
Abstract
There are over 350 million chronic carriers of hepatitis B virus (HBV) in the world, of whom about a third eventually develop severe HBV-related complications. HBV contributes to liver cirrhosis and hepatocellular carcinoma development. Remarkable progress has been made in selective inhibition of HBV replication by nucleoside analogs. However, how to generate protective antibody of HBsAb in HBV-infected patients after HBV-DNA becomes negative still remains a challenge for scientists. In this study, we show that OmpC-HBsAg ‘a’ epitope chimeric protein vaccine can break HBV tolerance and induce protective immunity in HBV transgenic mice based on mimicking T cell-independent antigen to bypass T cells from the adaptive immune system. The antibodies induced by the vaccine have the ability to prevent HBV virion infection of human hepatocytes.
Introduction
H
At present, the nucleoside analogs such as entecavir, tenofovir, and lamivudine show the evidence that HBV DNA in serum of HBV-infected patients could be undetected after antiviral therapy. However, how to generate protective antibody of HBsAb (also known as HBsAg-seroconversion) in HBV-infected patients after HBV-DNA becomes negative still remains a challenge for scientists.
In the humoral immune system, B cells arise from two distinct lineages: B-1 and B-2 cells. The majority of IgG are produced by B-2 cells. There are mainly three reasons of humoral immune tolerance caused by HBsAg in HBV-infected patients. First, HBsAg is a T cell-dependent (TD) antigen and HBsAg-specific B-2 cells need the help of CD4+ Th2 cells. However, HBsAg-specific CD4+ Th2 cells have function defects in HBV infection (1). Second, there exist HBsAg-specific suppressor T cells inhibiting B-2 cells to produce HBsAb (2). Third, recent studies on CD4+CD25+ regulatory T cells (Treg cells) revealed that Treg cells not only inhibit the humoral and cellular immune responses to HBV but also affect the prognosis of patients with HBV infection (3 –7). All this boils down to one point that B-2 cells need the assistance and regulation of T cells to generate antibody against HBsAg, a TD antigen in adaptive immune response.
How to induce humoral immune response to HBsAg in HBV patients who are under HBV-induced immune tolerance status? One of the strategies is to bypass T cells from the adaptive immune system. HBsAg-specific B-2 cells require interaction with CD4+ Th2 cells for their activation. On the contrary, B-1 cells, appearing early in embryogenesis, respond to T cell-independent (TI) antigens without Th2 help and Treg suppression. Compared with conventional B-2 cell, B-1 cell is a distinct B cell subset with unique features in its source, distribution, surface marker, gene expression, and function (8 –10).
B-1 cells play an important role in innate immunity. B-1 cells can generate neutralizing antibodies against mucosal pathogens within 48 h after encountering antigens, exhibiting TI activation (11). It was reported that B-1 cells have the regulatory function in the adaptive immune response (12,13). In B-1 cell-deficient mice, specific protective IgG antibodies cannot be induced by vaccination. This phenomenon can be rescued through inputting B-1 cells, suggesting their role in the adaptive immune system. In addition, B-1 cells also have memory function, which means they can proliferate and differentiate into plasma cells in the absence of T cells after encountering TI antigen, providing long-term or even lifetime protective antibodies (14 –17).
Since HBsAg belongs to TD antigen, B-2 cells need the help of CD4+ Th2 cells to produce HBsAb. However, HBsAg-specific CD4+ Th2 cells could not provide the second signal in HBV patients who are under HBV-induced immune tolerance status, leading to HBsAg-specific B-2 cell anergy. In contrast, B-1 cells can generate antibodies after encountering TI antigen in the absence of T cells. Therefore, B-1 cells are immune competent cells in immune tolerance status. Although the isotypes of antibodies generated by B-1 cells are mainly IgM, class switch to IgG can occur under specific cytokine environment, such as IFN-gamma (17,18).
In this study, we reported that OmpC-HBsAg ‘a’ epitope chimeric protein vaccine could break HBV tolerance and induce protective immunity in HBV transgenic mice based on mimicking TI antigen. The antibodies induced by OmpC-HBsAg ‘a’ epitope chimeric protein vaccine have the ability to prevent HBV virion infection.
Materials and Methods
Plasmid constructions
The amino acid sequence of OmpC-HBsAg ‘a’ epitope chimeric protein is shown in Figure 1D. To generate this chimeric gene, we cloned the OmpC gene from Salmonella typhimurium genome DNA using the primers, 5′-ATGAA AGTTAAAGTACTGTCCCTCCTGGTACCAGC-3′ and 5′-TTAGAACTGGTAAACCAGACCCAGCGCTAC-3′, and then ligated into pET28b. The full length of OmpC open reading frame (ORF) is 1,137 bp. We insert adr ‘a’ epitope of HBsAg, which contains a sequence of 24 amino acids, CTSPAQGTSMFPSCCCT KPSDGNC, into the site between Asp181 and Thr182 located in the fourth loop of OmpC. The OmpC-HBsAg ‘a’ epitope chimeric gene was constructed by overlap extension polymerase chain reaction (PCR). The primers are as follows: forward1: 5′-ACATGCCCATGGGTAAAGTTAAAGTACT GTCCCTCCTGGTACCAGC-3′, reverse1: 5′-CGTGCAGTTTTCACCGCTCACGCTGCCGTTTTTGC-3′, reverse2: 5′-GAGGTTCCTTGAGCAGGACTC GTGCAGTTTTCACCG CTC-3′, reverse3: 5′-GTACAGCAACAAGAGGGAAACATAGAGGTTCCTTGAGCAGGAC-3′, reverse4:
5′-CTCGGATCCGCTTAGAACTGGTAAACCAGACCC AGCGCTAC-3′, forward2: 5′-ACGGAAACTGCACCAAC GGTCGCAGCCTGC-3′, forward3: 5′-TTGCTGTACAAA ACCTTCGGACGGAAACTGCACCAACG-3′, and forward4: 5′-GGAACCTCTATGTTTCCCTCTTGTTGCTGTACAAA ACCTTCGGAC. To generate the DsbA gene, we cloned the DsbA gene from Salmonella typhimurium genome DNA using the primers, 5′-CTCGGATCCTCGGAGAGAGTTGATCATGAAAAAGATTTG-3′ and 5′-CCCAAGCTTATTTTTTATCAACCAAATATTTCACAGTATCAGCATACTG-3′, and then ligated into pET32b.
The expression of OmpC-HBsAg ‘a’ epitope chimeric protein in bacteria
The plasmids of PET28b-OmpC-HBsAg ‘a’ epitope chimeric protein and PET32b-DsbA were cotransfected into BL21 (DE3) and bacteria were grown in medium supplemented with 100 μg/mL of ampicillin and 50 μg/mL of kanamycin at 37°C. When A600 reached 0.4, IPTG was added into the medium (0.1 mM) at 16°C, 160 rpm/min, and the induction duration is 20 h.
Animals and vaccination
HBV transgenic (HBV25-8, H-2Kb) female mice, 6–8 weeks old, were purchased from Shanghai Biomodel Organism Science & Technology Development Co., Ltd. The genotype of HBV25-8 is C and the serotype of HBsAg is adr. The animals were randomly divided into three groups (n = 6 each) and kept under controlled temperature (22–23°C) with 12-h light–12-h dark cycle, having free access to water and food. Care and maintenance of all animals were in accordance with the guidelines of Institutional Animal Care and Use Committee of Nantong University. For immunizations, BL21 bacteria (1 × 108/50 μL) expressing OmpC-HBsAg ‘a’ epitope chimeric proteins were injected i.p. once a week for seven times.
On weeks 4 and 8, the orbital sinus blood from the mice was collected and the neutralizing HBsAb IgM and IgG were examined. The mice in control groups are injected with recombinant (yeast) hepatitis B vaccine (Engerix-B, HBsAg group) or BL21 bacteria transfected with empty vectors (vector group). For depletion of CD4+ T cells, HBV transgenic mice were daily injected with 300 μg of L3T4 anti-CD4 mAb i.p. during a period of 3 days. This treatment was shown as depletion of the corresponding splenic T cell subset.
Flow cytometry and fluorescence activating cell sorter analysis
To measure the expression of HBsAg ‘a’ antigenic determinant on BL21, 3E7 antibody (sc-57785, 3E7; Santa Cruz Biotechnology, Inc.) was used for staining. Approximately 1 × 108 live cells were washed with PBS, then stained with a mouse monoclonal antibody 3E7 specific for HBsAg ‘a’ antigenic determinant for 30 min at 4°C. After three washes, the secondary FITC-labeled goat anti-mouse IgG was incubated with cells for 20 min at 4°C. The stained cells were analyzed by FACSCalibur (BD Biosciences) and FlowJo software (Tree Star).
Immunofluorescence
After IPTG induction, BL21 bacteria were collected and gently resuspended in 2% FBS PBS. The mouse primary monoclonal antibody against HBsAg ‘a’ epitope (sc-57785, 3E7; Santa Cruz Biotechnology, Inc.) was used at 1:100 dilution, 4°C. The secondary antibody was an FITC-conjugated goat anti-mouse IgG antibody (Santa Cruz Biotechnology, Inc.) used at 1:1,000 dilution, 4°C. Then, they were washed and transferred to a 24-well cell culture plate and detected by a fluorescence microscope (Olympus IX-71).
Enzyme-linked immunosorbent assay
Induced IgM anti-HBsAg antibody in serum bound with HBsAg to form the antigen–antibody complex. To detect the level of IgM anti-HBsAg antibody, we adopted the following method: IgM is an antibody with low affinity, and a lower pH value benefits the dissociation of antigen-IgM–antibody complex. Therefore, we cross-linked a high-affinity IgG anti-HBsAg mAb with agarose and added them excessively into the diluted serum. We lowered the pH value by addition of acid to dissociate the Ag-Ab complex and then restore the pH value to 6.0 gradually. The dissociated HBsAg preferentially bound to high-affinity IgG anti-HBsAg mAb-agarose. After centrifuge, the supernatant was free of HBsAg and unbound IgM anti-HBsAg antibody can be detected. For enzyme-linked immunosorbent assay (ELISA), HBsAg proteins expressed in CHO cells were used to coat the wells of a microtiter plate. Anti-mouse IgG and anti-mouse IgM antibodies were conjugated with HRP. A direct ELISA was performed to analyze the value of IgG and IgM in serum, respectively.
Quantitative real-time reverse transcriptase-PCR
Total RNA was extracted from primary human hepatocellular cells with an RNeasy Mini kit (Qiagen) using DNase digestion protocol, followed by first-strand cDNA synthesis using the Sensiscript RT kit (Qiagen) according to the manufacturer's instructions. Quantitative real-time reverse transcriptase (RT)-PCR was performed on a PRISM 7500 system (Applied Biosystems) with designed primer pairs and the SYBR green PCR master mix (Applied Biosystems). β-Actin was used as reference gene for sample normalization of primary human hepatocellular cells. Relative quantitation of gene expression was calculated using the ΔCT method based on signal intensity of PCR results, according to the following formula: 2−ΔCT = [2−(sample Ct − normalizer Ct)], where Ct was the threshold cycle of the quantitative PCR. The primer sequences used were 5′-GACTTAGTTGCGTTACACCC TTTC-3′ (hβ-actin sense); 5′-TGCTGTCACCTTCACCG TTC-3′ (hβ-actin antisense); 5′-GTGTCTGCGGCGTTTT ATC-3′ (HBsAg sense); and 5′-TCCGTCCGAAGGTTTT GTA-3′ (HBsAg antisense).
Statistics
All statistical analyses in this study were performed with Student's t test, and a probability of less than 95% confidence limit (p < 0.05) was considered to be significant. Data are presented as mean ± standard deviation.
Results
The construction of OmpC-HBsAg ‘a’ epitope chimeric protein
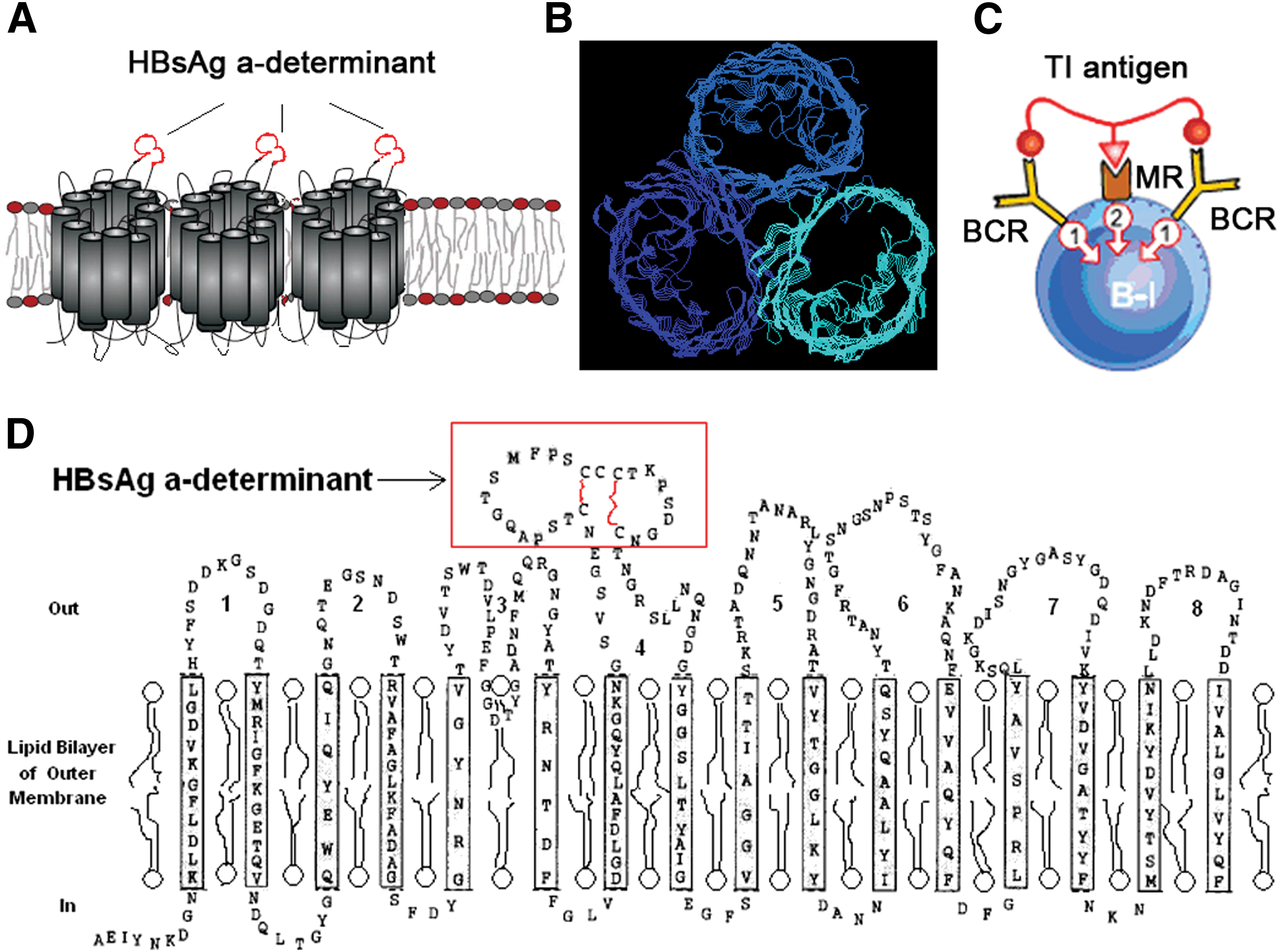
OmpC is an outer membrane channel protein of bacteria and it has an ability to activate B-1 cell without T cell assistance. Its three-dimensional structure is a homotrimer (Fig. 1A, B). It contains a specific epitope and a B cell mitogen. The specific epitope on OmpC can interact with B-1 cell and bind to B cell receptor (BCR), which provide the first signal for B-1 cell activation, while the latter can interact with mitogen receptor on B-1 cell, offering the second signal (Fig. 1C). We inserted adr ‘a’ epitope of HBsAg, which contains a sequence of 24 amino acids, CTSPAQGTSMFPS CCCTKPSDGNC, into the site between Asp181 and Thr182 located in the fourth loop of OmpC (Fig. 1D).
Formation of HBsAg ‘a’ conformational epitope on bacterial membrane with assistance of DsbA protein
The disulfide isomerase A (DsbA) of Salmonella typhimurium is an efficient enzyme in the periplasm that catalyzes the formation of disulfide bonds between cysteine residues as they fold. This allows HBsAg ‘a’ epitope to quickly find the correct arrangement of disulfide bonds in their fully folded state to form the accurate conformation. With the help of DsbA, the conformation of HBsAg ‘a’ antigenic determinant was formed. As shown in Figure 2A and Supplementary Figure S1 (Supplementary Data are available online at

Besides, the conformational formation of HBsAg ‘a’ antigenic determinant was also observed by fluorescence microscopy. The binding of fluorescent antibody on membrane of BL21 was apparently observed in the group of cotransfected DsbA with OmpC-HBsAg ‘a’ plasmid, but weak binding in the group of transfected OmpC-HBsAg ‘a’ vector alone, indicating that DsbA can effectively aid the conformation forming of HBsAg ‘a’ epitope on the membrane of prokaryotic bacteria. Then, we used β-mercaptoethanol to reduce disulfide bonds for testing whether the conformation forming of HBsAg ‘a’ epitope depends on DsbA. After β-mercaptoethanol treatment, the binding of fluorescent antibody was abolished and the fluorescence disappeared subsequently (Fig. 2B).
Breaking of immune tolerance in HBV transgenic mice by OmpC-HBsAg ‘a’ epitope chimeric protein vaccine
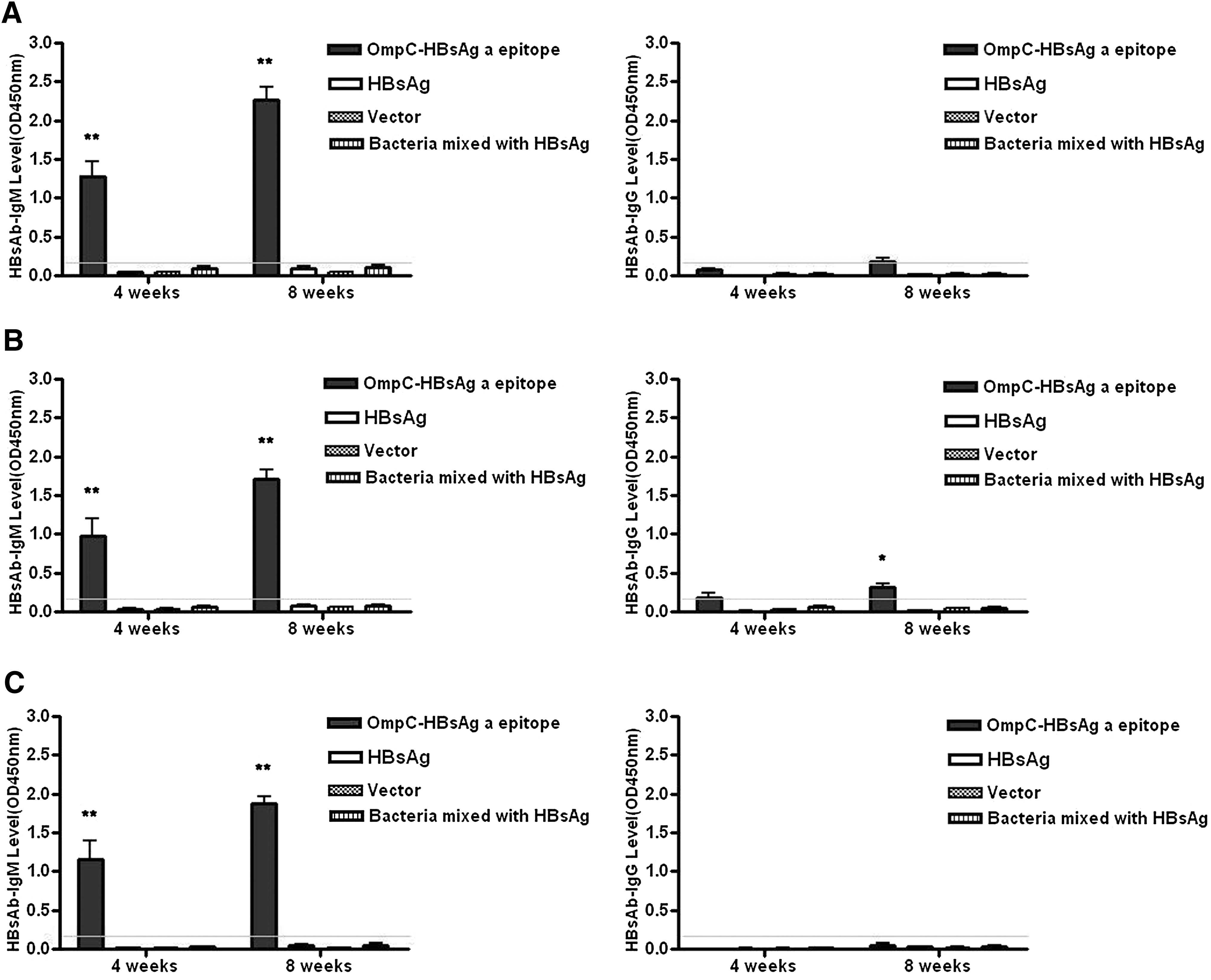
Immunization with bacteria expressing OmpC-HBsAg ‘a’ epitope chimeric protein on the membrane induced IgM anti-HBsAg antibody responses, which could be detected in the week after the first injection and increased after subsequent administrations in HBV transgenic mice. After 4 weeks, the mean OD value of IgM was 1.283 in the chimeric protein vaccine group (Fig. 3A), however, that of IgM neutralizing antibody was 0.046 in the control group, which was immunized with naive HBsAg protein. Eight weeks later, the mean OD value of IgM in the chimeric protein vaccine group increased to 2.268, but there was no obvious change in those of three control groups. In addition, no statistical difference was observed among four groups in IgG anti-HBsAg antibody responses, but it slightly increased after the injection of IFN-gamma in the chimeric protein vaccine group (Fig. 3B and Supplementary Fig. S2).
IgM anti-HBsAg antibody production was not significantly influenced when CD4+ T cells were deleted by injection of anti-CD4 mAb, indicating that antibody generation was T cell independent (Fig. 3C). Besides, purified OmpC-HBsAg ‘a’ epitope chimeric protein can only induce a low level of anti-HBsAg IgM in HBV transgenic mice, but when OmpC-HBsAg ‘a’ epitope chimeric protein was expressed on the membrane of the bacteria, high level of anti-HBsAg IgM can be induced in these mice (Fig. 4). There are two mechanisms for this phenomenon. One mechanism is that multiple intracellular signaling pathways are activated upon the recognition of pathogen-associated molecular patterns (PAMPs) by B-1 cells, which can offer extra signals for gene transcription and antibody production. Therefore, PAMPs on the surface of the bacteria assist OmpC-HBsAg ‘a’ epitope chimeric protein for anti-HBsAg IgM induction. Another mechanism is that various trimeric OmpC-HBsAg ‘a’ epitope chimeric proteins expressed on the surface of bacterial membrane could gather together in the same lipid raft, which enhances signal transduction of BCR. On the contrary, purified OmpC-HBsAg ‘a’ epitope chimeric protein lost the capability of gathering, thus activation of the BCR signal pathway by single protein is weaker.
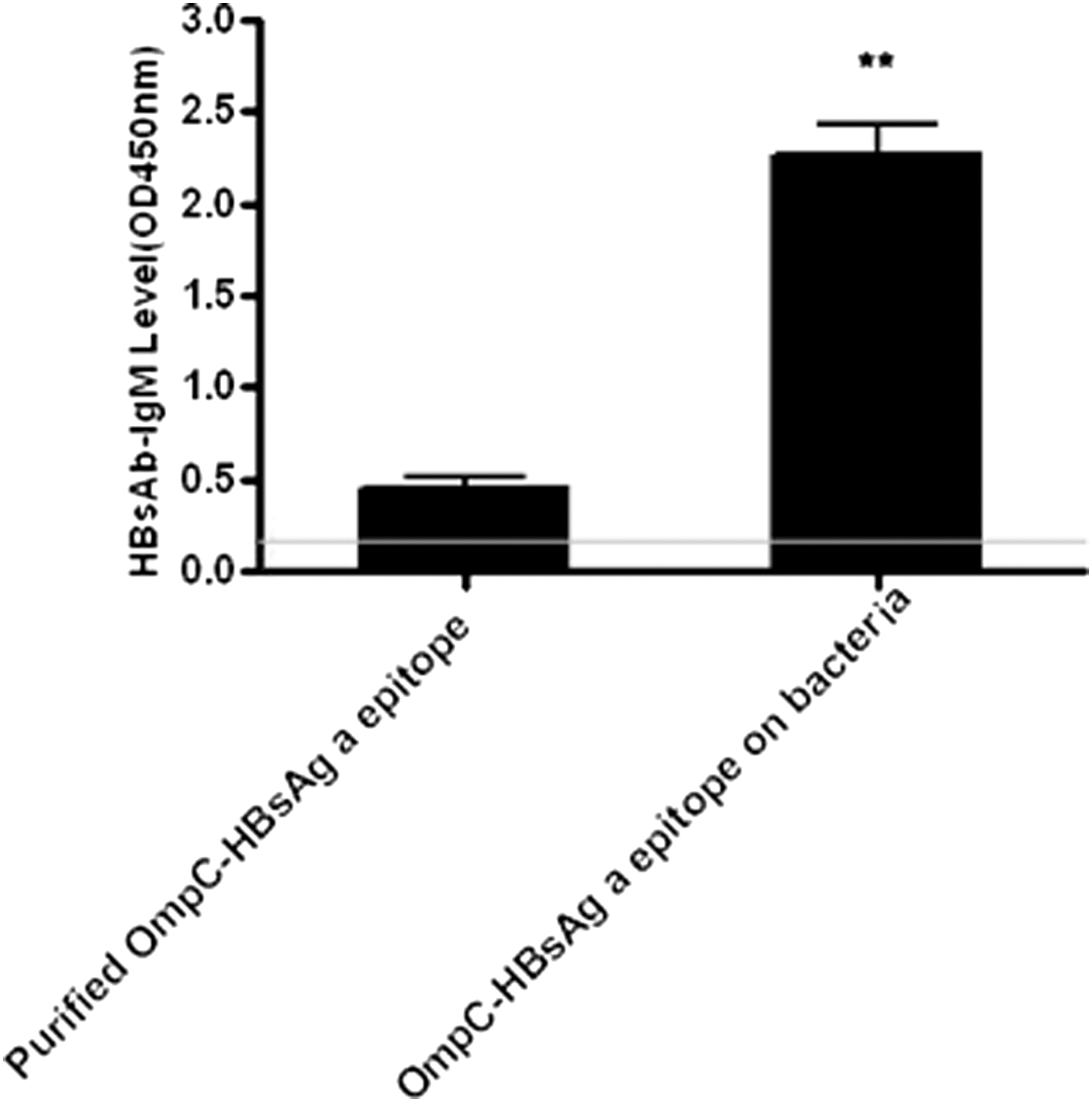
Purified OmpC-HBsAg ‘a’ epitope chimeric protein or OmpC-HBsAg ‘a’ epitope chimeric protein expressed on the membrane of bacteria induced different levels of anti-HBsAg IgM in HBV transgenic mice on week 8. **P < 0.01 versus purified OmpC-HBsAg ‘a’ epitope chimeric protein group.
Prevention of HBV infection using IgM anti-HBsAg antibody induced by OmpC-HBsAg ‘a’ epitope chimeric protein vaccine
In HBV transgenic mice, the isotype of antibodies induced by OmpC-HBsAg ‘a’ chimeric protein vaccine is mainly IgM. It should be confirmed whether the isotype IgM has the ability to prevent HBV virion infection. To address this question, the serum samples from immunized mice were collected to test HBV virion infection in primary human hepatocytes. The mRNA expression of HBsAg in infected hepatocytes was examined by real-time PCR. We found that the ratios of HBsAg gene expression in the chimeric protein vaccine group to that in control groups were at significantly low level (chimeric protein vaccine group/naive HBsAg protein group = 0.909%; chimeric protein vaccine group compared/vector group = 0.767%), indicating that the IgM anti-HBsAg antibody induced by OmpC-HBsAg ‘a’ chimeric protein vaccine can not only neutralize the HBV virion but also prevent HBV infection of human hepatocellular cells (Fig. 5).
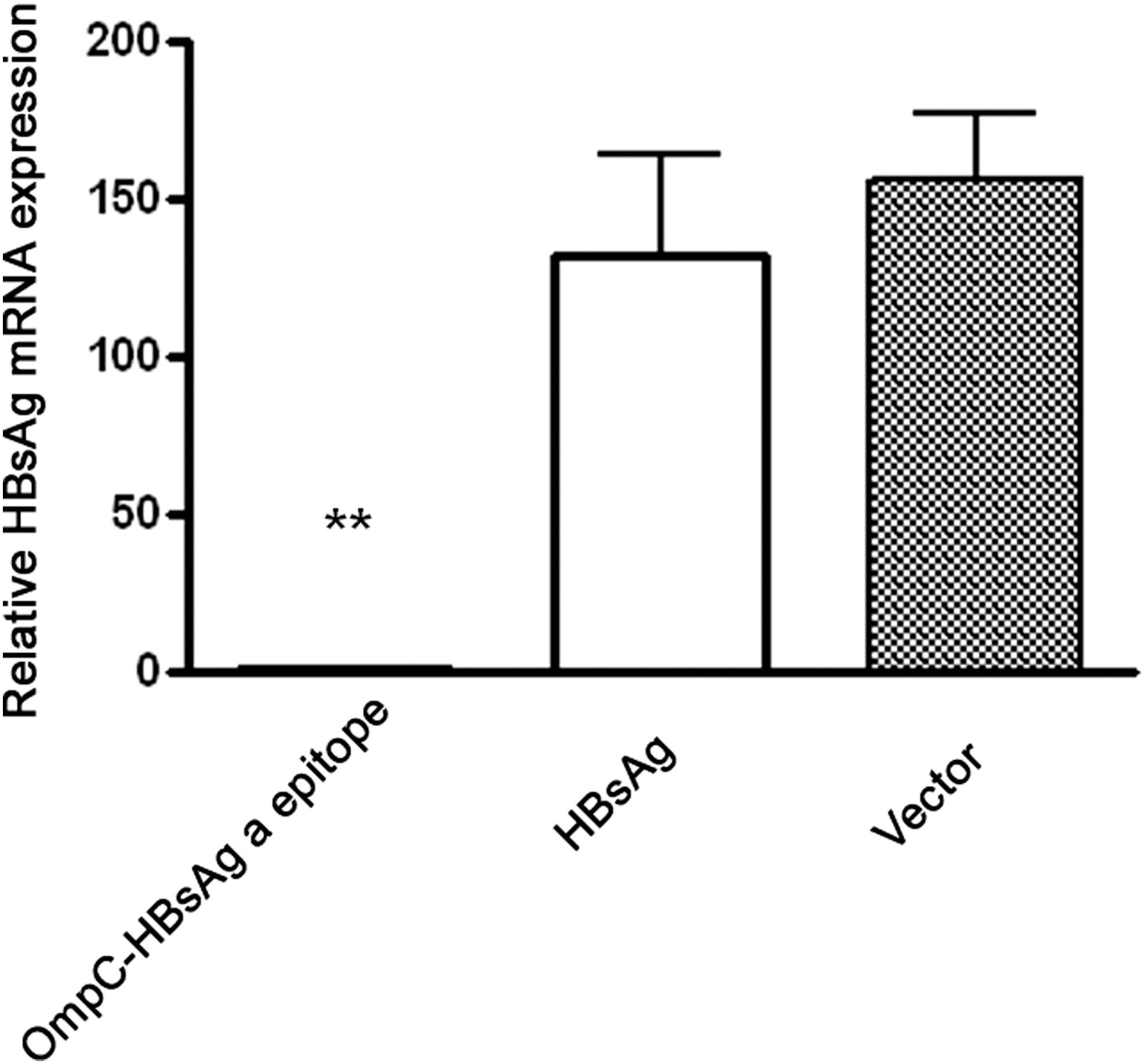
The serum samples from immunized HBV transgenic mice were collected to block HBV virion infection in primary human hepatocellular cells. Primary human hepatocytes were cultured in X-VIVO20 supplemented with 10 ng/mL EGF, 18 μg/mL hydrocortisone, and 40 ng/mL dexamethasone. HBV virion was obtained from the serum of the HBV patient with HBsAg adr subtype matching (HBV DNA = 3.5 × 106 copies/mL). We removed HBsAg from the immunized mice sera by using high-affinity IgG anti-HBsAg mAb-Agarose and lowering of pH. All the serum samples were diluted 1:100. The sera from humans and mice were incubated for 1 h, and then added into the medium of cell culture. For HBV viral infection on primary human hepatocytes, 1.5% PEG 8000 was present during the inoculation period for 16 h at 37°C as previously described by Gripon et al. and Schulze et al. (28,29). Subsequently, the culture medium was changed with fresh medium. The cells were cultured continuously for 48 h and then were harvested for RNA extraction. Real-time polymerase chain reaction was used for quantification of HBsAg mRNA expression. **P < 0.01 versus HBsAg group or vector group.
Discussion
As TI antigens, porins are β-barrel outer membrane proteins through which small solutes and metabolites diffuse. According to their different functions, porins are mainly divided into osmotic pressure channel protein such as OmpC and OmpF, phosphate channel protein such as PhoE, sucrose channel protein such as ScrY, and maltose channel protein such as LamB. Porin is a B-1 cell mitogen, which can polyclonally activate human and murine B-1 cells to produce antibodies in vitro. The three-dimensional structure of porin has been determined as a conserved homotrimer, which forms repetitive B cell epitopes.
Vordermeier and Bessler extracted porins from Salmonella that could induce high titers of effective antibodies in nude mice (19,20). The concentration of the antibodies was much higher than that of pokeweed mitogen (PWM). Several literatures revealed that porin directly induces B7 expression, which is an important costimulatory molecule on the surface of B cells. Porin also upregulates MHC II and enhances TLR2 and TLR4 expression (21 –24). In addition, porin can stimulate monocytes to produce TNF-α, IL-1α, and IL-6 and induce T cells to secrete IL-4, triggering Th2-type immune response (25,26). Due to its strong immunogenicity, porin has been successfully applied to human vaccines and adjuvants. When OmpC and OmpF extracted from Salmonella were administrated to healthy volunteers, anti-porin antibodies and T cell proliferation were detected 7 days postadministration (27). Secundino et al. demonstrated that sustainable antibody response was induced in BALB/c and C3H/HeJ mice after being immunized with OmpC and OmpF (15). Moreover, the antibody level induced by OmpC was higher than that by OmpF, indicating that OmpC from outer membrane of Salmonella is a potent immunogen.
In this study, we inserted the epitope of HBsAg ‘a’ antigenic determinant into the membrane protein of OmpC to develop a chimeric protein vaccine, which can activate the innate immune cell, B-1 cell, to produce HBsAb, thereby breaking the immune tolerance caused by HBV. For B cell epitopes from HBsAg three-dimensional structures, there is a hydrophilic peptide, S-(124-147aa), also called ‘a’ epitope, which is a highly conserved amino acid sequence on the surface of HBsAg. This epitope is easily accessible to BCR and also efficiently neutralized by HBsAb. Therefore, it is an important B cell epitope of HBsAg, which can induce protective antibodies.
In this epitope, there exists a stable ring-like secondary structure through forming two disulfide bonds (Cys124-Cys137 and Cys139-Cys147). If the two disulfide bonds did not correctly form, the ‘a’ epitope could not be shaped and recognized by antigen-specific B cell, which would be induced to secrete specific antibodies. However, the biggest obstacle in protein expression in prokaryotic organisms is the correct arrangement of disulfide bonds in their fully folded state. Moreover, the redox potential in the cytoplasm of prokaryotic organisms prevents the proper fold of protein. The disulfide bond can be formed only when the protein is transported to the periplasmic space. DsbA protein, a periplasmic disulfide isomerase, is localized in the periplasmic space and plays a key role in the formation of the protein disulfide on the bacterial membrane.
In addition, PAMPs on the membranes of bacteria such as lipopolysaccharides, flagellin, and peptidoglycan can be specifically recognized by the innate immune cells through pattern recognition receptors. After recognition, multiple intracellular signaling pathways are activated, which can offer an extra signal for gene transcription. It is important for B-1 cells to self-activate and product antibodies.
In summary, OmpC-HBsAg ‘a’ epitope chimeric protein vaccine can break HBV tolerance and induce protective antibody based on mimicking TI antigen. These findings elucidate that TD antigen can be transformed to TI antigen for bypassing T cells. For clinical application of OmpC-HBsAg ‘a’ epitope chimeric protein vaccine, it could be made as a live attenuated bacteria vaccine. The live attenuated bacteria vaccines such as attenuated cholera vaccine have been successfully applied in clinical immunology in the past. Besides, OmpC-HBsAg ‘a’ epitope chimeric protein and appropriate PAMP protein can be purified separately and recombined together in the same liposome as a proteoliposome vaccine in the future.
IFN-gamma can induce class switch from IgM to IgG. IFN-gamma has been approved by FDA (USA) for the treatment of metastatic renal cell carcinoma, trauma, atopic dermatitis, and granulomas, also by PMDA (Japan) for the treatment of renal cell carcinoma and mycosis fungoides, and by MHRA (UK) for the treatment of rheumatoid arthritis, but there is no clinical trial in CHB. The side effects of IFN-gamma are similar to that of IFN-alpha. IFN-alpha has been successfully applied in patients with CHB and therefore it is also possible to use IFN-gamma in clinical trials to treat CHB in the future.
In addition, we consider that OmpC-HBsAg ‘a’ epitope chimeric protein vaccine has greater efficacy under some conditions compared with conventional HBsAg vaccines. For example, when HBsAg in serum becomes negative after administration of RNA interference-based drug, such as ARC-520, the immune tolerance in patients still exists and conventional HBsAg vaccines cannot induce significant antibody response, while OmpC-HBsAg ‘a’ epitope chimeric protein vaccine has the ability to induce antibody response in this case. Moreover, conventional HBsAg vaccines usually fail to induce a significant humoral immune response in most non-CHB patients during hypoimmunity, such as long-period administration of prednisone, severe sepsis, and septic shock, thus OmpC-HBsAg ‘a’ epitope chimeric protein vaccine will come in handy at this time.
Footnotes
Acknowledgments
This work was supported by the projects of Nantong Social Development (S2009021), Nantong standardization of clinical diagnosis and treatment of key diseases (HS2014072), and National Nature Science Foundation of China (81272708).
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.