experimentation in BSL3+ facility. K.S.L.: Monitoring expression of recombinant proteins. V.A.A.: Planning the study and experiments, data analysis, and article preparation.
Author Disclosure Statement
No competing financial interests exist.