
Abstract
Infectious hematopoietic necrosis virus (IHNV) infects salmonid fish, resulting in high mortality and serious economic losses to salmonid aquaculture. Therefore, an effective IHNV vaccine is urgently needed. To select an inactivation agent for the preparation of an effective IHNV vaccine, rainbow trout were immunized with mineral oil emulsions of IHNV vaccines inactivated by formaldehyde, binary ethylenimine (BEI), or β-propiolactone (BPL). The fish were challenged 8 weeks after vaccination, and their IgM antibody response and relative percent survival (RPS) were evaluated. The results show that formaldehyde, BEI, and BPL abolished IHNV HLJ-09 infectivity within 24, 48, and 24 h at final concentrations of 0.2%, 0.02%, and 0.01%, respectively. The mean levels of specific IgM, both in serum and mucus (collected from the skin surface and gills), for the three immunized groups (from high to low) ranked as follows: the BPL group, BEI group, and formaldehyde group. From weeks 5 to 9, the mean log2 serum titers of IgM in the BPL group were significantly higher compared with those of the other groups (p < 0.05) during the 9 weeks of observation after vaccination (immunized at weeks 0 and6). Mucus OD490 values of the BPL group were significantly higher compared with those of the other groups (p < 0.05) when reaching their peak at weeks 5 and 8, but the difference between the formaldehyde and BEI groups was not significant (p > 0.05). The BPL-inactivated whole-virus vaccine had the greatest protective effect on the rainbow trout after challenge by an intraperitoneal injection of live IHNV, with an RPS rate of 91.67%, which was significantly higher compared with the BEI (83.33%) and formaldehyde (79.17%) groups. These results indicate that the BPL-inactivated IHNV oil-adjuvant vaccine was more effective than the formaldehyde- or BEI-inactivated vaccines. The results of this study provide an important foundation for further studies on inactivated IHNV vaccines.
Introduction
I
The ongoing threat IHNV poses to salmon and trout fisheries in conjunction with the increasing social and economic value of the fisheries has resulted in the practical need for a licensed, effective IHNV vaccine. In general, three vaccine types have been investigated for the control of IHNV: inactivated IHNV (4,22), attenuated IHNV (15,26), and recombinant IHNV. The last includes DNA or nucleic acid vaccines (5,14,20), N and G subunit vaccines, synthesized in prokaryotic or eukaryotic organisms (13,29), and the synthetic peptide vaccines (11). Inactivated, attenuated, and DNA vaccines have been proven to be effective in laboratory vaccine trials, with relative percent survival (RPS) values approaching 100% (18,19) in some instances. In contrast, subunit vaccines (N and G antigens synthesized by Escherichia coli) and those based on synthetic peptides have performed poorly or inconsistently in laboratory trials. Although there are many reports of IHNV vaccines, only one type of DNA vaccine, APEX-IHN, has been granted vaccine production licenses in Canada (1).
Subsequent to the initial reports of the significant protection afforded by formaldehyde-, binary ethylenimine (BEI)-, or β-propiolactone (BPL)-inactivated IHNV vaccines (4,22), a further study devoted to developing an inactivated IHNV vaccine that produces effective protection against local strains has been minimal. Formaldehyde is commonly used in the traditional preparation of inactivated vaccines. Billions of doses of vaccines prepared with BPL or BEI have been delivered to food animals, and no incidences of toxicity due to residual BPL or BEI have been reported from any country, and no toxicity is expected (6). Although BPL, BEI, and formaldehyde are alkylating agents, they target different molecules within the virion.
In this study, a whole, killed IHNV vaccine was prepared with formaldehyde, BEI, or BPL, and their effects were compared. We used mineral oil as an adjuvant to prepare three inactivated oil-emulsion vaccines and injected rainbow trout intraperitoneally to analyze the immune effects. From an applied science standpoint, our results may provide an experimental basis for developing inactivated vaccines against IHNV through the screening of inactivation agents.
Materials and Methods
Viruses, cells, and antibodies
IHNV HLJ-09 (Accession Number JX649101) was isolated and purified from an acute disease outbreak in Heilongjiang in 2009 by our laboratory and was used for the preparation of the vaccines and the challenge experiment. The CHSE-214 cells were stored by our laboratory and grown in an L15 medium supplemented with 10% fetal bovine serum (FBS). The anti-IgM antibody was produced by healthy New Zealand rabbits, which were injected with the purified protein expressed by the IgM heavy chain gene in rainbow trout emulsified with Freund's adjuvant. The titers of the anti-IgM antibody were 1:204,800, as determined by indirect ELISA analysis.
Fish
Healthy rainbow trout with no previous history of IHNV and an average weight of 23.81 ± 1.09 g were obtained from a cold-water fish experimental station (Mudanjiang, China). The spleens and kidneys of 10 apparently healthy rainbow trout were collected randomly to confirm that there was no existing IHNV infection according to the method of Yue et al. (32). Fish were acclimated to laboratory conditions for 2 weeks and resided in tanks (120 cm × 50 cm × 60 cm) with aeration at 16°C. The dissolved oxygen was kept above 8 mg·l−1. Fish were fed daily with dry pellet food. Fish tanks were cleaned once a week.
Virus propagation
The CHSE-214 cells were grown in T-75 flasks (Corning) in the Leibovitz medium 15 (Cellgro) supplemented with 10% FBS (Excell Bio) at 20°C. IHNV HLJ-09 was inoculated onto confluent CHSE-214 monolayers at a dose equal to a multiplicity of infection of 0.1, and the presence of cytopathic effects (CPEs) were checked for daily through microscopy (Leica). When CPEs were apparent in ∼80% of the monolayer, the viruses were harvested, freeze-thawed twice, and then centrifuged at 2,000 g for 30 min at 4°C to remove cell debris. The supernatant was collected and stored at −80°C for vaccine preparation and the challenge test. The virus titer was determined according to the method of Reed and Muench (25) and expressed as the tissue culture infective dose (TCID50) mL−1.
Virus inactivation
For formaldehyde inactivation, formaldehyde (Aladdin) was added to live IHNV HLJ-09 at a final concentration of 0.2%, and the mixture was agitated at 25°C. Sodium pyrosulfite (Hailun) at a final concentration of 0.05% was added to terminate the inactivation.
For BEI inactivation, a freshly prepared solution of BEI (Solarbio, China) was added to live IHNV HLJ-09 at a final concentration of 0.01%, 0.02%, or 0.04%. The mixture was agitated at 32°C. Virus inactivation was terminated by the addition of sodium thiosulfate (Hailun) at a final concentration of 2%.
For BPL inactivation, BPL (Sigma) was mixed with live IHNV HLJ-09 at a final concentration of 0.005%, 0.01%, or 0.02% and the mixture was agitated at 6°C. The residual BPL was hydrolyzed at 37°C for 2 h. The virus titer was determined before the treatment and at the end of the incubation.
Confirmation of virus inactivation
The loss of viral infectivity after inactivation was confirmed by inoculating cells and fish. CHSE-214 cells were inoculated with IHNV HLJ-09 following the termination of inactivation. The cells were monitored daily for 7 days for the presence of CPEs. Next, 1 mL of the cell culture supernatant was inoculated onto CHSE-214 monolayers in a 25 cm2 flask and monitored daily for 2 weeks for the development of CPEs. A second blind passage was conducted using the same method. Rainbow trout were injected intraperitoneally with 0.2 mL of inactivated IHNV HLJ-09 and their health condition was monitored daily. Live IHNV HLJ-09 was injected as a positive control.
Emulsification of the vaccine
The adjuvant used in this study was mineral oil (Sigma), and water/oil emulsions were made using a homogenizer (IKN) with a standard emulsification stator/rotor connected to an emulsion screen. The oil-based antigen preparation was formulated as water-in-oil, where the water phase with IHNV HLJ-09 was inactivated with 0.2% formaldehyde (agitated at 25°C for 24 h), 0.02% BEI (agitated at 32°C for 48 h), or 0.01% BPL (agitated at 6°C for 24 h) was dispersed into an oil phase (continuous phase containing emulsifiers and stabilizers). The ratio of the water phase to oil phase was 2:3. The three types of vaccines had equal concentrations of inactivated IHNV, equivalent to an IHNV titer of 4 × 106.82 TCID50/mL.
Immunization and sample collection
To evaluate the efficacy of the three inactivated vaccines, healthy rainbow trout were randomly divided into three immunized groups and one control group (100 fish per group). Before vaccination, the experimental fish were anesthetized by immersion in eugenol (1: 5,000 dilution; Shybio). The fish in the three immunized groups were injected intraperitoneally with 0.2 mL of formaldehyde-, BEI-, or BPL-inactivated vaccine (each vaccine contained 8 × 105.82 TCID50 of inactivated IHNV), while the control group received the same volume of saline. The enhanced vaccination was performed after 6 weeks using the same method and volume used for the first vaccination.
The experimental fish were held in separate tanks, with the water temperature maintained at 16°C. The fish were fed with artificial feed once daily. The rainbow trout were anesthetized for 5 min, as indicated above, before the collection of blood and skin samples. The blood samples were collected from three fish in each group by tail venipuncture and were allowed to clot at 4°C for 10 h and then centrifuged at 1,000 g for 10 min. We gently scraped the surface of the trout's skin and its gill with a clean glass to obtain mucus samples and centrifuged the mucus at 1,000 g for 30 min at 4°C with an equal volume of saline before the supernatant was collected. The serum and mucus harvested from each sample were stored at −20°C. The blood and skin samples were collected from 1 to 9 weeks postimmunization.
Determination of the specific IgM in serum and mucus
The levels of the specific IgM in the serum and mucus of the rainbow trout were detected using an indirect ELISA on a 96-well reaction plate that was coated with 200 μL of 107.5 TCID50/mL of IHNV HLJ-09 for 18 h at 4°C, blocked with 100 μL of 5% skim milk for 2 h at 37°C, and incubated in 100 μL of serial twofold dilutions of the test serum or undiluted mucus samples for 1 h at 37°C, followed by incubation with 100 μL of rabbit anti-IgM in juvenile rainbow trout hyperimmune serum (1:1,000) for 1 h at 37°C. Next, 100 μL of HRP-labeled goat anti-rabbit IgG (1:2,000 dilution; TaKaRa) was added for 1 h at 37°C, followed by an incubation in OPD-H2O2 for 15 min at 37°C against an opaque background. After adding the termination liquid, the light absorbance values (at a wavelength of 490 nm) of the samples were determined using a Microelisa Stripplate Reader (Bio Tek).
Challenge test
Duplicate groups of 50 fish were challenged at 8 weeks postvaccination with an inoculation of 10 × LD50 of live IHNV [LD50 = 10−3.36, 0.1 mL per fish, intraperitoneal injection; dose was determined from the preexperiment using the method of Reed and Muench (25)]. The fish were monitored daily, and dead fish were removed from the tank until mortality ceased at day 21. A portion of the dead fish was analyzed for the presence of IHNV. The spleens and kidneys of the dead fish were collected to confirm the pathogen using a PCR assay (32). The RPS was calculated using the following formula: RPS = (1 − mortality in vaccinated fish/mortality in control fish) × 100%.
Statistical analysis
Statistical analysis of the data was performed using JMP version 7.0. The results are presented as the mean ± standard error. The statistical significance was calculated by one-way analysis of variance (ANOVA) and defined at the level of p < 0.05.
Results
Virus inactivation
BEI at final concentrations of 0.01%, 0.02%, and 0.04% abolished IHNV HLJ-09 infectivity and reduced TCID50 value to zero within 96, 48 and 48 h, respectively (Table 1). BPL at final concentrations of 0.0025%, 0.005%, and 0.01% reduced the TCID50 value of IHNV HLJ-09 to zero within 72, 48, and 24 h, respectively (Table 2). With the exception of the mixture with a final concentration of 0.0025% BPL, the other virus mixtures, whose TCID50 values dropped to zero, abolished viral infectivity in the CHSE-214 cells, which was confirmed by inoculation followed by two blind passages on the CHSE-214 cells without apparent CPEs.
BEI, binary ethylenimine; IHNV, infectious hematopoietic necrosis virus.
BPL, β-propiolactone.
Rainbow trout injected intraperitoneally with the inactivated IHNV HLJ-09 preparations were healthy, whereas rainbow trout injected with the live IHNV HLJ-09 began to die at day 4, and the cumulative mortality rate reached 96.7% by day 21. The dead rainbow trout had the same symptoms as rainbow trout naturally infected with IHNV.
Treating IHNV HLJ-09 with formaldehyde at a final concentration of 0.2%, followed by agitation at 25°C for 24 h, abolished the viral infectivity in the CHSE-214 cells and the rainbow trout.
Antibody responses
In the three immunized groups, the titers of the specific IgM in serum did not increase significantly until week 3 postimmunization compared with those of the control group. From weeks 3 to 9, the levels of IgM observed in the serum (from high to low) in each group ranked as follows: the BPL group, the BEI group, and the formaldehyde group. The mean IgM titers reached a peak at week 5 (formaldehyde group: 7.67 log2, BEI group: 7.67 log2, and BPL group: 10 log2), and the difference among the three immunized groups was significant (p < 0.05) at this time point. The mean titers reached a second peak at week 8 (due to the enhanced immunity administered at week 6); at this time point, the differences in the mean titers among the BPL group (11.33 log2), the BEI group (9.67 log2), and the formaldehyde group (8.67 log2) were also significant (p < 0.05) (Fig. 1a).
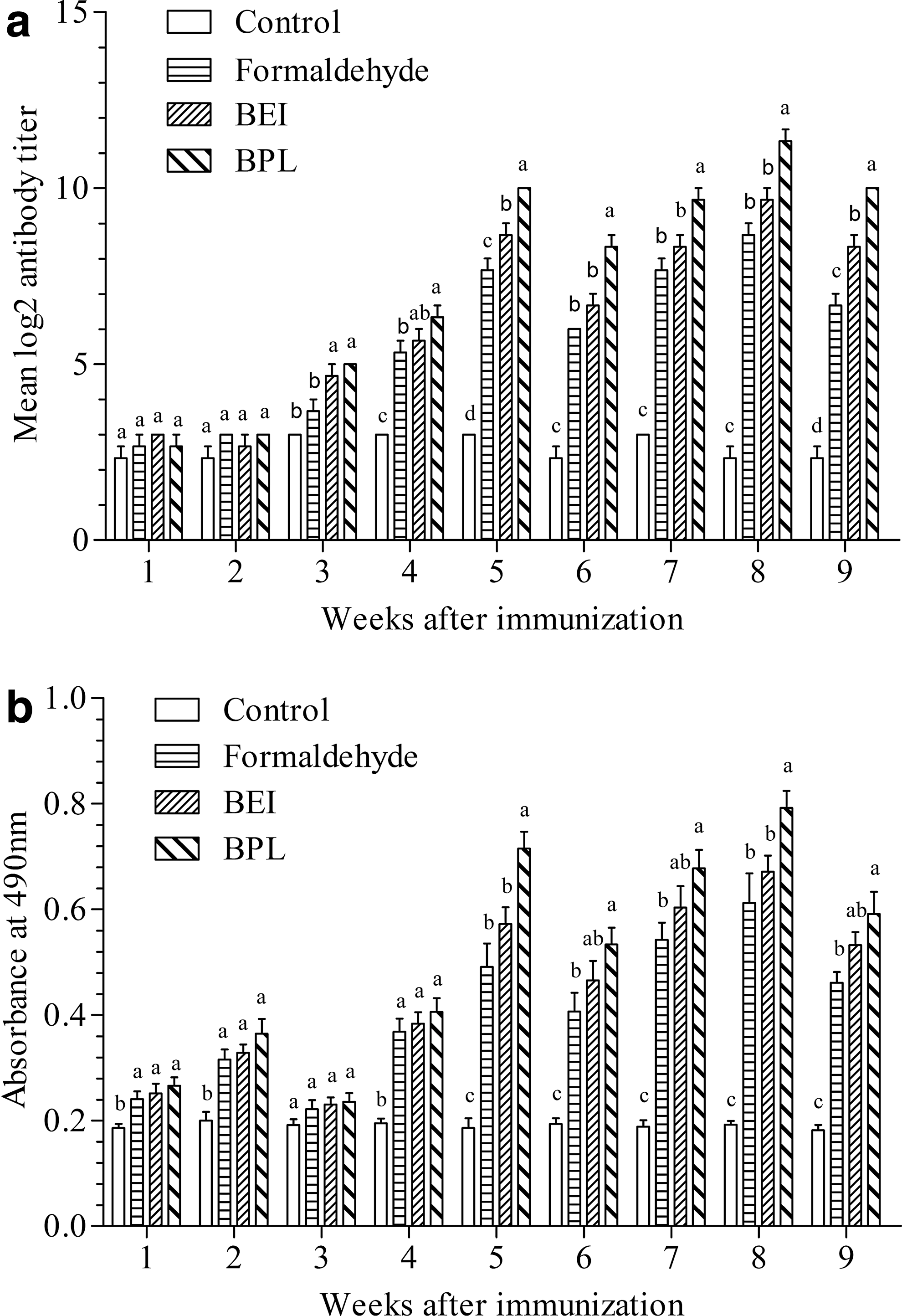
Specific IgM antibody titers in immunized rainbow trout sera
For the mucus antibody levels, there were significant increases in the OD490 values at weeks 1, 2, and 4 postimmunization for the three immunized groups compared with the levels in the control group. Subsequently, the values increased rapidly, reaching their peak at weeks 5 and 8. Throughout the experiment, the OD490 values for the three immunized groups observed in the mucus (from high to low) ranked as follows: the BPL group, the BEI group, and the formaldehyde group. At weeks 5 and 8 postimmunization, the IgM levels of the BPL group were significantly higher compared with the levels of the other two immunized groups (p < 0.05), while the difference in IgM levels between the formaldehyde and BEI groups was not significant (p > 0.05) (Fig. 1b).
Challenge test
Mortality in the control group occurred between 4 and 14 days postinjection, with a cumulative mortality rate of 96%. The fish exhibited decreased appetite and severe hemorrhages on the body surface accompanied with exophthalmia and swelling. The gills were pale and anemic, and the kidney and spleen were also anemic. The mortality that occurred in the immunized groups was slower compared with that in the control group. However, the signs of disease in the dead fish in the immunized groups were similar to those observed in the control group. The spleens and kidneys collected from the dead fish were positive for IHNV based on a PCR assay (data not shown), confirming that the pathogen causing death was IHNV. The relative survival rates of the three immunized groups (from high to low) were 91.67% (for the BPL group), 83.33% (for the BEI group), and 79.17% (for the formaldehyde group) (Fig. 2).

Cumulative mortalities of immunized and control rainbow trout (n = 50) challenged with 10LD50 of the live IHNV (LD50 = 10−3.36, 0.1 mL) by intraperitoneal injection 56 days after immunization. IHNV, Infectious hematopoietic necrosis virus.
Discussion
Specific antibodies play crucial roles in providing protection against some viral diseases (7,10). A high level of anti-IHNV antibodies can render trout resistant to subsequent challenges with a wild-type virus and prevent the fish mortality observed from the IHNV DNA vaccine, pIHNwG, encoding the IHNV G (18,19). In this study, we observed that the RPS rates and the specific IgM antibody titers were positively correlated among the three immunized groups. The BPL group had the highest level of IgM in both their serum and mucus samples after vaccination, while the formaldehyde group had the lowest level of IgM, which was likely due to antigen modification caused by the inactivating agent. Formaldehyde is a polyfunctional alkylating agent that modifies carboxyl, hydroxyl, and sulfhydryl groups of proteins, as well as adenine and guanine (21). Thus, it is possible that formaldehyde destroys the immunogenicity of viral antigens, while BPL and BEI primarily introduce nucleic acid adducts, principally 7-(2-carboxyethyl) guanine (8,30).
The early mucosal responses in trout may consist of heterogeneous forms of Ig being produced, which differ in their characteristics relative to serum Ig (9). Another report showed that after parasitic infection by Ceratomyxa shasta, the specific IgM present in the intestinal mucus of rainbow trout did not change significantly, although the level of the specific IgT was 51 times higher compared with the control group (33). This agrees with the results of our study showing that the specific IgM levels were relatively low in the early stage after immunization. Since the fourth week after immunization, the level of IgM in the mucus increased rapidly, showing a trend almost parallel to the changes in the serum. This phenomenon was likely a result of the passive transfer of serum-derived antibodies into mucosal sites, and we will attempt to confirm this hypothesis in future studies.
In a study by Anderson et al., the formaldehyde-inactivated whole-virus vaccine was effective 7 or 11 days after the vaccination of rainbow trout, but it performed inconsistently when tested at later time points, and the BEI-inactivated whole-virus vaccine was not effective in vaccinated rainbow trout challenged by a waterborne exposure to IHNV 56 days (15°C) after immunization. For comparison, in this study, formaldehyde- and BEI-inactivated whole-virus vaccines performed effectively when trout were challenged at 56 days (16°C) after immunization, which may be because of the enhanced vaccination. Previous studies have shown that rainbow trout are able to produce an immunological memory to fish disease bacterins administered by flush exposure (3). Moreover, the immune and nutritional status of the fish can have a great impact on the overall efficacy of the resulting immune response. Although the BPL-inactivated oil-adjuvant whole-virus vaccine was the most effective, the high cost of manufacturing this vaccine and the limited study in China on BPL use as an inactivation agent contribute to the small market for BPL-inactivated vaccines.
In conclusion, the BPL-inactivated IHNV HLJ-09 oil-adjuvant vaccine was the most effective vaccine candidate when compared to the effects of the formaldehyde- and BEI-inactivated vaccines. It stimulated higher levels of the specific IgM in both the serum and mucus (body surface and gills) of rainbow trout and significantly protected against IHNV. These findings provide an experimental basis for developing an inactivated vaccine against IHNV.
Footnotes
Acknowledgments
This study was supported by the National Natural Science Foundation of China (Grant No. 31372568; 31511130137), the Northeast Agricultural University of China Talent Foundation (Grant No. 2012RCB66), and the National Science and Technology Pillar Program of China (Grant No. 2013BAD12B02).
Author Disclosure Statement
No competing financial interests exist.