
Abstract
Nucleotide-binding domain leucine repeats (NLRs) are required for the recognition of various molecules that are expressed within microbes and are able to actuate appropriate immune responses via activation of cytokines. The current study evaluates the expression levels of NLRP1 and NLRC4, which are components of inflammasomes, in chronic hepatitis B (CHB) virus-infected patients. This study recruited two series of CHB patients (each contained 60 patients) and 60 healthy controls. Real-time polymerase chain reaction (PCR) was employed to evaluate mRNA expression levels of NLRP1, NLRP3, and NLRC4 as well as hepatitis B virus (HBV)-DNA copy number. Serum levels of liver markers were also used to evaluate the patients. Hepatitis B envelope antigen (HBeAg) and hepatitis B surface antigen (HBsAg) were also examined in all patients to evaluate infection. The data showed that expression levels of NLRC4 and NLRP1 were not significantly different in circulating monocytes of CHB patients when compared with those of healthy controls. Furthermore, the data indicate that mRNA levels of NLRP1, NLRP3, and NLRC4 were also not altered in CHB patients regardless of HBV-DNA copy numbers/mL and HBeAg status. The data revealed that mRNA expression levels of NLRP1 and NLRC4 were not altered in CHB patients, suggesting that these genes are not responsible for the impaired immune responses against HBV observed in these patients.
Introduction
C
Materials and Methods
Subjects
This case–control study was performed at the Immunology of Infectious Diseases Research Center, Rafsanjan University of Medical Sciences between 2014 and 2015. This study was performed on two series of CHB patients. The first group was 60 CHB patients with various HBV-DNA copy numbers, and the second series comprised the CHB patients who were all under 20,000 HBV-DNA copy number/mL. The mRNA levels of NLRP1 and NLRC4 were measured in the first series, and the mRNA levels of NLRP3 were measured in the latter series. The control group consisted of health workers who were selected with the same sex and age of the CHB patients. Inclusion criteria for the patients group were as follows: All patients were seropositive to HBsAg for at least 6 months and had been referred to the hepatitis clinic of the Kerman University of Medical Sciences.
CHB patients infected with hepatitis A, C, D, and E virus, cytomegalovirus, human immunodeficiency virus and Epstein–Barr virus, or with liver diseases, previous liver transplantation, cirrhosis, alcoholic liver disease, Wilson syndrome, psychiatric conditions, or evidence of hepatocellular carcinoma (HCC) were excluded from this study. Patients using antiviral and immunosuppressive drugs, as well as pregnant or breast-feeding patients, or those having an age <18 years or >55 years, were excluded from the study.
Whole-blood samples were collected in 5.5 mL tubes with and without EDTA anti-coagulant for mRNA extraction and isolation of serum, respectively. The CHB disease was diagnosed by an infectious disease specialist according to the “Guide of Prevention and Treatment in Viral Hepatitis” protocol (13). The ethical committee of the Rafsanjan University of Medical Sciences approved the protocol of this study, and all of the attendees filled out and signed an informed consent form before sample collection.
Laboratory-based HBV markers
The protocols and methodology for detection of serological HBV markers and HBV-DNA quantification are described in our previous study (15).
RNA extraction, reverse transcription, and quantitative real-time PCR
The methods for total RNA isolation, cDNA synthesis, and real-time PCR are previously described (3). Briefly, mRNA was purified from peripheral blood immune cells by using a commercial kit (Cinnclon, Tehran, Iran) and was converted to cDNA by using a commercial kit from Pars Tous Company (Mashhad, Iran). Real-time PCR was performed by using a BIO-RAD instrument (BIO-RAD CFX96). The specific primers used for the gene analysis of each of the inflammasome genes, as well as beta actin as a housekeeping gene, are shown in Table 1. Gene expression was evaluated by using the ratio for each query transcript from a patient sample against the housekeeping gene. This was calculated by using the Livak method (2exp-ΔΔCt).
Data analysis and statistical methods
The data normality distribution was checked by using the Kolmogrovo–Smirnov (KS) test and according to the non-normal data distribution, the Mann–Whitney and One-Sample KS Test within the SPSS software version 18 were used for a comparison between two groups. The relationship between serum levels of liver enzyme and expression levels of NLRP1, NLRP3, and NLRC4 was analyzed by using the Pearson correlation coefficient test. A p value <0.05 was considered significant.
Results
Detection of HBsAg/HBeAg and quantification of HBV-DNA copy numbers
The patients and controls were matched regarding age and sex variations (Table 2). All patients examined in this study were initially recruited in a previous investigation, and the data for hepatitis B surface antigen (HBsAg)/HBeAg positivity as well as HBV-DNA copy numbers were reported elsewhere (15). In brief, the results showed that all of the patients in the first series were HBsAg positive, with 26.6% (16 out of 60) of the patients being positive for HBeAg as well. The CHB patients in the second series were HBsAg positive, with 3.3% (4 out of 60) of the patients being positive for HBeAg as well. The patients in the first series were grouped according to HBV-DNA copy numbers/mL, and these groups were classified as either <20,000 (28 patients), between 20,000 and 100,000 (14 patients), or greater than 100,000 (18 patients). However, all of the patients in the second series were under 20,000 HBV-DNA copy numbers/mL.
CHB, chronic hepatitis B.
Expression of target genes
Our results indicated that mean relative expression levels of NLRP1 in the peripheral blood immune cells of CHB patients and healthy controls were 49.69 and 48.10, respectively. Statistical analysis revealed that the difference between these groups was not significant (p = 0.782). The results also showed that mRNA levels of NLRP3 significantly (p < 0.001) increased in CHB patients (72.81) in comparison to healthy controls (39.72). The mRNA levels of NLRC4 were 50.00 in CHB patients and 48.86 in healthy controls (p = 0.844) (Fig. 1), and they were not statistically different. Furthermore, the results demonstrated that mRNA levels of NLRP1 (p = 0.958) and NLRC4 (p = 0.078) were not significantly different between CHB patients carrying different viral loads (Fig. 2, NLRP3 data are not shown here, because they were only evaluated in the series 2 patients, see next). For the following analysis, which includes NLRP3 data, all the samples were taken from the series 2 patients (i.e., patients with <20,000 HBV-DNA copy numbers/mL). Expression levels of NLRP1 (p = 0.241), NLRP3 (p = 0.652), and NLRC4 (p = 0.390) were not different between HBeAg-positive and -negative CHB patients (Fig. 3).
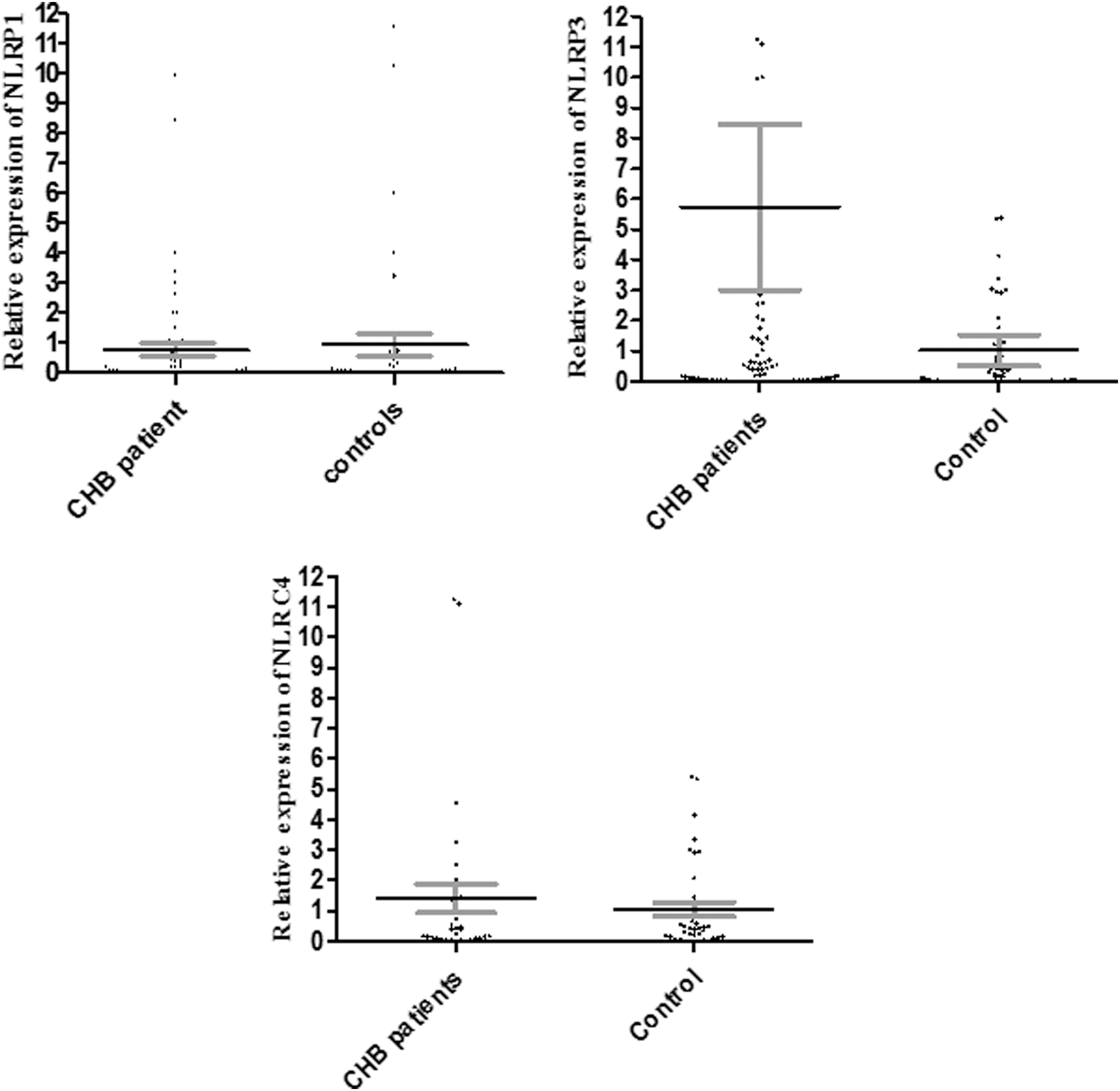
Expression levels of NLRP1 and NLRC4 in CHB patients in comparison to healthy controls. The figure indicated that mRNA levels of NLRP1 (p = 0.782) and NLRC4 (p = 0.844) were not changed in CHB patients in comparison to healthy controls. The dots in the dot plot figures indicate the situation of each participant, and error bars indicate the mean and SE, respectively. CHB, chronic hepatitis B.

mRNA levels of NLRP1 and NLRC4 in CHB patients with various HBV-DNA copy numbers/mL; <20,000 A, between 20,000 and 100,000 B, and greater than 100,000 C. The analysis of data indicated that expression levels of NLRP1 (p = 0.241) and NLRC4 (p = 0.390) were not significantly different between CHB patients carrying different viral loads. The dots in the dot plot figures indicate the situation of each participant, and error bars indicate the mean and SE, respectively. HBV, hepatitis B virus.
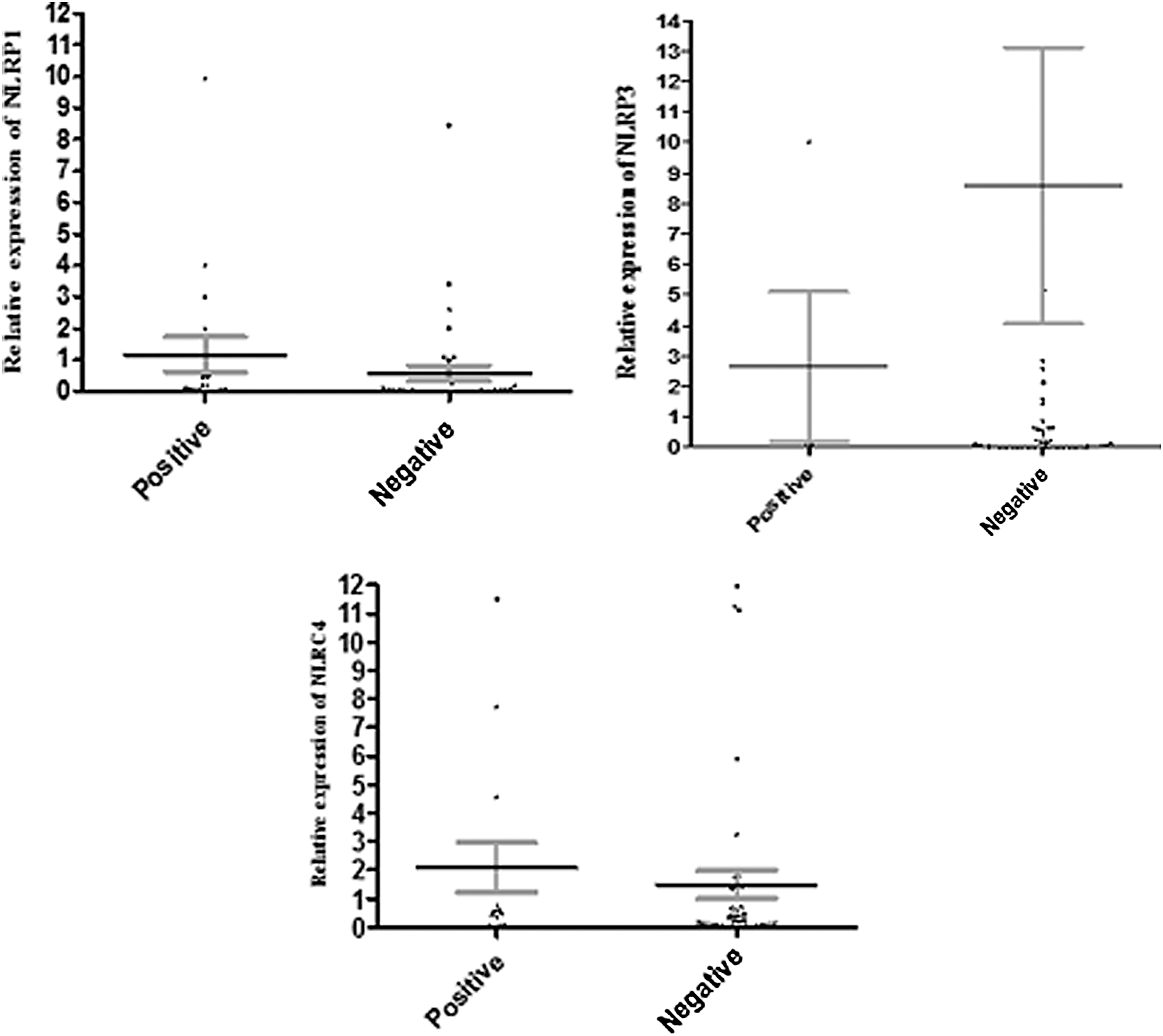
Relative expression of NLRP1 and NLRC4 in HBeAg-positive and -negative CHB patients. The results revealed that the mRNA levels of NLRP1 (p = 0.241) and NLRC4 (p = 0.390) were not different between HBeAg-positive and -negative CHB patients. The dots in the dot plot figures indicate the situation of each participants, and error bars indicate the mean and SE, respectively. HBeAg, hepatitis B envelope antigen.
Serum levels of liver function tests
The results revealed that serum levels of liver enzymes were within normal ranges in the patients (Table 3). The results also demonstrated that there is no significant relationship between expression levels of NLRP1, NLRP3 (except alkaline phosphatase [ALP], p = 0.003), and NLRC4 with serum levels of liver enzymes (Table 4).
ALT, alanine aminotransferase; ALP, alkaline phosphatase; AST, aspartate aminotransferase.
The data suggest that serum levels of ALT, AST, ALP, and total and direct bilirubin were not associated with expression levels of NLRP1 and NLRC4; whereas serum levels of ALP have a poor positive correlation with NLRP3.
SGOT, serum glutamic-oxaloacetic transaminase; SGPT, serum glutamic-pyruvic transaminase.
Discussion
NLRC4, NLRP1, and NLRP3 are components of the inflammasome family and they play fundamental roles in the activation of caspase-1 and, in turn, the stimulation of IL-1 and IL-18, which are important molecules required to combat HBV (19,20). Our results demonstrated that the mRNA levels of NLRC4 and NLRP1 expressed in the peripheral blood immune cells did not change but NLRP3 increased in CHB patients when compared with healthy subjects. Based on the present results, it may be hypothesized that NLRP3, but not NLRC4 and NLRP1, is an important intracellular sensor for HBV recognition and, hence, induction of immune responses against the virus via activation of IL-1β and IL-18. However, the data do not assess activation of NLRP1 and NLRP1 or recruitment to multi-protein complexes (12) and they do not exclude the possibility that downstream activation of IL-1 and IL-18 is stimulated through these processes. To the best of our knowledge, this is the first study that evaluated expression levels of NLRC4, NLRP3, and NLRP1 in peripheral blood immune cells of CHB patients. However, Wu et al. reported on the expression of another component of inflammasomes, the Absent In Melanoma-2 (AIM 2), and showed that it was significantly reduced in CHB patients in comparison to acute infected patients (22). Interestingly, the investigators claimed that the expression of AIM-2 was positively correlated with HBV clearance (22). Our previous investigations revealed that mRNA levels of AIM-2 were not changed between CHB patients and healthy controls; however, mRNA levels of caspase recruitment domain (ASC) and caspase-1 were significantly reduced in CHB patients when compared with healthy controls (9). ASC is a downstream signaling molecule of inflammasomes and based on the interaction of its Pyrin domain with the Pyrin domain of inflammasomes, it will be activated, which leads to activation of caspase-1 and cleavage of pro-IL-1 and pro-IL-18 (12). Based on the fact that NLRP1, NLRP3, and NLRC4 are also engaged in the activation of pro-IL-1 and pro-IL-18 (12), hence, it may be concluded that the specific tolerance to HBV in CHB patients may be related to the inhibition of molecules downstream of inflammasomes. Moreover, Manigold et al. reported that hepatitis B core antigen (HBcAg) induces the secretion of IL-18 from immune cells as a result of induction of caspase-1, which itself is a target of inflammasomes (14). Therefore, it appears that a complete HBV particle can inhibit expression of inflammasomes responses but not HBcAg alone. It has been documented that NLRP3 is an important component of the inflammasomes (17) and our results suggest, in our cohort, that NLRP3 was the only component of the inflammasomes that shows increased expression during immune responses to hepatitis B. According to our previous study, ASC was decreased in CHB patients; hence, two hypotheses are plausible. Although mRNA levels of NLRP3 are elevated in the CHB patients, it appears that decreased expression of ASC disrupts the NLRP3 pathway. Otherwise, NLRP3 may use an unknown intracellular pathway to induce immune responses. Clarification is required before these data may improve our understanding of CHB pathogenesis. A better understanding of the inflammasomes and their downstream signaling will help with the diagnosis of CHB and provide relevant data to assist in the design of molecular-based therapeutic protocols.
IL-1 and IL-18 are downstream targets of inflammasome signaling pathways; compelling evidence demonstrates that IL-1 and IL-18 not only play key roles in the initiation and progression of immune responses against HBV but also participate in hepatitis B-associated complications such as liver cirrhosis and HCC (23,24). There is no evidence regarding the roles of inflammasomes during CHB infection; however, some studies have evaluated the relationship between inflammasomes and other viral hepatitis, including HCV. For example, Negash et al. reported that in chronic hepatitis C patients the elevated levels of serum IL-1B were associated with NLRP3 (16). Based on the fact that several HCV-infected patients were suffering from liver complications, including cirrhosis and HCC, hence, it seems that the inflammasomes may play some role in the pathogenesis of the liver complications but their precise action during hepatitis B infections needs to be explored by further investigation.
The current results revealed that expression levels of the inflammasomes' component transcripts were not significantly different among CHB patients, regardless of the HBV-DNA copy numbers, nor was there a correlation between HBeAg-negative and HBeAg-positive CHB patients. Therefore, it appears that HBV replication is not associated with altered expression of NLRP1, NLRP3, and NLRC4. In addition, the results showed that only ALP has a poor positive correlation with NLRP3, whereas serum levels of liver enzymes and total and direct bilirubin are not associated with NLRP1, NLRP3, and NLRC4. Hence, it may be concluded that the expression of NLRP3, but not the other inflammasome components, may be associated with hepatitis B complications such as liver damage. It appears that more studies on inflammasomes need to be performed to demonstrate the roles of these molecules in the pathogenesis of hepatitis B.
Our previous studies revealed that, in contrast to inflammasomes, expression levels of other intracellular receptors, including stimulator of interferon genes (STING) (11) and melanoma differentiation-associated protein 5 (MDA5) (6), were significantly decreased in our CHB population. Thus, it seems that HBV targets several PRRs to induce immune tolerance to its antigens.
Footnotes
Acknowledgments
The authors take this opportunity to thank all of the CHB patients and healthy controls who participated and cooperated in this study. This project was derived from an MSc thesis (ID: 37) from Rafsanjan University of Medical Sciences.
Author Disclosure Statement
No competing financial interests exist.