
Abstract
In the developing countries, Hepatitis E virus (HEV) is a predominant cause of sporadic acute hepatitis in adults and waterborne epidemics leading to high mortality in pregnant women. Vaccine development mainly focuses on the structural capsid protein open-reading-frame-2 (ORF-2) of the virus. We successfully evaluated liposome-adjuvanted recombinant neutralizing epitope protein (rNEp), a part of ORF-2, 458-607aa, in mice and rhesus macaques. We compared immune response to adjuvants alone, rNEp alone, or adjuvanted with liposome (lipo-rNEp)/alum (al-rNEp) in mice following intramuscular administration of two doses of 5 μg each. IgG anti-HEV titers (enzyme-linked immunosorbent assay), immunophenotyping (flow cytometry, CD3+CD4+, CD3+CD8+, CD11c+, CD11b+, CD19+ cells; costimulatory markers CD80, CD86, MHC-I, MHC-II, and early activation marker CD69), and levels of Th1/Th2 cytokines (IL-2/IFN-γ/IL-4/IL-5 and additionally IL-1β/IL-6/IL-10/TNF for early time points) were determined at early (4/12/24-h postdose-1) and later time points (2 weeks post-both doses). IgG anti-HEV titers were higher in the lipo-rNEp group than al-rNEp post-both doses (p < 0.05). At early time points, cell type proportions were comparable at the site of injection; IL-Iβ levels increased in lipo-rNEp, 24 h, while IL-6 levels rose in lipo-rNEp/al-rNEp/alum-alone groups, 4 h, compared to controls. In the draining lymph nodes (DLNs), CD11c+CD86+ cells increased at 24 h in liposome-alone/lipo-rNEp groups. A rise in the CD11c+CD69+ cells was noted in the lipo-rNEp group compared to other groups (p < 0.05). Cytokine levels in the spleen/sera remained unchanged in all the groups (p > 0.05). At 2 weeks postdose-2, CD11c+MHC-II+/CD11b+MHC-II+ cells increased in the spleen in the lipo-rNEp and al-rNEp groups, respectively. In the DLNs, CD19+MHC-II+ cells increased in rNEp/al-rNEp/lipo-rNEp groups post-both doses and CD11c+CD86+ cells in the lipo-rNEp group. A balanced Th1/Th2 response was evident in the lipo-rNEp, while a Th2 bias was noted in al-rNEp. Different immune response gene clustering patterns were noted in uncultured spleens from immunized mice and cultured-stimulated splenocytes. In conclusion, lipo-rNEp is a better immunogen, works through dendritic cells, and elicits a balanced Th1/Th2 response, while alum functions through macrophages and induces a Th2 response.
Introduction
H
Inability of the virus to grow in culture systems or small laboratory animals makes recombinant technology the best possible option for vaccine development. Most commonly targeted is the open reading frame-2 (ORF-2) capsid protein. Antibodies against ORF-2 in humans and infected animals are long lived and cross-reactive among diverse HEV genotypes, and neutralize HEV in vitro (1,29,31,35).
So far, two vaccine candidates using parts of ORF-2 successfully completed clinical trials. One was a 56 kDa (HEV495, 112-607aa) protein produced in insect cells, while the other was a bacterially expressed protein of 239aa (HEV239, 368-606aa) (36,46). HEV239 vaccine (Hecolin®) is now commercially available for use only in China. Our vaccine candidate (recombinant neutralizing epitope protein [rNEp]) is the 150aa neutralizing epitope protein, 458-607aa, represented in both the vaccine formulations completing clinical trials. ORF-2-encoded protein forms the viral capsid through its homodimeric subunits (domain E2, aa 394–606 and domain E2s, aa 455–602). The HEV neutralizing antibody recognition site was mapped to the E2s (I) domain (26). Both HEV495 and HEV239 form virus-like particles (VLPs), contain immunodominant epitopes, and are highly immunogenic when administered with alum as the adjuvant (33,40,42 –45). In fact, an addition of 26 amino acids at the N terminus of E2 protein was shown to improve immunogenicity when administered as a single dose of VLP preparation (27). rNEp is the shortest form that can dimerize and has the major neutralizing epitopes (43).
During the course of vaccine development, we evaluated ORF-2 and neutralizing epitope containing region (NE) in mice and rhesus monkeys, using several approaches such as DNA, DNA prime/protein boost and encapsulation of DNA, and the corresponding proteins in liposomes (3,12). Furthermore, liposome-encapsulated rNEp alone or with hepatitis B surface antigen (HBsAg) could induce comparable antibody titers in mice and rhesus monkeys (37, unpublished observations). Recently, a single dose of rNEp was shown to be well tolerated and highly immunogenic in pregnant mice (19). Importantly, IgG-anti-HEV antibodies elicited in mice (12) and rhesus monkeys (unpublished observations) were shown to bind HEV in an enzyme-linked immunosorbent assay (ELISA)-based in-vitro assay. As we first assessed the DNA and corresponding protein formulations as vaccine candidates, liposome was the adjuvant of choice for encapsulation. However, rNEp being the smallest protein considered as vaccine candidate, a comparison of alum, the time-tested adjuvant and liposome in generating immune response were undertaken.
Materials and Methods
Immunogen preparation
The rNEp was cloned, expressed, and purified as described previously (3,12,19,37). In brief, the HEV strain (PM2000, genotype-1, GenBank accession No. DQ459342) collected during an outbreak in Lonawala, Western India, in 2000 was used to express rNEp in the bacterial system (pET-15b vector, 5,708 bp; Novagen) and purified using Nickel-chelating resin (Invitrogen). Endotoxin levels of protein in the final formulations were <10 EU/mg of protein by the LAL (Limulus amebocyte lysate) test (Pierce LAL Chromogenic Endotoxin Quantitation Kit; Thermo Fisher Scientific). rNEp adjuvanted with aluminum hydroxide (al-rNEp) was prepared by adsorbing rNEp on to aluminum hydroxide (alum) (Sigma-Aldrich) in the mass ratio 1:3, antigen:adjuvant. Liposomes were prepared by the dehydration–rehydration method, and rNEp adjuvanted with liposomes (lipo-rNEp) was prepared by mixing rNEp with the liposomes in the mass ratio of 1:200, antigen:adjuvant as described previously (19,37).
Mice immunization
The study was approved by the institutional animal ethics committee (IAEC) and experiments were carried out according to the guidelines. Table 1 represents experimentwise allotment of mice groups. Female balb/c mice, 6–8 weeks, were immunized with 5 μg of rNEp, al-rNEp, and lipo-rNEp formulations. Two doses were administered 2 weeks apart intramuscularly in the quadriceps muscles. Mice were sacrificed at 2 weeks postdose 1 and 2 weeks postdose-2. For studies on immune response at early time points, phosphate-buffered saline (PBS) alone, rNEp alone (5 μg), alum alone, liposome alone, al-rNEp (1:3), and lipo-rNEp (1:200) were injected and immune response at 4, 12, and 24 h was studied. Mice were bled through retro-orbital plexus and sera collected were stored at −20°C until tested for ELISA. Spleens/draining lymph nodes (DLNs)/muscles were harvested and either processed immediately for surface staining or stored in −80°C for cytokine estimation/gene expression studies.
alum, aluminum hydroxide adjuvant; al-rNEp, rNEp adjuvanted with aluminum hydroxide; lipo-rNEp, rNEp adjuvanted with liposomes; PBS, phosphate-buffered saline; rNEp, recombinant neutralizing epitope protein.
IgG anti-HEV titers by ELISA
Anti-HEV positivity and titers were determined by a baculovirus expressed recombinant ORF-2 protein (rORF-2p)-based ELISA as described previously (3,4,12,19,37). Briefly, Sf9 (Spodoptera frugiperda) cell lysate containing rORF-2p was diluted in 50 mM carbonate buffer (pH 9.5) and used for coating (100 μL/well) 96-well microtiter plates (MaxiSorp Nunc). Following coating at 37°C for 2 h, the wells were blocked with a blocking solution at same temperature for 30 min. After washes, mice sera were added to wells at appropriate dilutions in the blocking solution. Incubation was continued for 30 min. A 1:10 dilution of preimmune sera served as negative control. Horseradish peroxidase (HRP)-conjugated goat anti-mouse IgG (Sigma Chemicals) was used as a detector antibody. The enzymatic reaction with substrate (O-phenylenediamine and urea peroxide) was stopped by the addition of 4N H2SO4 and the absorbance measured at 492 nm. A serum sample was considered to be reactive when the optical density (OD) value was greater than or equal to thrice mean OD value (ELISA cutoff) of preimmune sera (negative control). The reciprocal of the highest dilution with an absorbance greater than or equal to the ELISA cutoff was considered as the HEV-specific antibody titer.
For isotyping, sera samples from a group were pooled. Isotype detection was carried out using the same protocol with an additional step. Reaction with test serum was followed by incubation at 37°C for 30 min with goat anti-mouse IgG isotype antibodies (IgG1 and IgG2a) (Sigma Chemicals). HRP-conjugated rabbit anti-goat IgG (Sigma Chemicals) was used as the detector antibody.
Cytokine estimation in sera/muscle/spleen
Cytokine estimation (IL-2, IFN-γ, IL-4, IL-5, and TNF) was carried out using the BD Th1/Th2 Mouse Cytokine Kit (BD Biosciences). For estimation of IL-6, IL-1β, and IL-10, cytokine ELISAs were performed according to the manufacturer's instructions (BD Biosciences). The muscles and spleen were cut into small pieces and digested with 0.05% type II collagenase (Roche) in RPMI (Roswell Park Memorial Institute; Invitrogen) medium for 30 min at 37°C under constant agitation. The cell suspension was centrifuged and the supernatants were stored in −80°C until further analysis.
Surface staining of splenocytes/DLN cells
Spleens/DLNs were teased gently with forceps to produce a single-cell suspension in incomplete RPMI and staining was carried out as described previously (19). Briefly, 1X BDFACS lysing solution (1%) was added to lyse erythrocytes. Following three to four washes with RPMI, cells were centrifuged at 1,000 rpm for 10 min. After the final wash, cells were filtered through a 70-μm cell strainer and viability was checked using 0.1% trypan blue. One million cells were labeled with anti-mouse fluorochrome (FITC/PE/APC/APC-Cy7.7) (BD Pharmingen)-labeled antibodies against CD3 (17A2), CD4 (GK1.5), CD8 (53–6.7) (T-cell markers), CD11b (M1/70) (macrophage marker), CD11c (HL3) (dendritic cell marker), CD49b (DX5) (NK-cell marker), and CD19 (ID3) (B cell marker) cells. Costimulatory markers CD80 (B7-1), CD86 (B7-2), MHC-II (M5/114.15.2), MHC-I (H2Kb), and early activation marker, CD69 (H1.2F3), on cells were studied by dual/triple staining. For T-cell activation, the anti-mouse T-lymphocyte activation cocktail was used (BD Pharmingen). Isotype antibodies were included. Following addition of antibodies, cells were kept in dark for 30 min. Cells were washed with PBS and suspended in 400 μL 1% paraformaldehyde. A minimum of 20,000 events were acquired for every sample on BD FACS ARIAII flow cytometer (BD Biosciences). Analysis was carried out using the BD FACS DIVA software (BD Biosciences).
Estimation of cytokines in rNEp stimulated splenocytes by cytometric bead array
To assess the Th1 (IL-2 and IFN-γ)/Th2 (IL-4 and IL-5) cytokines in the supernatants of splenocytes and DLN cells, the Cytometric Bead Array (CBA) Kit (BD Biosciences) was used as per the manufacturer's protocol and the assay was set up as reported previously (34). Briefly, 1 million cells were seeded in RPMI supplemented with 2% fetal bovine serum per well and were stimulated with 1 μg of rNEp. Cell supernatants were collected after 72 h and stored in −80°C till tested. Concanavalin A (Con A)-stimulated (2.5 μg/well) and unstimulated cells served as positive and negative controls, respectively.
Gene expression analysis by TaqMan low-density array
Total RNA extraction (Ambion RiboPure kit; Thermo Fisher Scientific) and gene expression analysis (TaqMan array mouse immune panel; Applied Biosystems) were carried out from frozen spleen samples as described previously (32). The TaqMan low-density array (TLDA) card was loaded on to the 7900HT Fast Real Time PCR system (Applied Biosystems). Five hundred nanograms of RNA was used for complementary DNA (cDNA) synthesis, and cDNAs from spleens of naive mice were used as calibrators. The 18s gene was kept as endogenous control. Relative quantification (RQ) values below 0.5 were considered as downregulation and values >2 were considered as upregulation.
Gene expression from stimulated splenocytes (from immunized mice)
Spleens were processed immediately to obtain single-cell suspensions. One million cells were stimulated with 1 μg rNEp. 4 h and 24 h poststimulation, cells were centrifuged at 1,000 rpm for 10 min at 4°C. RNA extraction was carried out and processed for TLDA, as mentioned earlier.
Statistical analysis
Student's t-test and one-way analysis of variance with post hoc Tukey's test were performed using the GraphPad Prism 5.0 (GraphPad Prism, Inc.). A p-value <0.05 was considered significant. Nonresponders in each group were included in the analysis.
Results
Dose/route optimization
We first compared the anti-HEV antibody response of different doses (100 ng, 1 μg, 5 μg and 10 μg) of alum- or liposome-adjuvanted rNEp administered by subcutaneous and intramuscular routes. For both adjuvants, the intramuscular route was superior (data not shown) and used for subsequent experiments. Figure 1A and B depict seroconversion rates following immunization with two doses of the formulations. At lower doses (100 ng and 1 μg), a distinct difference was noted between the adjuvants, liposome inducing earlier and higher seroconversion rates. At 5 and 10 μg antigen concentrations, both adjuvants led to 100% seroconversion after first dose. Based on these results, a 5 μg dose was considered appropriate and used to compare different parameters in mice immunized with rNEp alone, adjuvant alone, or adjuvanted rNEp (al-rNEp or lipo-rNEp).
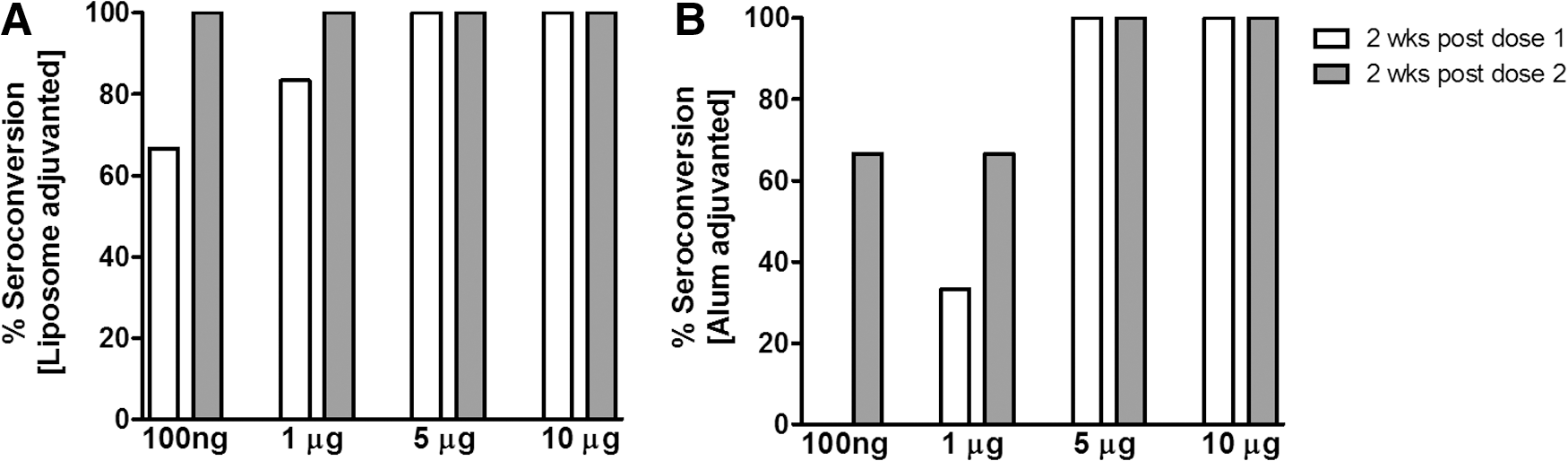
Seroconversion rates in mice immunized intramuscularly with
IgG anti-HEV titers and subtype analysis
IgG anti-HEV antibodies were not detected in the protein-alone group after first dose, while the geometric mean titer (GMT) post-second dose was 24 ± 1.3. In the alum-adjuvanted group, antibody titers after first (251 ± 54) and second (317 ± 43) doses were comparable (p > 0.1). lipo-rNEp elicited significantly higher antibody titers at 2 weeks post-second dose (1,131 ± 179) when compared to 2 weeks post-first dose (513 ± 66) (p < 0.005). After each dose, anti-HEV titers were higher in the lipo-rNEp than in al-rNEp group (p < 0.05 and <0.005, respectively). For both adjuvants, no significant decline was noted at 1 month post-second dose, while in the protein-alone group, antibody titers diminished (p < 0.05) (Fig. 2A). Anti-HEV-IgG subtype analysis showed that both the adjuvanted formulations favored Th2 response (titers of IgG1 higher than IgG2a, Fig. 2B).

IgG anti-HEV titers and subtype analysis in immunized mice.
Immunophenotyping and cytokines at early time points 4, 12, and 24 h postimmunization
In the muscles at the site of injection, activation of cell types was not recorded as evidenced by comparable levels of all the cell surface markers studied in any of the groups compared to controls (CD3+CD4+: 6–10%, CD3+CD8+: 5–9%, CD3+CD69+, CD19+CD69+: 4–8%, CD19+MHC-II+: 5–10%, CD11c+: 0–1.1%, CD11b+MHC-II+: 6–9%, CD3-CD49b+:2–4%, and CD11b+CD69+: 4–6%, p > 0.05).
A significant rise was noted in IL-1β levels in the liposome-alone and lipo-rNEp groups at 24 h when compared to the controls and other groups (p < 0.05 for all). Raised IL-6 levels (p < 0.05) were recorded for alum-alone, al-rNEp, and lipo-rNEp groups at 4 h compared to the controls (Fig. 3A, B). IFN-γ, IL-2, IL-4, and IL-5 were undetectable, while IL-10 (10–12.6 pg/mL) and TNF (10–14.4 pg/mL) remained comparable to the controls (p > 0.05).
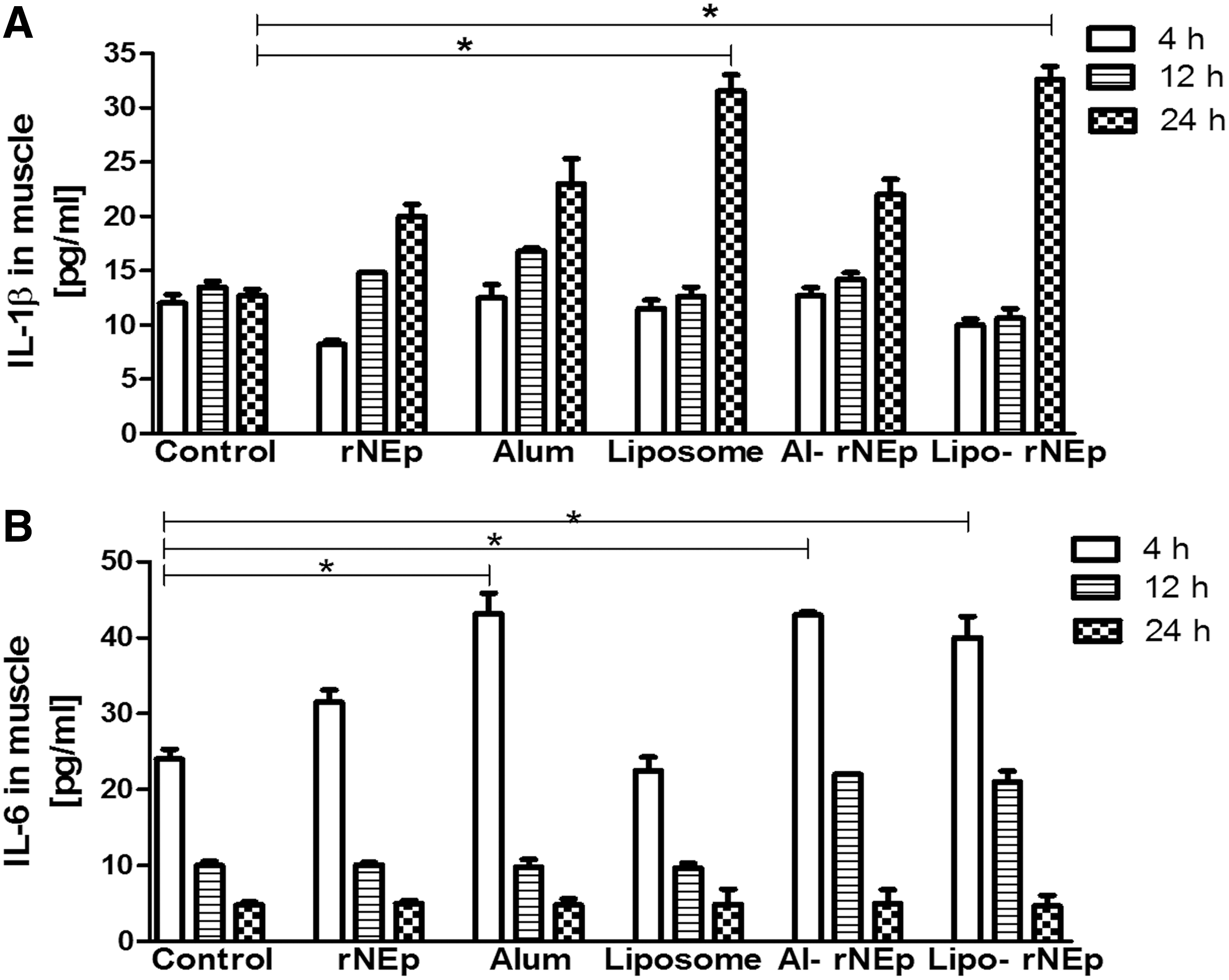
Cytokine levels of
In the DLNs, no significant changes were noted for any of the surface markers in the rNEp-alone, alum-alone or al-rNEp groups (CD3+CD4+: 45–50%, CD3+CD8+: 22–25%, CD3+CD25+: 4–8%, CD3+CD4+CD69+: 2–5%, CD3+CD8+CD69+: 0–2%, CD19+MHC-II+: 2.5–4%, CD19+CD69+: 4–8%, CD19+CD86+: Nil, CD19+CD138+: 7–9%, CD11c+MHC-II+: 0.5–1%, CD11c+CD69+: 0.2–0.8%, CD11c+CD80+: 0.4–0.8%, CD11b+CD69+: 4–6%, MHC-I: 3–3.5%, CD3-CD49b+CD69+: 4.5–5.2%, p > 0.05 for all). At 24 h, expression of the costimulatory molecule CD86 on CD11c cells increased in both liposome-alone and lipo-rNEp groups (p < 0.05). However, no difference was noted between the liposome-alone and lipo-rNEp categories (p > 0.05) (Fig. 4A).
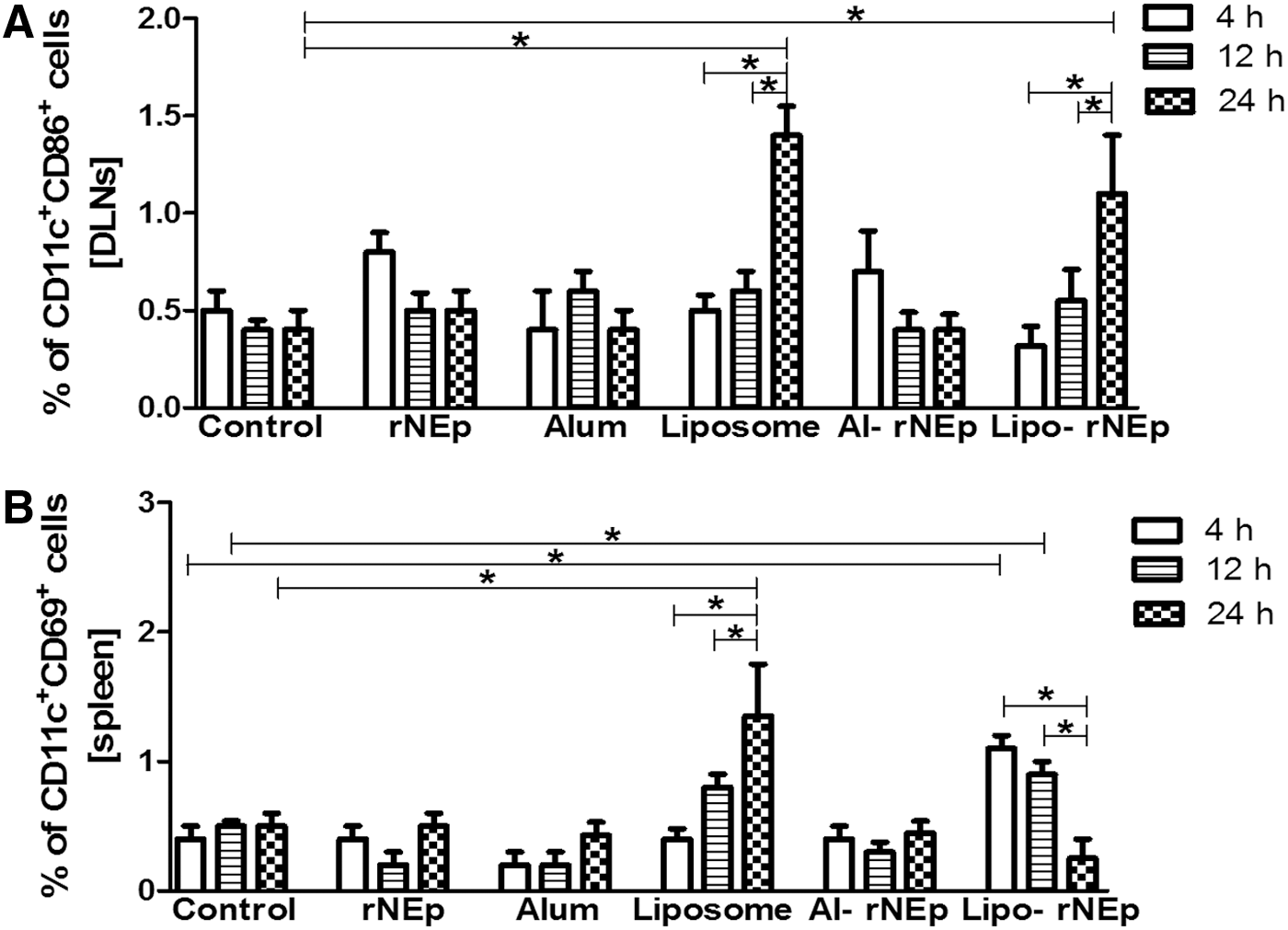
Percentage of dual positive CD11c cell types in DLNs and spleen at early time points
In the spleen, a rise in CD11c cells was noted at 24 h postdose in liposome-alone and lipo-rNEp groups. In the liposome-alone group, CD11c+CD69+ cells increased at 24 h (p < 0.05), while lipo-rNEp led to an earlier activation of dendritic cells (DCs) as indicated by higher levels at 4 and 12 h (p < 0.05) (Fig. 4B). The other markers in the rNEp-alone, alum-alone, and al-rNEp groups were comparable to the controls at all the time points (p > 0.05). Th1, Th2, and proinflammatory cytokines in the sera and spleens were comparable to PBS controls at respective time points in each of the groups (p > 0.05).
Immunophenotyping and cytokines at 2 weeks post-first and post-second doses
Draining lymph nodes
At 2 weeks post-first and post-second dose, a significantly higher expression of MHC-II on CD19 B cells was noted in the rNEp, al-rNEp, and lipo-rNEp groups when compared to the controls (p < 0.05, Fig. 5A). A higher number of CD11c+CD86+ cells were recruited in the liposome-adjuvanted group after both doses (p < 0.05, Fig. 5B), with no difference within the groups. Other markers were comparable to the controls (p > 0.05).
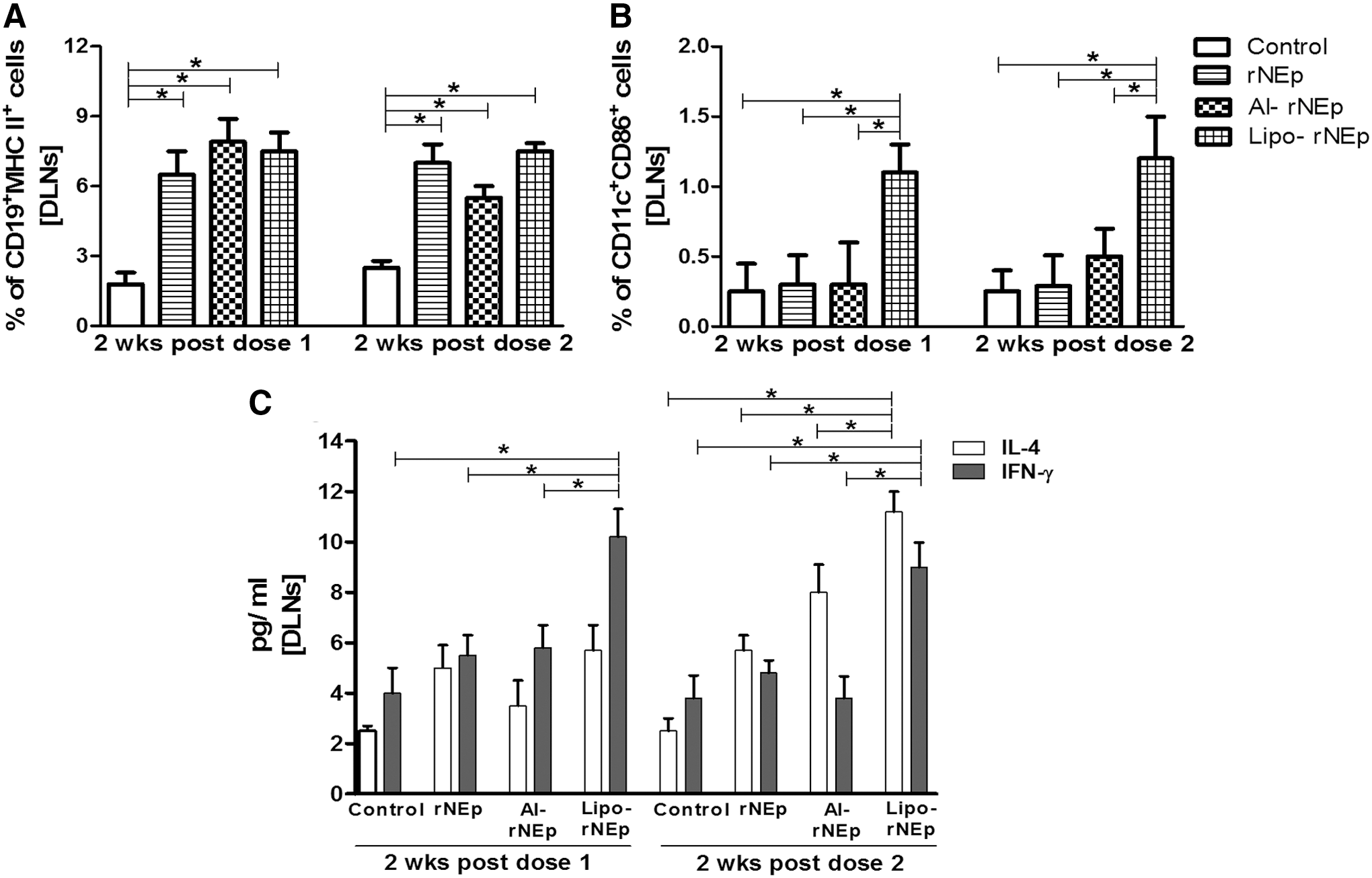
Percentage of cell types and cytokine estimation in the DLNs post-both doses.
In the lipo-rNEp group, IFN-γ levels were significantly higher after both doses than in the other groups (p < 0.05), while IL-4 levels increased post-second dose, compared to the controls (p < 0.05, Fig. 5C). No changes were noted in the al-rNEp group (p > 0.05). IL-2 and IL-5 levels were below the detection limits.
Spleen
A significant increase in the total population of CD11c cells was noted in the lipo-rNEp group after both doses (2.3–2.6% postdoses vs. 0.8–1% in controls) (p < 0.05). Post-second dose, a higher proportion of CD11b+MHC-II+ cells and CD11c+MHC-II+ cells were recorded in the al-rNEp and lipo-rNEp groups, respectively (p < 0.05, Fig. 6A). Expression levels of CD3+CD4+, CD3+CD8+, CD3+CD25+, CD19+MHC-II+, CD19+CD80+, CD19+CD86+, CD19+CD138+, CD11c+CD80+, CD11c+CD86+, MHC-I, CD3-CD49b+ remained comparable to the controls (p > 0.05).
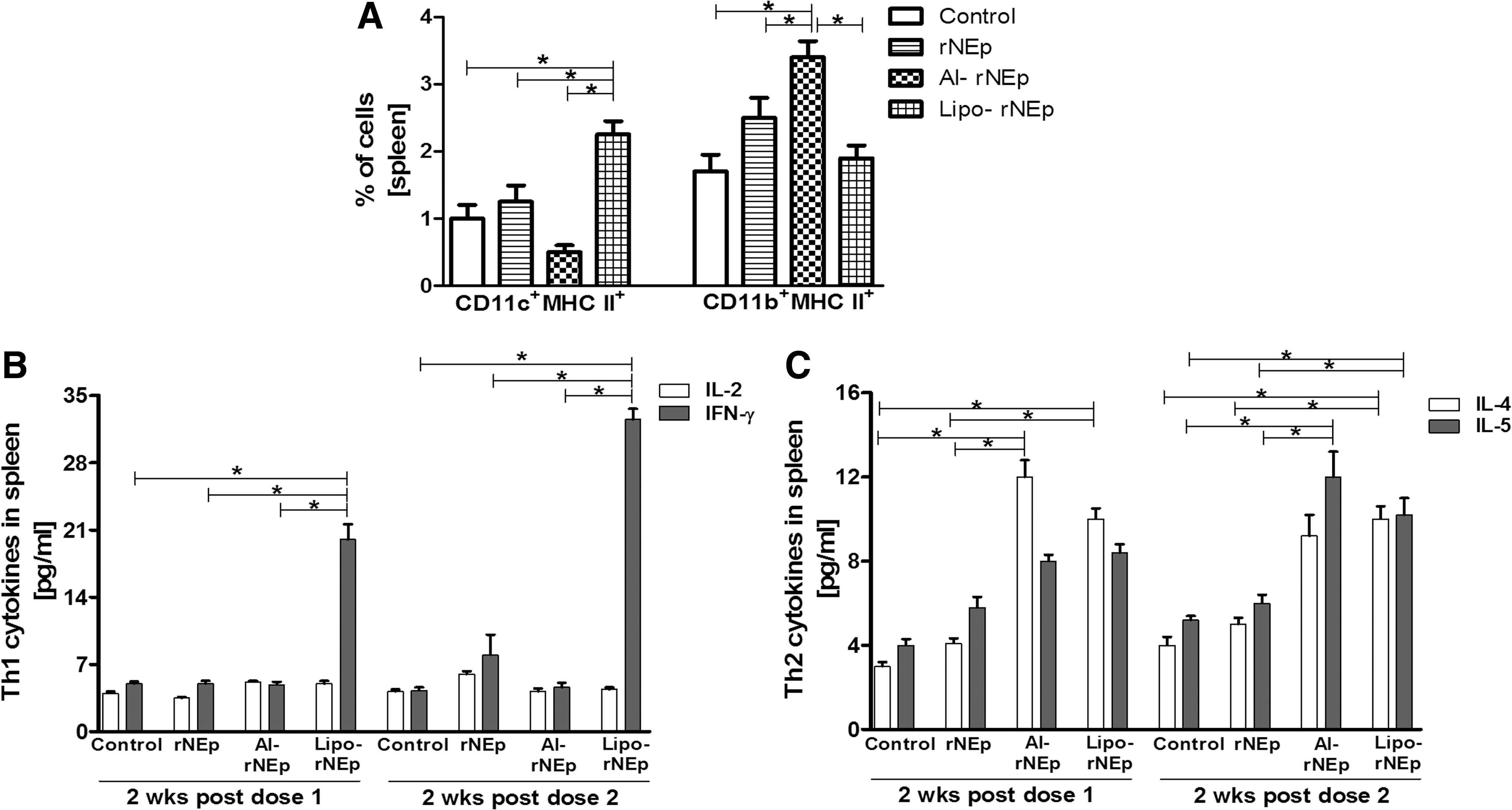
Percentage of cell types (2 weeks post-second dose) and cytokine estimation (post-both doses) in the spleen.
At 2 weeks postdose, the levels of Th1 cytokine IFN-γ and Th2 cytokine IL4 increased (p < 0.05), whereas IL-2 and IL5 levels remained comparable to the controls in the lipo-rNEp group. At 2 weeks post-second dose, IFN-γ, IL-4, and IL-5 showed a rise compared to controls (p < 0.05). A rise in the IL4 (2 weeks postdose-1) and IL5 (2 weeks postdose-2) was recorded in the al-rNEp group (p < 0.05). Th1 cytokines remained unchanged (Fig. 6B, C). No induction of cytokines or recruitment of immune cells was noted for the rNEp group.
Expression of immune response genes in the spleens of immunized mice
We determined mRNA levels of immune response genes (IRGs) in the (1) spleens of immunized mice and (2) splenocytes, cultured and restimulated with rNEp (Table 2 and Figure 7A, B). Induction of IFN-γ and IL-12 genes in the spleens of lipo-rNEp-immunized mice as well as stimulated splenocytes, while alum induced Th2 cytokine gene IL-10 in the respective stimulated splenocytes.
2w-1, 2 weeks postdose-1; 2w-2, 2 weeks postdose-2.
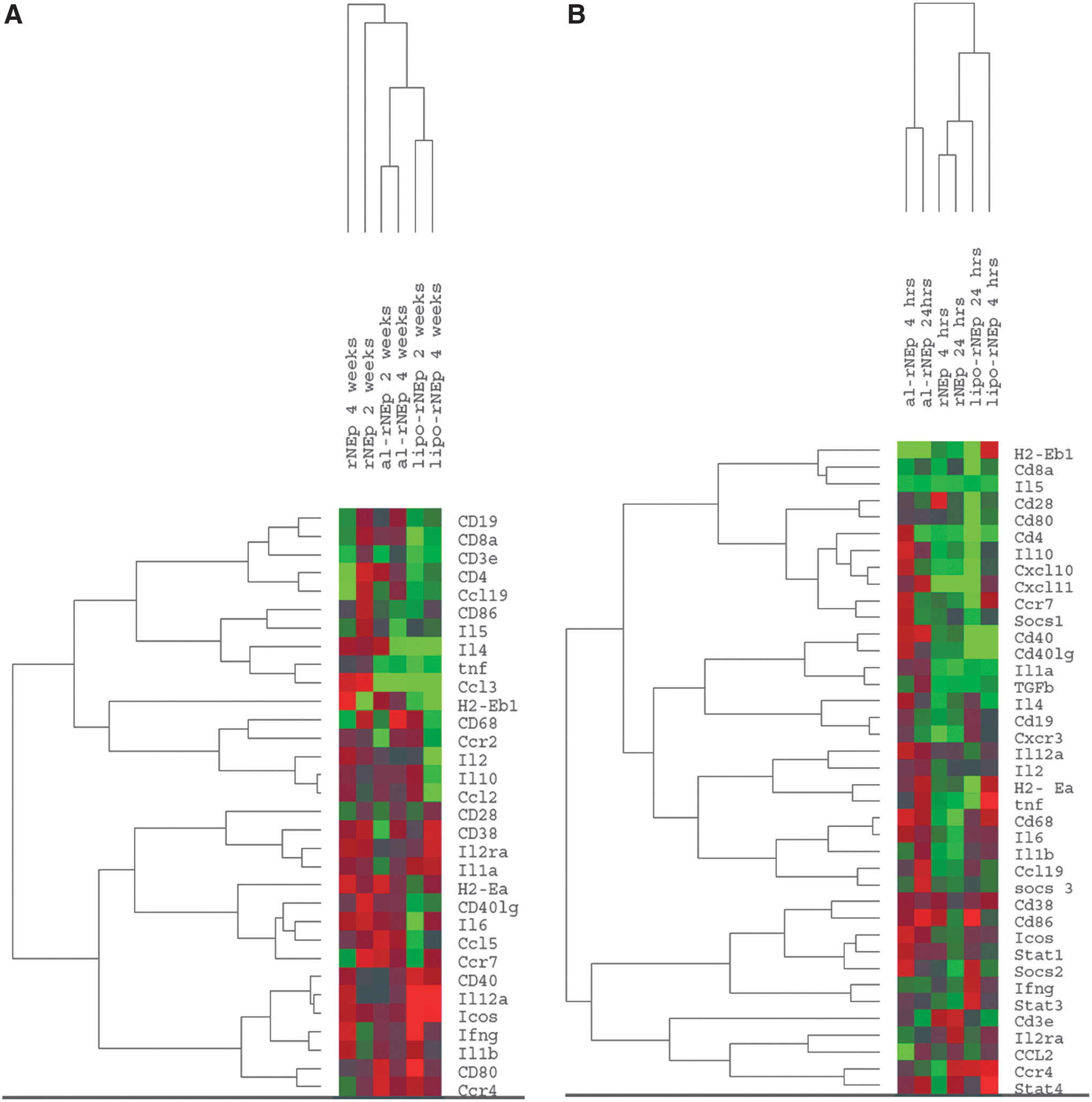
Heatmap of immune response gene expression in splenocytes.
Gene expression pattern of splenocytes (Fig. 7A) and cultured stimulated splenocytes (Fig. 7B) from the immunized mice showed that while in the first category, nonadjuvanted and adjuvanted groups were separated, the alum-adjuvanted group formed a distinct cluster when the antigen stimulation was considered. In accordance with higher gene expression levels, the lipo-rNEp, stimulated for 4 h, formed a distinct branch.
Discussion
For the development of hepatitis E vaccine, c-terminal ORF2 protein fragments of different sizes such as 495aa, 239aa and 150aa have been evaluated (3,12,19,36,37,46). Being the smallest protein, we opted for liposome as the adjuvant in our protein formulations, earlier used in the DNA+corresponding protein format (3,12). Furthermore, a preliminary analysis of probable mechanisms of adjuvant action was also undertaken. It is pertinent to note here that although alum is a conventional adjuvant for human vaccines used over 80 years, the exact mechanism of adjuvant action is being understood only recently (14,18,20,25,30).
rNEp alone was a poor immunogen eliciting antibodies only after the second dose that diminished significantly after 1 month, necessitating a definite need for a potent adjuvant. The immunogenicity of E2 was dramatically improved by the addition of 26 amino acids at the N terminal that led to the formulation of VLPs (27). Alum was the adjuvant used. With rNEp, addition of both adjuvants improved immunogenicity, intramuscular route being superior to the subcutaneous. Liposome had a higher impact on the humoral response than alum as evident by the antigen-sparing capacity and significantly higher antibody titers after each dose (Fig. 2A). Although alum is known to predominantly induce humoral immunity, liposome was a stronger inducer than alum. Despite the limitations of murine model, the possibility of higher initial titers leading to longer antibody persistence cannot be ruled out. Based on mathematical modeling, the only licensed vaccine was predicted to confer persistence of antibodies for 8 or >30 years depending on the model type (10). Compared to E2, HEV239 vaccine (additional 26 amino acids at the N terminal of E2) led to 260-fold higher seroconversion in mice. Both formulations were alum adjuvanted and particulate antigens (27).
The inherent immune stimulatory properties of adjuvants play a significant role in the generation of specific immune response that is antigen driven. When adjuvant alone was used, activation of innate immunity was evident by the secretion of inflammatory cytokine IL-1β (liposome) and proinflammatory cytokine IL-6 (alum) in the muscles at early time points following immunization (Fig. 3A, B). Raised proportion of CD11c+CD86+ DCs in the DLNs and activated CD11c+CD69+ DCs in the spleen at 24 h postdose suggests DC-dependent action of liposome (Fig. 4A, B). Taken together, compared to alum, liposome is a better immune stimulator. These results are in accordance with earlier studies demonstrating better immunostimulatory potential of liposomes compared to alum (13,16,23). B-cell activating property of rNEp, a small protein (23 kDa, 150aa), as reflected by the induction of CD19+MHC-II+ cells in the DLNs post-both doses is noteworthy (Fig. 5A).
Comparison of adjuvanted formulations at early (4, 12, 24 h postdose-1) and later (2 weeks postdose-1 and -2) time points could distinguish the modes of action. Immunization with lipo-rNEp activated cells associated with innate immune response leading to secretion of inflammatory cytokines IL-1β and IL-6 (both probably by macrophages) at the site of injection (Fig. 3A, B), while IL-6 was secreted by the al-rNEp group. Activation of innate immunity is an important requisite for a vaccine formulation to be immunogenic (6). Although we did not detect recruitment of antigen presenting cells (APCs) in the muscles, increased levels of CD11c+CD86+ DCs in the DLNs and activated DCs (CD11c+CD69+) along with the total DC population in the spleens from the immunized mice at 4 h postdose reveal that DCs represent the predominant APCs transporting rNEp to the DLNs and spleen (Fig. 4A, B). In this study, the addition of adjuvant was beneficial as judged by earlier detection of DCs with early activation marker CD69 (4 h) compared to liposome alone (24 h). In contrast, except for the rise in IL-6 levels at the site of injection, alum-adjuvanted formulation did not lead to activation of the immune cells investigated. These early events clearly demonstrate that liposome is a better immune stimulator than alum when adjuvanted with rNEp. Our results confirm earlier observations of the absence of activation of B, T, DCs or NK cells in DLNs when alum was adjuvanted with several antigens (9,28).
At later time points, that is, 2 weeks post-first or post-second dose, a further difference emerged. For antigen presentation and subsequent T-cell activation, maturation of APCs is obligatory. MHC molecules are important surface markers indicative of the maturation levels of APCs and potential pathways of subsequent antigen presentation. The immunogen itself (rNEp) could recruit CD19+MHC-II+ cells to DLNs after both the doses, while no difference was seen in the spleen (Fig. 5A). Both adjuvanted groups retained the immunogen-induced recruitment of B cells as APCs. In addition, the lipo-rNEp formulation could recruit CD11c+CD86+ DCs to the DLNs and CD11c+MHC-II+ DCs in the spleen (Figs. 5B and 6A). The al-rNEp group was characterized by raised proportion of macrophages carrying MHC-II molecule in the spleen (second dose). Thus, the two adjuvanted formulations recruit B cells as the APCs in the DLNs, the property of the immunogen, while differ in recruiting distinctly different cell types in the spleen; liposome functions through DCs and alum through macrophages. Previously, it was shown that when ovalbumin adjuvanted with DDA liposomes was used for mice immunization, an enhanced antigen uptake by splenic DCs, macrophages, and B cells but not by T cells was recorded (21). Our observation of lack of increased CD3+CD4+CD69+, CD3+CD8+CD69+ cells representing T-cell activation indicates similar activity profile.
A comparison of cytokine responses in the DLNs and spleens of immunized mice revealed adjuvant-driven differences. In the DLNs, an initial Th1 response (elevated IFN-γ levels, postdose-1) and subsequent balanced response (both IFN-γ and IL4 elevated) were induced by lipo-rNEp, while no significant variation was recorded in the al-rNEp group. In the spleen, a more pronounced Th1/Th2 balanced response was seen in the lipo-rNEp group and a distinct Th2 bias was evident in the alum category (Figs. 5C, 6B, and C). Gene expression studies (2 weeks post-second dose) of increased expression of IRGs in the splenocytes of mice from lipo-rNEp group and normally expressed cytokine genes in the alum category confirm higher adjuvanticity of liposome than alum (Fig. 7A, B). At this time point, both IgG subtype analysis (IgG1/IgG2a) and cytokine levels showed a clear Th2 bias in the alum-adjuvanted group (Figs. 2B and 6C). Alum is known to be a Th2 inducer and can directly modulate B cells to differentiate mainly into IgG1 B cells (18).
In the absence of similar studies with HEV239 and HEV495 vaccines, a significant HEV-specific Th1 response (raised IFN-γ levels, Enzyme-Linked Immuno Spot) in mice immunized with a combination HEV (same as HEV239) and HAV (VP1-198aa) vaccine is noteworthy (15). The recombinant viral proteins fused to tuftsin, a naturally occurring, immunostimulatory tetra-peptide derived from the Fc portion domain of the heavy chain of IgG, induced a significantly higher IFN-γ response than when used alone. The role of antibodies in protection against HEV challenge was confirmed by passive transfer experiments in monkeys, whereas the contribution of Th1 response is not yet clear and needs to be evaluated especially after the vaccine-induced anti-HEV antibodies disappear (27).
The question is what is the basis of differential adjuvant effects of liposomes and alum when used with the same amount of rNEp? It is proposed that protection of the entrapped immunogen from extracellular degradation as well as antibody-dependent neutralization plays an important role in determining the adjuvant effect of liposomes, while alum is known for its depot effect. Liposome formulation used in the present study led to early seroconversion, antigen sparing, and higher antibody titers. With our liposomes, similar results were reported earlier with a small hypervariable region of HCV and influenza virus proteins (16). The use of liposomes similar to ours led to an enhanced immune response in mice immunized with HA protein of influenza virus (5). However, immunization with HCV-CpG-NS3 encapsulated in cationic liposomes led to delayed antibody response suggesting alternate mechanisms, probably slow release of the antigen (17). It may be surmised that the type of liposome/antigen seems to affect the rapidity of immune response and needs further investigations.
The type and proportion of APCs recruited by different adjuvants could be another important factor. Recruitment of DCs, the professional APCs well known in enhancing T-cell responses by lipo-rNEp, could possibly be responsible for the enhanced immune response compared to alum. Poly-lactic acid (PLA) adjuvant is shown to function by enhancing CD86 and MHC-II levels on CD11c cells (41). The lower response with alum observed by us could be due to the lower alum concentration compared to other reports (8,22). Comparison of the three most widely used alum formulations, Alhydrogel, Imject alum, and conventional precipitated alum with the antigen NPCGG (4-hydroxy-3-nitrophenyl acetyl ester, a hapten bound to chicken γ globulin), revealed a variable humoral response (7).
Conclusion
In conclusion, our study shows the superiority of liposomes in terms of early seroconversion, antigen sparing, and higher antibody titers when administered with rNEp, than the alum-adjuvanted formulation. These preliminary data also suggest that liposomes work through DCs, while alum functions through macrophages for rNEp. Functional assays for DCs in the context of lipo-rNEp in future are warranted. Although the liposome formulation has produced sterilizing immunity in rhesus monkeys, a comparison with alum was not made in this animal model for hepatitis E.
Footnotes
Acknowledgments
We thank the Director, National Institute of Virology, Pune, for all the facilities. S.S.J. acknowledges the Indian Council of Medical Research (ICMR) for the senior research fellowship and project funding.
Author Disclosure Statement
No competing financial interests exist.