
Abstract
Pro- and anti-inflammatory markers (tumor necrosis factor [TNF]-α, TNF-β, interferon [IFN]-γ, interleukin [IL]-6, IL-8, IL-10, and C-reactive protein [CRP]) were investigated in 80 patients infected with dengue viruses, 100 patients presenting with febrile illness but negative for dengue, and 99 healthy subjects. Immunoenzyme methods were used for quantitative assays in the plasma. Polymorphisms of TNF-α, TNF-β, IL-6, IL-8, and IL-10 genes were assessed by polymerase chain reaction (PCR)-restriction fragment length polymorphism and allele-specific oligonucleotide (ASO)-PCR for the IFN-γ. The highest mean serum levels of TNF-α, IFN-γ, IL-8, and CRP were observed in dengue-positive individuals. TNF-β, IL-6, and IL-10 levels were significantly higher in the dengue-negative individuals. No cytokine expression pattern was evidenced according to virus serotype. Genotypic frequency distributions were statistically significant for the polymorphisms of TNF-α and IFN-γ among positive, negative, and control dengue groups and IFN-γ among groups DENV-1, DENV-2, DENV-3, and controls. Modulation of cytokine expression and polymorphisms is a complex matter and needs further explanation considering the ethnic origins of the Brazilian population.
Introduction
D
Among the several mechanisms that have been described for DHF pathogenesis, the most studied corresponds to the shift of the cytokine-secreting profile of T helper (Th) lymphocytes (16,17). The pathogenesis of the disease caused by dengue virus is not yet fully elucidated, and some associations between increased cytokine levels and different clinical manifestations have been reported (12,16,32). Plasma levels of cytokines, such as tumor necrosis factor (TNF)-α, interferon (IFN)-γ, interleukin (IL)-2, IL-6, IL-8, and IL-10, and C-reactive protein (CRP), are also significantly increased in DHF patients compared to dengue fever (DF) patients (5,12,39,49,67).
Polymorphisms in the promoter regions of several cytokine genes can affect gene transcription and, consequently, cytokine production, which might be associated with disease outcome (23,68). Several studies have shown that cytokine gene polymorphisms can influence the evolution of several pathologies, such as autoimmune diseases, neoplasias, and post-transplant complications, and they are thus considered important markers of susceptibility to several diseases (10,20,23,35,70,74,75).
The present study investigated the frequency of polymorphisms and the expression of pro- and anti-inflammatory response markers (TNF-α, TNF-β, IFN-γ, IL-6, IL-8, IL-10, and CRP) and their possible association with dengue viral infections.
Materials and Methods
Characterization of the groups
During the dengue epidemic period, which ranged from October 2006 to April 2007, 180 persons presenting with a febrile illness were randomly selected, according to their free will in accepting to take part in the study, as they sought medical attention and laboratory diagnosis at the outpatient unit of the Laboratory of Arbovirology and Haemorrhagic Fevers of the Evandro Chagas Institute, Belem, Para.
Two groups were initially formed. The first group included 80 patients infected with DENV-1 (n = 19), DENV-2 (n = 8), and DENV-3 (n = 53). There were 34 women and 46 men, with a mean age of 33 years. The second group included 100 patients with a febrile disease without dengue infection and was formed by 47 women and 53 men, with a mean age of 31 years. A third group was formed as a control and included 99 samples from healthy volunteers of the serum bank of Virus Laboratory of the Federal University of Para and included 23 women and 76 men, with a mean age of 26 years.
All subjects included in the infected and noninfected groups answered an epidemiological questionnaire and had a blood sample drawn. Specifically, peripheral blood was collected into 4-mL tubes containing EDTA as anticoagulant and then transported to the Virus Laboratory. Samples were centrifuged, and plasma and whole blood were aliquoted separately and stored at −20°C until used.
Dengue viral infection diagnosis was performed at the Laboratory of Arbovirology and Haemorrhagic Fevers of the Evandro Chagas Institute by means of virus isolation using cell culture of Aedes albopictus clone C6/36 cells (73). Intracellular infection of DENV was confirmed by an indirect immunofluorescence test (IFI) using ascitic fluid of mice with purified antidengue monoclonal antibodies (DENV-1, DENV-2, DENV-3, and DENV-4) produced by Technology Laboratory of Monoclonal Antibodies (LATAM/BIO-MANGUINHOS/FIOCRUZ) (28).
The present project was approved by the Research Ethics Committee of the Pará Hemotherapy and Hematology Center Foundation (HEMOPA). All individuals were previously informed about the project, and those who agreed to enroll signed an informed consent form.
Quantitative determination of cytokine and CRP levels in peripheral blood
Quantitative determinations of TNF-α, TNF-β, IFN-γ, IL-6, IL-8, and IL-10 levels in the plasma were performed by means of an immunoenzyme assay (Human ELISA Ready-SET-Go; EBioscience, Inc., San Diego, CA). Cytokine levels were calculated based on standard curve values using the software GraphPad InStat 3.06 (San Diego, CA), and the results were expressed as picograms per milliliter. The CRP levels were also assessed by means of an immunoenzyme assay (Microwell ELISA C-reactive protein; Diagnostic Automation, Inc., Calabasas, CA). The detection limits ranged between 0.1 and 10 mg/L. Samples with CRP levels above 10 mg/L were diluted at 1:1,000 and retested. All tests were performed according to the manufacturers' instructions.
Cytokine gene polymorphism analysis
Genomic DNA was extracted from peripheral blood leukocytes using a modified phenol–chloroform method (69).
Polymorphisms in the TNF-α (−308G/A; rs1800629), TNF-β (+252G/A; rs909253), IL-6 (−634C/G; rs1800796), IL-8 (−353A/T), and IL-10 (−1082 G/A; rs1800469) genes were assessed by restriction enzyme analysis using the restriction fragment length polymorphism method on polymerase chain reaction (PCR)-amplified fragments, as described in other studies (2,22,48,52,57). The IFN-γ (+874 T/A; rs243056) polymorphism was investigated by allele-specific oligonucleotide PCR (ASO-PCR) (61).
The amplification and digestion products were subjected to agarose gel (1% to 4%, according to product size) electrophoresis (100 V/45 min) in 1 × TAE buffer (40 × TAE stock—1.6 M Trisbase, 0.8 M sodium acetate, and 40 mM EDTA-Na2/1,000 mL deionized water), containing 5 μL SYBR® Safe, and visualized using an ultraviolet (UV) transilluminator.
Statistical analyses
Kruskal–Wallis one-way analysis of variance (ANOVA) was used to compare more than two samples of serum cytokine levels. The odds ratio was calculated for the analysis of cytokine polymorphisms. Cytokine allele and genotype frequencies were obtained by direct gene count and are expressed as percentages. The null hypothesis was rejected at a significance level of 0.05 (5%) (p ≤ 0.05). Statistical analyses were performed using the software BioEstat 5.0 (4).
Results
The highest mean serum levels of TNF-α, IFN-γ, IL-8, and CRP were observed in dengue-positive individuals compared with the noninfected and control groups. Significant associations in the comparison of serum levels among the three groups were found for IFN-γ (p < 0.01) and CRP (p < 0.01). Associations were also found in the comparison of IL-8 serum levels between those of dengue-positive and control groups and dengue-negative and control groups (p < 0.05). There were no significant differences in TNF-α levels among the groups (p = 0.0558) (Fig. 1). The mean serum levels of TNF-β, IL-6, and IL-10 were significantly higher in the dengue-negative group compared to the dengue-positive and control groups.
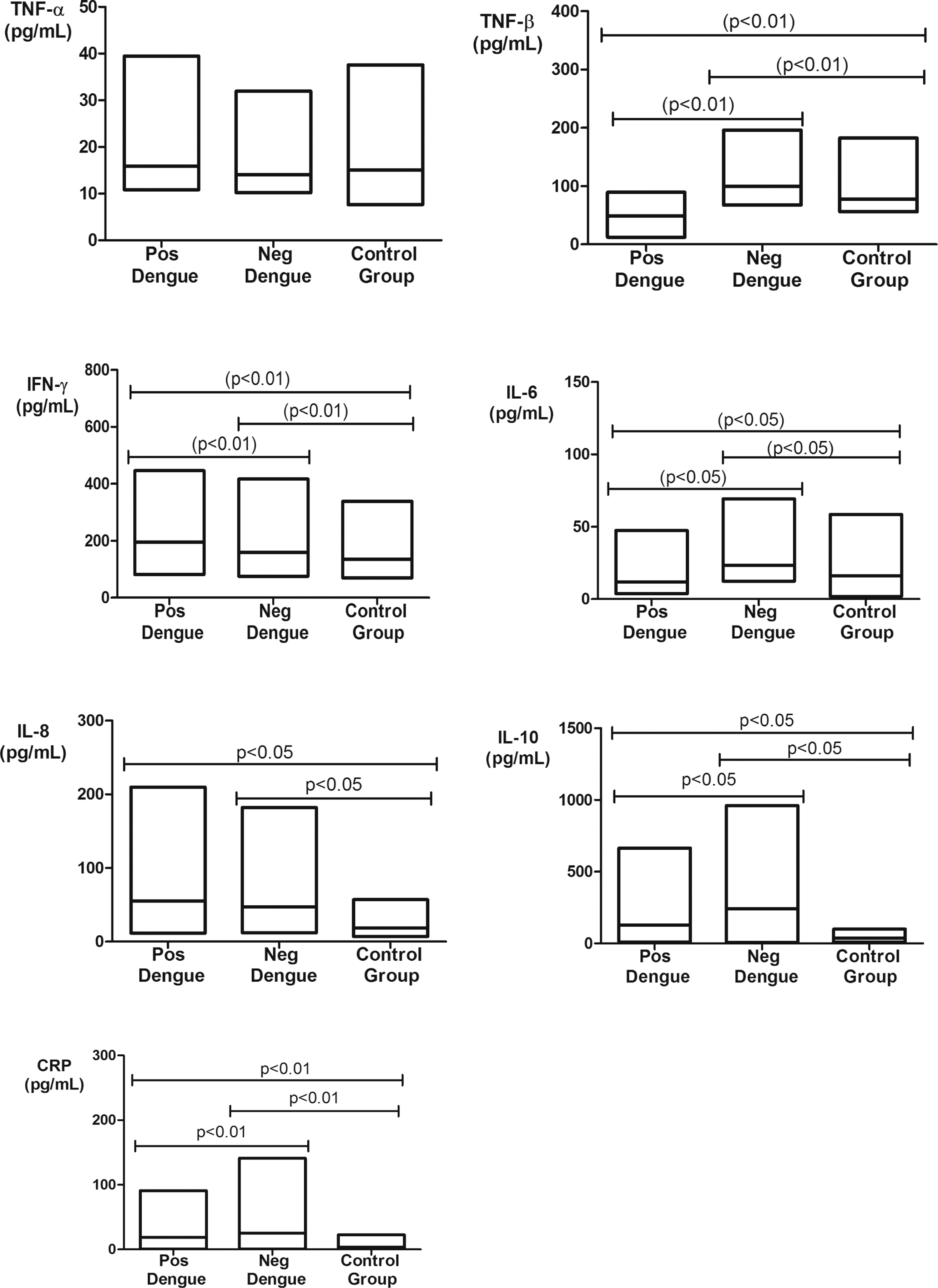
Serum TNF-α, TNF-β, IFN-γ, IL-6, IL-8, IL-10, and CRP levels in dengue-positive, dengue-negative, and control groups. TNF-α: dengue positive (14.6 ± 3.1 pg/mL), dengue negative (13.3 ± 2.1 pg/mL), and control group (14.3 ± 4.9 pg/mL); TNF-β: dengue positive (49.3 ± 14.9 pg/mL), dengue negative (96 ± 22.3 pg/mL), and control group (73.8 ± 14.8 pg/mL); IFN-γ: dengue positive (192.1 ± 86.2 pg/mL), dengue negative (152.1 ± 59.5 pg/mL), and control group (123.6 ± 39.3 pg/mL); IL-6: dengue positive (10.4 ± 6.0 pg/mL), dengue negative (20.3 ± 5.7 pg/mL), and control group (15.5 ± 10.3 pg/mL); IL-8: dengue positive (50.8 ± 31.2 pg/mL), dengue negative (46.1 ± 34.4 pg/mL), and control group (16.8 ± 7.4 pg/mL); IL-10: dengue positive (86.4 ± 62.4 pg/mL), dengue negative (184.4 ± 132.7 pg/mL), and control group (37.1 ± 14.5 pg/mL); and CRP: dengue positive (14.5 ± 5.4 mg/L), dengue negative (8.6 ± 6.9 mg/L), and control group (2.8 ± 2.2 mg/L).
The genotype and allele frequencies of polymorphisms are listed in Table 1. The distribution of genotype frequencies of the −308 G/A polymorphism in TNF-α showed the GG genotype at the highest frequency in the dengue-negative group compared to controls (p = 0.0395), which is opposite to what was observed with respect to the heterozygous genotype (p = 0.0052). There was no association in the comparison of allele frequencies.
N, number of analyzed individuals; OR, odds ratio; p1, p-value of dengue-positive versus dengue-negative groups; p2, p-value of dengue-positive versus control groups; p3, p-value of dengue-negative versus control groups.
The +874 T/A polymorphism in IFN-γ exhibited a significantly higher frequency for the AA genotype in the comparison between the dengue-positive group and the other groups (p = 0.0255 and p = 0.0165) and for the TT genotype when comparing between the dengue-positive and control groups (p = 0.0145). The frequency of the heterozygous genotype (AT) was lower in the dengue-positive group, with a significant increase in the dengue-negative and control groups (p = 0.0088, p < 0.0001, and p = 0.0304). There was no association in the comparison of allele frequencies.
There were no significant associations of the genotype and allele frequencies of polymorphisms in TNF-β (+252 G/A), IL-6 (−634 C/G), IL-8 (−353 A/T), and IL-10 (−1082 G/A) in any of the studied groups.
The mean level distributions of TNF-α, TNF-β, IFN-γ, IL-6, IL-8, IL-10, and CRP according to viral serotypes are displayed in Figure 2. The mean serum TNF-α levels were highest in the DENV-3 group, with a significant difference compared to the DENV-1 (p < 0.05).
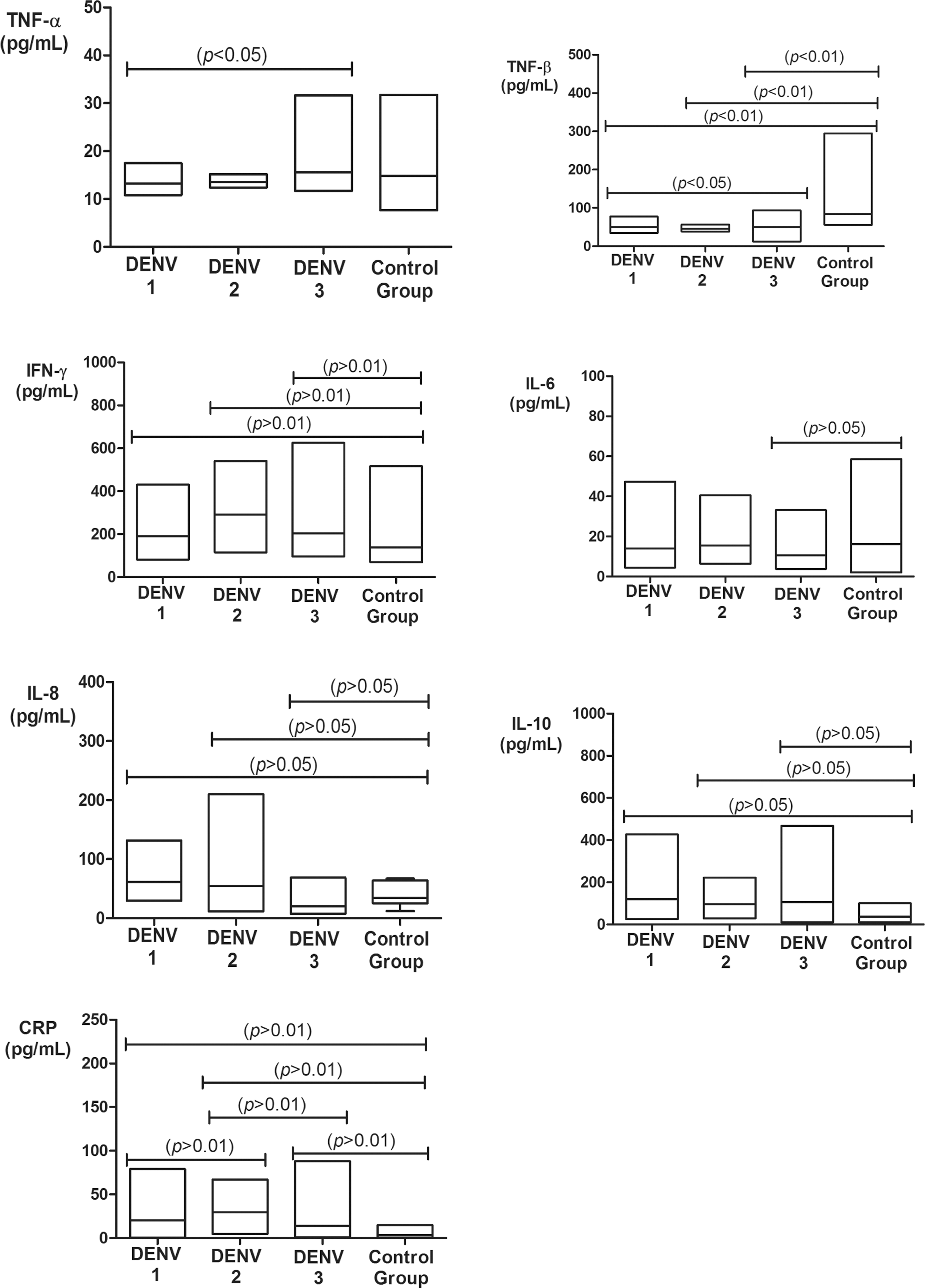
Serum TNF-α, TNF-β, IFN-γ, IL-6, IL-8, IL-10, and CRP levels in the dengue-positive group according to serotype and control group. TNF-α: DENV-1 (13.2 ± 2.1 pg/mL), DENV-2 (13.6 ± 1.2 pg/mL), DENV-3 (15.2 ± 3.5 pg/mL), and control group (14.3 ± 4.9 pg/mL); TNF-β: DENV-1 (53 ± 12.5 pg/mL), DENV-2 (50.6 ± 14.5 pg/mL), DENV-3 (43.7 ± 5.1 pg/mL), and control group (73.8 ± 14.8 pg/mL); IFN-γ: DENV-1 (175.5 ± 84.2 pg/mL), DENV-2 (181.6 ± 38.6 pg/mL), DENV-3 (179.3 ± 64.1 pg/mL), and control group (123.6 ± 39.3 pg/mL); IL-6: DENV-1 (11.9 ± 7.0 pg/mL), DENV-2 (15.6 ± 12.7 pg/mL), DENV-3 (9.6 ± 5.4 pg/mL), and control group (15.5 ± 10.3 pg/mL); IL-8: DENV-1 (61.3 ± 33.5 pg/mL), DENV-2 (41.9 ± 21.3 pg/mL), DENV-3 (47.9 ± 31 pg/mL), and control group (16.8 ± 7.4 pg/mL); IL-10: DENV-1 (101.5 ± 71.2 pg/mL), DENV-2 (96.2 ± 70.2 pg/mL), DENV-3 (99 ± 57.7 pg/mL), and control group (37.1 ± 14.5 pg/mL); and CRP: DENV-1 (14.1 ± 5.6 mg/L), DENV-2 (23.8 ± 5.3 mg/L), DENV-3 (7.1 ± 3.3 mg/L), and control group (2.8 ± 2.2 mg/L).
As for TNF-β, the highest mean serum levels were observed in the control group, with significant differences between DENV-1 and DENV-3 groups (p < 0.05) and between DENV-1, DENV-2, and DENV-3 compared to the control group (p < 0.01).
IFN-γ, IL-6, and CRP showed the highest mean serum levels in the DENV-2 group. A significant difference in serum IFN-γ levels was found between the DENV-1, DENV-2, and DENV-3 groups and that of the control group (p < 0.01). With respect to IL-6, there was a significant difference in the mean serum levels between the DENV-3 and the control group (p < 0.05). For CRP, there were significant differences in the mean serum levels when comparing all groups with each other (p < 0.01).
The mean serum IL-8 and IL-10 levels were highest in the DENV-1 group, and the mean serum levels of both cytokines were significantly different in the comparison of all serotype groups with the control group (p < 0.05). Additionally, no significant correlation was observed between cytokine genotypes and serum levels (data not shown).
The genotype and allele frequencies of polymorphisms according to viral serotype are listed in Table 2. The TT genotype of the IFN-γ +874 polymorphism showed a higher prevalence among patients infected with DENV-2 (37.5%), followed by those infected with DENV-3 (16.9%), with significant associations observed when the genotype frequencies were compared to the control group (p = 0.0079 and p = 0.0332, respectively), although not when the allele frequencies were compared. Regarding heterozygosis (AT), significant differences were found for the frequencies of the three dengue serotypes compared to the controls (p = 0.0341, p = 0.0010, and p = 0.0027, respectively). No significant associations were found when comparing the genotypes or the allele frequencies of the remaining polymorphisms.
ND, not done; p1, p-value of DENV 1 versus DENV 2; p2, p-value of DENV 1 versus DENV 3; p3, p-value of DENV 1 versus control group; p4, p-value of DENV 2 versus DENV 3; p5, p-value of DENV 2 versus control group; p6, p-value of DENV 3 versus control group.
Discussion
In the present study, serum TNF-α, TNF-β, IFN-γ, IL-6, IL-8, IL-10, and CRP levels were quantified and compared among dengue-positive individuals, dengue-negative febrile patients, and a control group of healthy individuals. Among the several hypotheses explaining the pathogenesis of the severe disease caused by dengue virus, cytokines play a highly relevant role (16,17,65).
There are reports of significantly higher levels of TNF-α in DHF patients (5,12,14,49) but not among febrile patients as reported here and elsewhere (11,19,77). TNF-α acts as a part of the extrinsic pathway for apoptosis induction, and it also interacts with receptors on endothelial cells, inducing increased vascular permeability, which allows for leukocytes to extravasate into the site of infection. Increased TNF-α levels may be associated with the hemorrhagic phenomena observed in dengue. Given that the patients examined in the present study exhibited no signs of hemorrhage, it is possible that the results observed are compatible with lower TNF-α expression similar to the values found in the control group.
IFN-γ plasma levels were significantly higher in dengue-positive patients compared to the dengue-negative and control groups, which is in agreement with the results from other studies (5,14). It has been shown to be much higher in DF compared to DHF cases (11,14,62). In Taiwan, Chen et al. (19) reported lower IFN-γ levels in patients who died from DHF compared to survivors and surviving DHF patients who exhibited higher levels than DF patients and controls. The immunoregulatory function of IFN-γ during cell-mediated cytotoxicity probably leads in its increased production to a protective role in patients with acute, mild, and moderate disease.
IL-8 induces endothelial cell changes by increasing cell permeability and plasma extravasation, which are relevant in DHF and dengue shock syndrome (DSS) (32,71). A strong correlation has been demonstrated between increased IL-8 levels and DHF pathogenesis (7,9,16,62), as well as a direct relationship between disease severity and fatality and IL-8 levels (64). Although only moderate levels of IL-8 were present in the dengue-positive group, it was not significantly different from the dengue-negative group. Increased IL-8 levels were observed in both dengue patients and a group of patients with a bacterial infection compared to the control group (40). The same has been observed with other viral infections, such as those caused by influenza virus, respiratory syncytial virus, rhinovirus, paramyxovirus, and hepatitis C virus (3,8,27,53,72), which implies in a nonspecific immunological reaction.
Although acute phase protein synthesis, including that of CRP, may be stimulated by IL-6 (30,54), in the present study, there was no relationship between CRP and IL-6, given that the highest serum CRP levels were detected in the dengue-positive group, whereas the highest IL-6 levels were detected in the dengue-negative febrile group. CRP levels are significantly increased in DHF patients, a finding that was further associated with the occurrence of secondary dengue virus infection. Finding higher levels among dengue-positive patients reinforces its role as a marker of the inflammatory response to the virus, even among patients with less severe acute forms of the disease.
TNF-β, IL-6, and IL-10 were higher in the dengue-negative febrile group, and TNF-β and IL-6 levels were lower in dengue patients compared to controls. Gagnon et al. (25) obtained similar results with TNF-β in peripheral blood mononuclear cells obtained from children infected with dengue, and there was no significant association among DHF patients, DF patients, and those with febrile illnesses other than dengue. IL-6 findings contradict reports of increased IL-6 levels in dengue patients (24,49,60,67). Chaturvedi et al. (16,17) reported its gradual increase, reaching maximum levels on the second day of the disease, and a rapid decrease during the course of the disease. It is possible that this was not observed in the present study, as most of the samples were collected between the fourth and fifth days, for the purposes of diagnosis. High levels of IL-6 were reported in severe cases of DHF and DSS, which may contribute to modulate coagulation and plasma extravasation (11,32,33,39,62,63), as well as among dengue-negative febrile patients with dengue-like symptoms, which may be related to the presence of this cytokine (26,31,38,66). Our findings may be explained by the proinflammatory action produced by activated macrophages and T lymphocytes, which plays important roles in fever induction (at the level of the hypothalamus) and acute and transient inflammatory responses.
Increased IL-10 levels (an anti-inflammatory cytokine, which is characteristically known to modulate the Th1 response) were observed in dengue patients, particularly among DENV-1 serotypes. Its increase is a possible result of either a tissue damage controlling Th2 response or the action of T-bet+Foxp3-Th1 cells, as has been suggested (36); IFN-γ-producing Th1 cells can simultaneously synthesize IL-10 as a form of autoregulation during a response to intracellular infectious agents. Other studies also suggest the association of IL-10 with the pathogenesis of several diseases other than dengue (18,43,53,56). Increased IL-10 levels seem to contribute to the development of DHF and have been associated with reduced numbers of platelets (5,51), transient T-cell anergy (5), and reduced T-cell apoptosis (55). Maximum IL-10 levels were detected in grade IV DHF patients, whereas milder dengue cases exhibited lower or absent IL-10 levels (16). Additionally, the progression to more severe cases was accompanied by a shift of the immune response from Th1 to Th2.
A cytokine expression pattern, according to dengue virus serotype, could not be evidenced. IL-8 and IL-10 were exacerbated in DENV-1 infections; DENV-2 infections exhibited higher levels of IFN-γ, IL-6, and CRP, and DENV-3 infections stimulated higher production of TNF-α and TNF-β. The lack of association between the three serotypes has also been shown by others with TNF-α (49), IL-10 (51), and IFN-γ and IL-6 (6).
The influence of cytokine gene polymorphisms on several infections and diseases has been frequently reported (2,13,42,58,75,81). The distribution of genotype frequencies of TNF-α at position −308 demonstrated the increased presence of the heterozygous AG genotype in the control group with respect to the dengue-positive and dengue-negative groups, but only the comparison between the dengue-negative and control groups exhibited a significant association. A significant increase in the TNF-308A allele in DHF patients compared to DF patients, but not with the control group, was described, suggesting that the allele is associated with the production of higher levels of TNF-α (23,59).
The genotype and allele frequencies of the TNF-β +252 G/A polymorphism in the dengue-positive, dengue-negative, and control groups exhibited no significant associations when compared. It is relevant to mention that there are no reported studies investigating the TNF-β gene polymorphism at position +252 in dengue virus-infected patients, but this polymorphism has been associated with hepatitis B and hepatitis C virus infections (37) and noninfectious diseases, such as multiple sclerosis, systemic lupus erythematosus, and acute ischemic stroke (1,21,41).
The +874 T/A polymorphism in IFN-γ exhibited a higher frequency of the AT genotype accompanied by a low frequency of the TT genotype in the control group with respect to the dengue-positive and dengue-negative groups. A significant association was found between the genotypes of all three groups; however, this association did not occur in the comparison of allele frequencies. The association was not described by others (23,59).
The gene polymorphism analysis of IL-6 (−634 C/G) and IL-10 (−1082 G/A) showed similar genotype and allele frequencies among the groups of the present study. The same was investigated by others (23,59), the same distribution of polymorphisms in IL-6 at position 174 C/G and IL-10 at position −1082 G/A. The association was detected by Perez et al. (59) with the ACC/ATA (−1082, −819, and −592) haplotype while jointly studying the polymorphisms at positions −1082 (G/A), −819 (T/C), and −592 (A/C) of the IL-10 promoter region.
The −353 A/T polymorphism in IL-8 was detected in a higher distribution in the dengue-positive group (59.5%), although neither the genotype nor the allele frequencies were significant. There are more than 70 IL-8 gene polymorphisms known to be associated with several pathologies (44,46,47,50,78), but as yet, there are no studies associating the −353 A/T polymorphism in IL-8 with dengue, despite the evidence of its involvement in the pathogenesis of infection (7,9,16,40,62). The A allele of the −353 A/T polymorphism in IL-8 has been correlated with higher IL-8 production in comparison with the T allele (34), but no study has been able to show associations when analyzing this polymorphism individually, suggesting that the combined effect of several IL-8 polymorphisms is necessary (45,47,50).
The genotype frequencies were in Hardy–Weinberg equilibrium for TNF-β, IL-6, IL-8, and IL-10 but not for TNF-α and IFN-γ. However, according to Weinberg and Morris (79), several single base polymorphisms can violate the Hardy–Weinberg equilibrium for noncausal reasons, especially in ethnically diverse populations, as was the case in the present study.
The only significant association between the frequencies of polymorphisms and serotypes of dengue virus was shown between the genotype frequencies of IFN-γ +874 T/A and the three serotypes, DENV-1, DENV-2, and DENV-3, and control groups. No significant difference was detected by Perez et al. (59) in the distribution of IFN-γ (+874) and IL-10 (−1082) alleles and genotypes, but they were able to associate the presence of the TNF-α (−308) polymorphism with the risk of DHF development in patients infected with DENV-2. It is believed that cytokines might be involved in the pathogenesis of severe dengue, and thus, the levels of cytokines, such as IFN-γ, TNF-α, IL-6, IL-8, and IL-10, are possibly higher in DHF and DSS patients than in DF patients.
In conclusion, the present study corroborates previous studies showing that cytokine levels, including those of IFN-γ, CRP, IL-8, and IL-10, might be increased in dengue virus-infected patients. However, these cytokines were also elevated in individuals from the dengue-negative group, and there was no specific profile of these cytokines in any given group. In contrast to what has been reported, increased IL-6 levels were not detected among the dengue-positive group. It is also relevant to mention that the present study was performed with all the markers in a single population group, with the same genetic diversity within the country characteristics, while most of the published data consider several distinct population groups, a variable that can account for the differences found when comparing some of the results. The results reveal the complex dynamics of cytokine expression and polymorphisms that modulate their expression as a response to more or less severe infections. It is also important to take into consideration the vast genetic diversity of the Brazilian population, which is due to the high degree of ethnic miscegenation, which may generate specific allele and genotype associations with susceptibility or resistance to given infections, depending on the region of the country.
Footnotes
Acknowledgments
We would like to thank all the individuals who participated in this research. The study was funded by Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq), Ministério da Ciência e Tecnologia (MCT), and Programa de Apoio ao Desenvolvimento Científico e Tecnológico (PADCT).
Author Disclosure Statement
No competing financial interests exist.