
Abstract
Vaccine immunogen with expanded T cell coverage for protection against HIV-1 diversity is the need of the hour. This study was undertaken to examine the ability of T cells to respond to a broad spectrum of potential T cell epitope (PTE) peptides containing variable as well as conserved sequences that would most accurately reflect immune responses to different circulating strains. Set of 320 PTE peptides were pooled in a matrix format that included 40 pools of 32 peptides per pool. These pools were used in interferon-γ enzyme-linked immunospot assay for screening and confirmation of HIV-1 PTE Gag-specific T cell immune responses in 34 HIV-1 seropositive Indian individuals. “Deconvolute This” software was used for result analysis. The dominant target in terms of magnitude and breadth of responses was observed to be the p24 subunit of Gag protein. Of the 34 study subjects, 26 (77%) showed a response to p24 PTE Gag peptides, 17 (50%) to p17, and 17 (50%) responded to p15 PTE peptides. The total breadth and magnitude of immune response ranged from 0.75 to 14.50 and 95.02 to 1,103 spot-forming cells/106 cells, respectively. Seventy-six peptides located in p24 Gag were targeted by 77% of the study subjects followed by 51 peptides in p17 Gag and 46 peptides in p15 Gag with multiple variants being recognized. Maximum study participants recognized PTE peptide sequence Gag 271→285NKIVRMYSPVSILDI located in p24 Gag subunit. T cells from HIV-1-infected individuals can recognize multiple PTE peptide variants, although the magnitude of the responses can vary greatly across these variants.
Introduction
T
Frequent and robust immune responses against immunodominant regions of HIV-1 Gag are well-documented with many studies demonstrating inverse correlation of plasma viremia with the magnitude of anti-Gag CD8+ T cell responses (5,30). Kaushik et al. have also proposed that a stronger and broader CTL immune response in terms of magnitude and breadth of responses was found to be directed against the Gag subunit (17). Few other studies have also pointed out that low viral load is associated with T cell recognition of multiple epitopes within specific Gag regions (9,18). Hence, the collective data suggest that the inclusion of immunodominant sequences of Gag cross-reactive between different clades in future candidate vaccines would be the logical step in tackling HIV-1 diversity globally (7). In support of this, Geels et al. reported that HIV-seropositive individuals can elicit broad cross-clade HIV-1 T cell responses specifically against Gag subunit (8).
In this study, we aimed to identify the cross-clade-specific HIV-1 Gag response among HIV-1-infected Indian individuals using 15 mer peptide sequences, designated potential T cell epitope (PTE), that are designed to reflect cross reactivity of immune response against circulating strains of HIV present in target viral population. We investigated the magnitude and breadth of HIV-specific T cell response by interferon (IFN)-γ enzyme-linked immunospot (ELISpot) assay using PTE peptides. We next identified the most immunodominant and targeted T cell epitopes located in HIV-1 PTE (Gag) peptides in HIV-1 subtype C population and determined the cross-reactivity of T cell responses by using the Los Alamos Immunology Database.
Materials and Methods
This study was conducted at the Immunology Division, Department of Microbiology, All India Institute of Medical Sciences (AIIMS), New Delhi, a tertiary healthcare centre. The study was approved by the Institutional Ethics Committee and written informed consent was obtained from all subjects before entering the study.
Study population
Thirty-four treatment-naive HIV-seropositive patients recruited from Y.R. Gaitonde Center for AIDS Research and Education (YRG CARE) clinic, Chennai, were enrolled in the study. At the time of enrolment, all the study subjects were asymptomatic and had CD4 T cells count greater than 500 cells/μL and viral load less than 50,000 copies/mL. Ten HIV-negative healthy individuals (six females, four males) with no history of past infection were included in the study as healthy controls.
CD4+ T cell counts and viral RNA loads
Clinical data were collected prospectively from all the study subjects. After taking informed consent, 5 mL of whole blood was collected from study subjects and healthy controls by venipuncture in K3EDTA vacutainer tubes. Blood samples were processed within 2 hours of collection. CD4 T cell count was performed on a FACS Count™ system (Becton Dickinson, San Jose, CA). EDTA plasma samples were quantified for HIV-1viral load using the COBAS TaqMan HIV-1 Test, v2.0 (Roche Diagnostics, IN), and the lower limit of detection was 47 copies/mL.
HIV-1 PTE Gag peptides
A lyophilized panel of 320 peptides that permit expression of the most frequent PTE embedded in sequences of circulating HIV-1 strains of HIV-1 worldwide were kindly provided by Dr. Clive Gray from South Africa (courtesy NIH AIDS Research and Reference Reagent Program; NARRRP, Bethesda, MD). These peptides are 15 amino acid (a.a.) in length and contain naturally occurring 9 a. a. sequences that are potential T cell determinants in HIV. The peptides were dissolved in DMSO at a concentration of 20 mg/mL (stock) and 40 pools were made with a final concentration of 1.5 μg/mL per peptide for stimulation. For second round assays, individual peptides were also tested at a final concentration of 1.5 μg/mL.
Sample preparation
Blood samples were collected in EDTA-treated vacutainers (Becton Dickinson, San Jose, CA) by venipuncture. Peripheral blood mononuclear cells (PBMCs) were isolated using density gradient centrifugation over Ficoll-Histopaque (Sigma Aldrich). The cells thus collected were cryopreserved in liquid nitrogen and were later used for IFN-γ ELISpot assay.
Deconvolution of HIV peptide-specific T cell response
“Deconvolute this” software programme designed by Mario Roederer (Vaccine Research Center, NIAID, NIH, readily provided upon request) was used for matrix design, pooling of peptides, and determination of second round assays based on screening results. The matrix included 40 pools of 32 peptides per pool. These 40 pools were used in the IFN-γ ELISpot assay for screening the possible immunodominant regions. A 4 × 4 matrix-based strategy was then used to pool these peptides. Second round experiments were performed based on the number of pools that test positive during the screening round and peptides were then individually retested in a second ELISpot assay.
IFN-γ ELISpot assay
Commercially available Human IFN-γ ELISpot kits (R&D Systems, Minneapolis, MN) were used to screen HIV-1 PTE Gag-specific T-cell immune responses as per the manufacturer's instruction. PBMCs stimulated with phytohemagglutinin (PHA at 2 μg/mL; Sigma–Aldrich, St. Louis, MO) and kit control run in duplicate wells were used as positive controls to ensure that the cells were responsive, and cells without antigenic stimulation served as background controls. ELISpot plate reader (Carl Zeiss Vision GmbH, Hallbergmoos, Germany) was used for estimation of number of spots per well. Results were expressed as spot-forming cells (SFC)/106 PBMCs. The threshold for a positive response was set at greater than 100 SFC/106 PBMCs. Peptides that came positive in both four pools and three pools were confirmed individually by a second ELISpot assay to rule out the possibility of missing any potential positive peptides. Samples from 10 HIV-seronegative individuals recruited from YRG CARE clinic, Chennai, were also assayed in parallel to confirm any nonspecific reactivity to the synthetic peptides.
Statistical analyses
All statistical analyses were performed using the STATA 7.0 software. The median and range were calculated for all the study variables. The bar diagrams and scatter plots were drawn using Microsoft Excel programme (version 5.0; Microsoft Corp.). Correlations were performed using Spearman's ranked test. Data are expressed as median (interquartile range [IQR]) unless otherwise indicated. A p-value of <0.05 was considered statistically significant in all statistical analyses performed. All data were expressed as medians with IQR.
Results
The median age of the study population was 31 (21–44) years. The median CD4+ T cell counts and viral load was 842 cells/mm3 (605.8–1,036) and 6,070 copies/mL (496–25,408), respectively. The demographic and clinical characteristics of study subjects are enumerated in Table 1.
ND, not detected.
HIV-specific T cell responses by IFN-γ ELISpot assay using mosaic antigen set
We investigated HIV-specific T cell immune response using IFN-γ ELISpot assay against PTE Gag peptides. A total of 26 study subjects responded to these mosaic antigens, while the remainder did not recognize any of the peptides tested. The total magnitude of immune response with the median response to PTE Gag peptides was found to be 299.4 (95.02–1,103) SFC/106 PBMCs. Similarly, study subjects targeted PTE peptides differentially with the median breadth of 4.5 (95.02–1,103) peptide/subject. The magnitude and breadth of HIV-specific T cell responses recognizing individual PTE Gag peptides are shown in Figure 1.
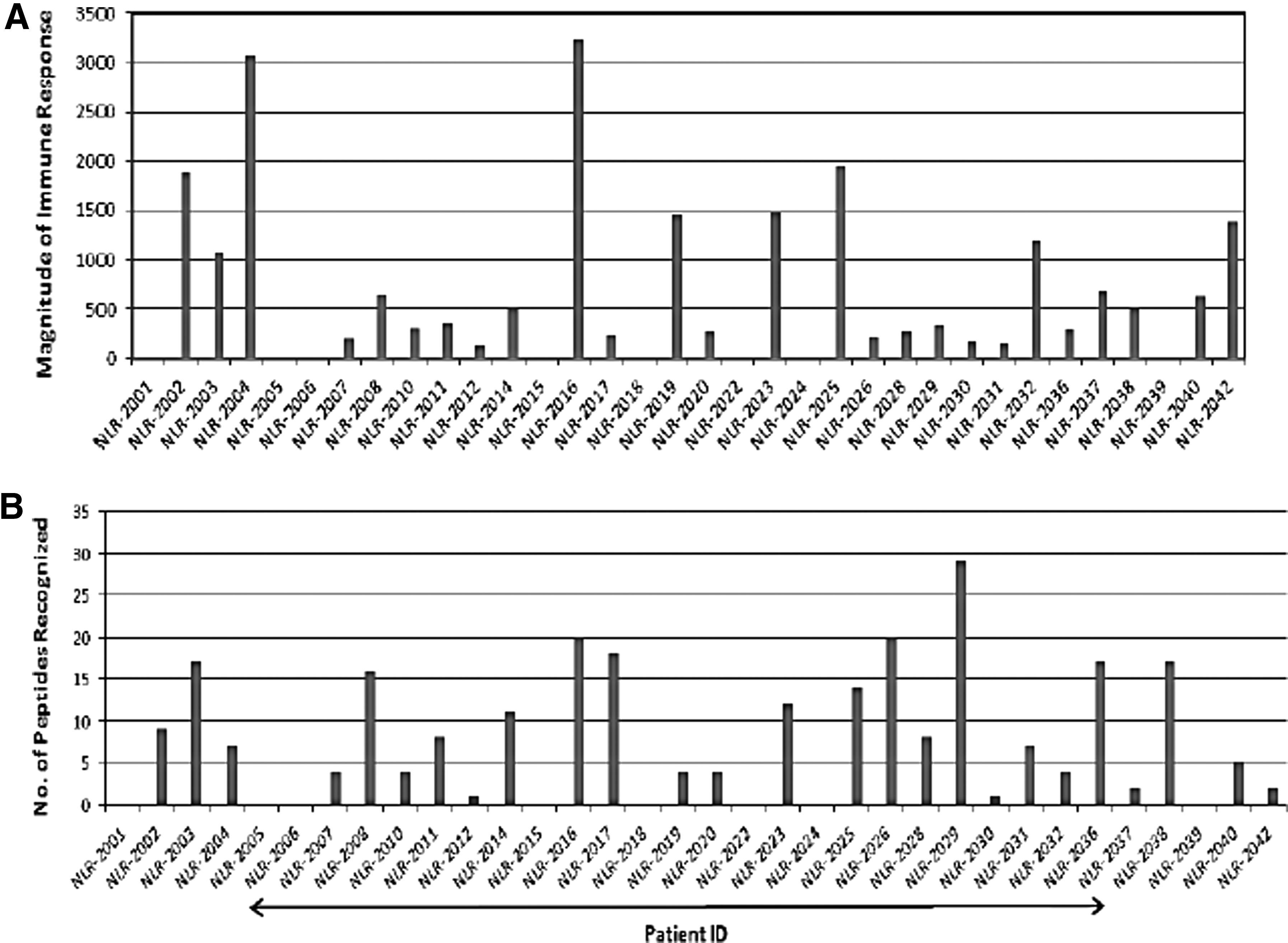
Graphical representation of magnitude and breadth of immune response against PTE Gag peptides. X axis of each graph represents the study participants, while bars correspond to magnitude and breadth of HIV-specific responses
Out of 320 peptides, 166 peptides were recognized by study population within the Gag region with majority, and 29/166 (17.46%) recognized by only one participant. p24 HIV-1 Gag subunit (76/166; 45.78%) was found to be the dominant target in terms of magnitude and breadth of responses followed by p17 Gag (51/166; 30.7%) and p15 Gag subunits (46/166; 27.7%) as depicted in Figure 2A–C. Some peptides were preferentially targeted over others, for instance, PTE Gag13271–285 was targeted by 7 (20.5%) participants followed by Gag214142–156 peptide targeted by six individuals (17.64%) within the p24 Gag region (Fig. 2A). In total, there were 13 peptides that were targeted by more than two individuals and strikingly all these peptides were found to be part of p24 Gag subunit.
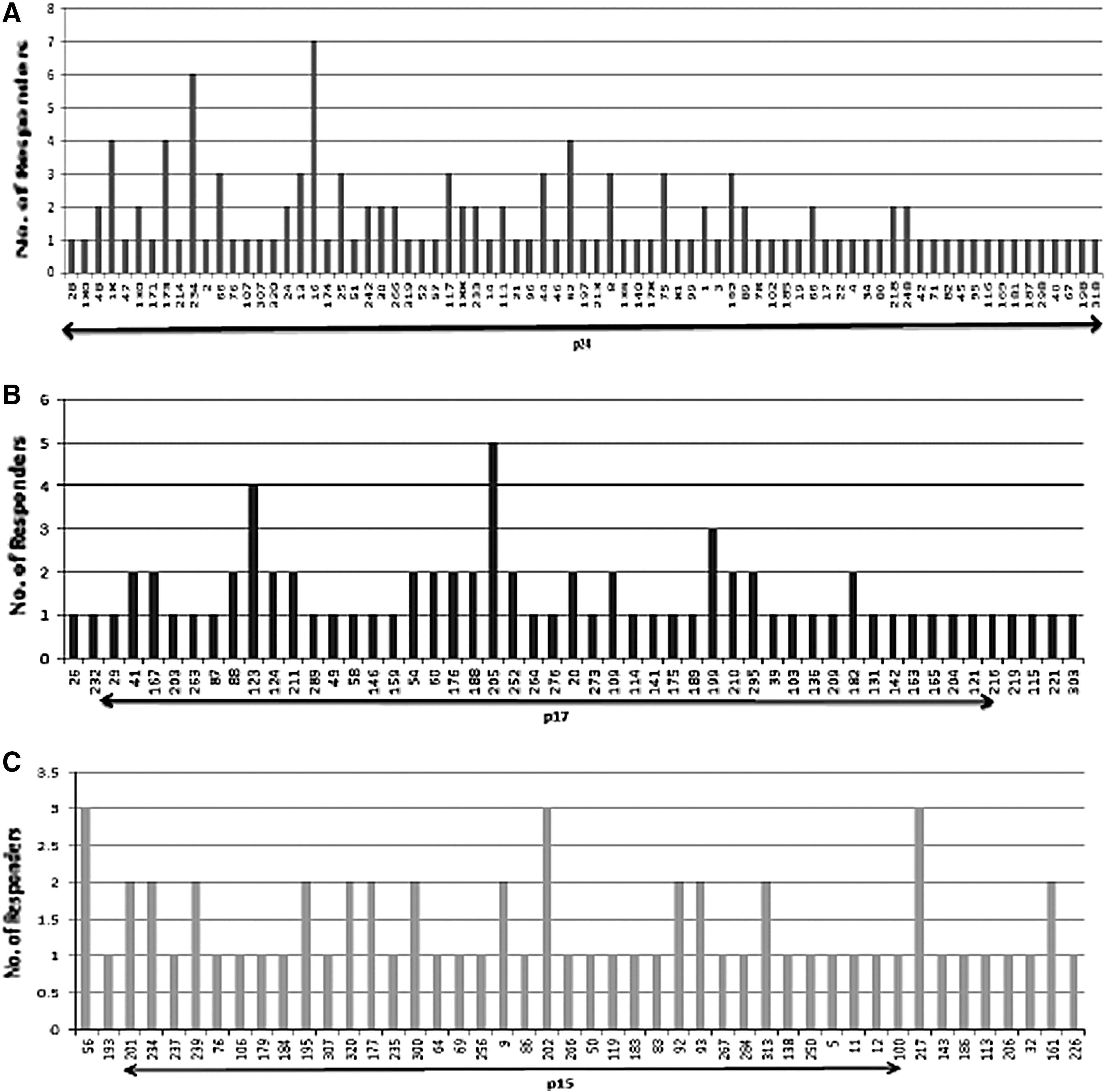
Recognition pattern of different HIV-1 Gag subunits by HIV-infected individuals represented as number of individuals responding to each Gag subunit.
Immunodominant peptides identified within PTE HIV-1 Gag
Peptides that were recognized in at least three study individuals were classified as immunodominant. A total of 18 peptides were identified as the most immunodominant peptides targeted with p24 subunit dominating the Gag region wherein a total of 13 such reactive peptides were recognized by study participants followed by three peptides in p17 region and two peptides in p15 region of Gag region respectively as shown in Table 2.
The cross-reactivity is given by the number of variants recognized for that particular peptide.
The magnitude of interferon-γ response shown in the table is the magnitude of response to the variant that gave the highest response for that particular peptide out of all variants that were reactive in the study individuals.
PBMCs, peripheral blood mononuclear cells; SFC, spot-forming cells.
Recognition of multiple variants
Study participants were found to recognize peptide variants spanning the p24, p17, and p15 region of HIV-1 Gag with up to two variants being recognized for a particular peptide within the PTE peptide set. Recognition of multiple epitope variants was highest for p24 Gag (71%), followed by p15 (67%), and last for p17 (63%) as depicted in Figure 3. Within the p24 region, no variants were recognized for 48/76 total peptides targeted by study subjects, while 29 peptides were categorized in 13 different groups with at least two variants (Tables 3
–5). Similarly, in the p17 HIV-1 Gag region, 32/51 peptides were found to have the unique sequences against which no variants were detected, while 8 groups included 19 peptides along with their variants. Last, 30 peptides without any variants were targeted in p15 region with seven groups having 16 peptides with multiple variants. Strikingly, there was heterogeneity in response for different variants of same peptides even with single amino acid change, for instance, peptides 28 (PFRDYVDR
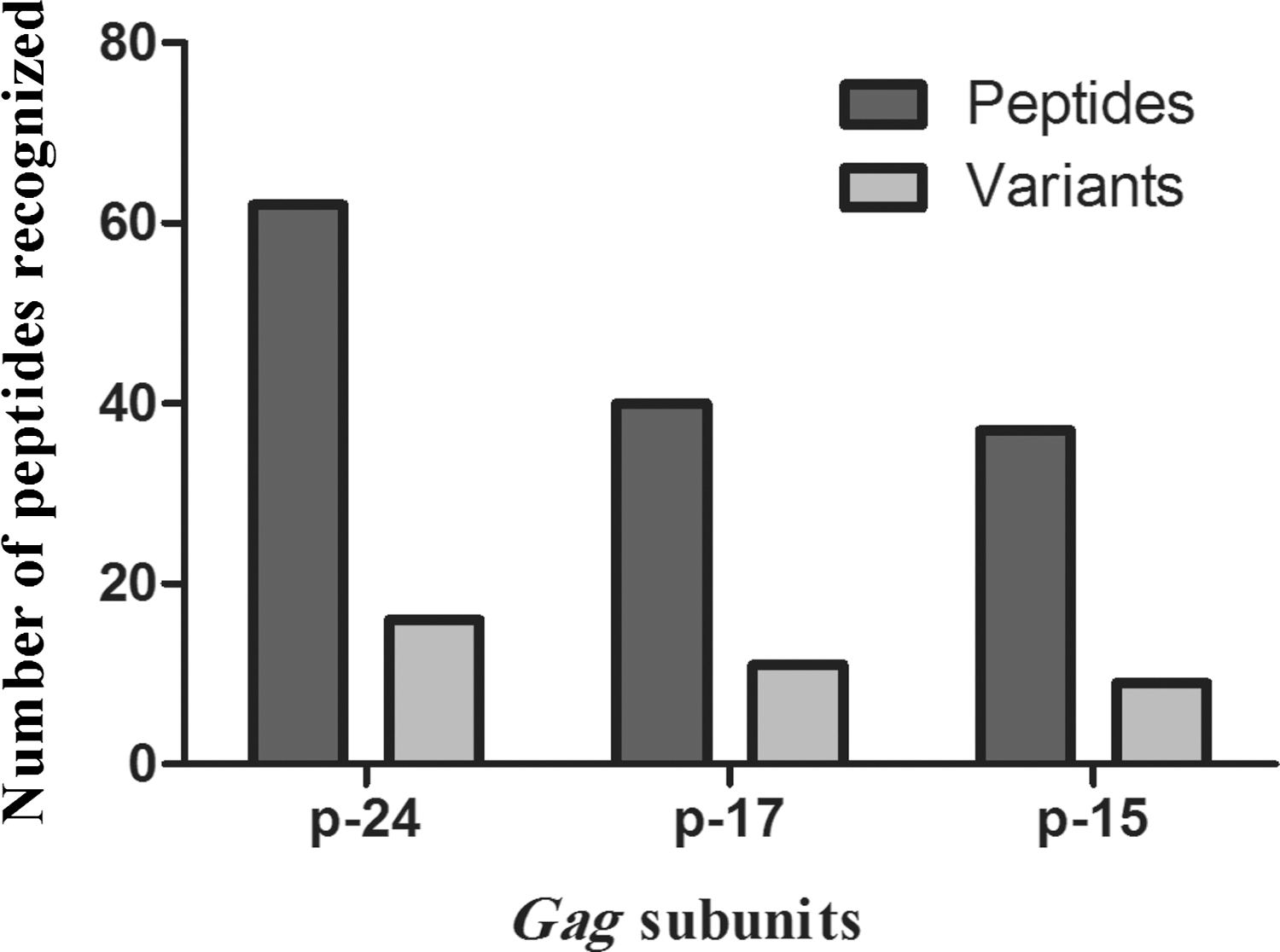
Proportion of recognized peptides with targeted variants. The number of peptides that were targeted along with variants (light gray bars) or with no targeted variants (dark gray bar) shown in p24, p17, and p15 Gag subunits.
Amino acid mismatches between corresponding peptide pairs are represented in bold.
PTE, potential T cell epitope.
Amino acid mismatches between corresponding peptide pairs are represented in bold.
Amino acid mismatches between corresponding peptide pairs are represented in bold.
An overall trend of dissimilarity was observed when we compared the magnitude of immune response among variants of the same targeted peptide in different individuals. The magnitude of immune response against these variants was found to be strikingly different, for example, NLR-2010 responded with 816.7 SFC/106 PBMCs against PTEGag28 PFRDYVDR
Shannon entropy: cross-reactive potential of different mosaic peptides
Shannon entropy score was used to determine which regions of HIV-1 Gag protein were more likely to be cross-reactive. It is a measurement of uncertainty that calculates the conserved (low entropy) as well as variations (higher entropy) of each amino acid in protein sequence, represented in the form of alignment score of each reactive peptide in p17, p24, and p15 region. Higher variability was observed for p17 (amino acids 1–145) and p15 (amino acids 379–520) Gag regions, and lower for the p24 (amino acids 146–378) region (Fig. 4).

Shannon entropy score of HIV-1 Gag regions.
Correlation between the number of PTE peptides recognized, magnitude of immune response, CD4 T cell count, and plasma viremia
We next analyzed and compared the number of peptides recognized and the total magnitude of peptide-specific immune responses with CD4 T cell count and viral load in study subjects. Interestingly, a strong positive correlation was observed between breadth and magnitude of immune response (p < 0.0001, r = 0.625), suggesting a strong role of T cell pool in eliciting a strong immune response against HIV (Fig. 5). We further investigated the correlation of disease progression markers with breadth and magnitude of immune response and no significant association was observed with either VL (Breadth: r = 0.1718, p = 0.3313; Magnitude: r = 0.1072, p = 0.5460) or CD4 count (Breadth: r = −0.06119, p = 0.7310; Magnitude: r = 0.01384, p = 0.9381).
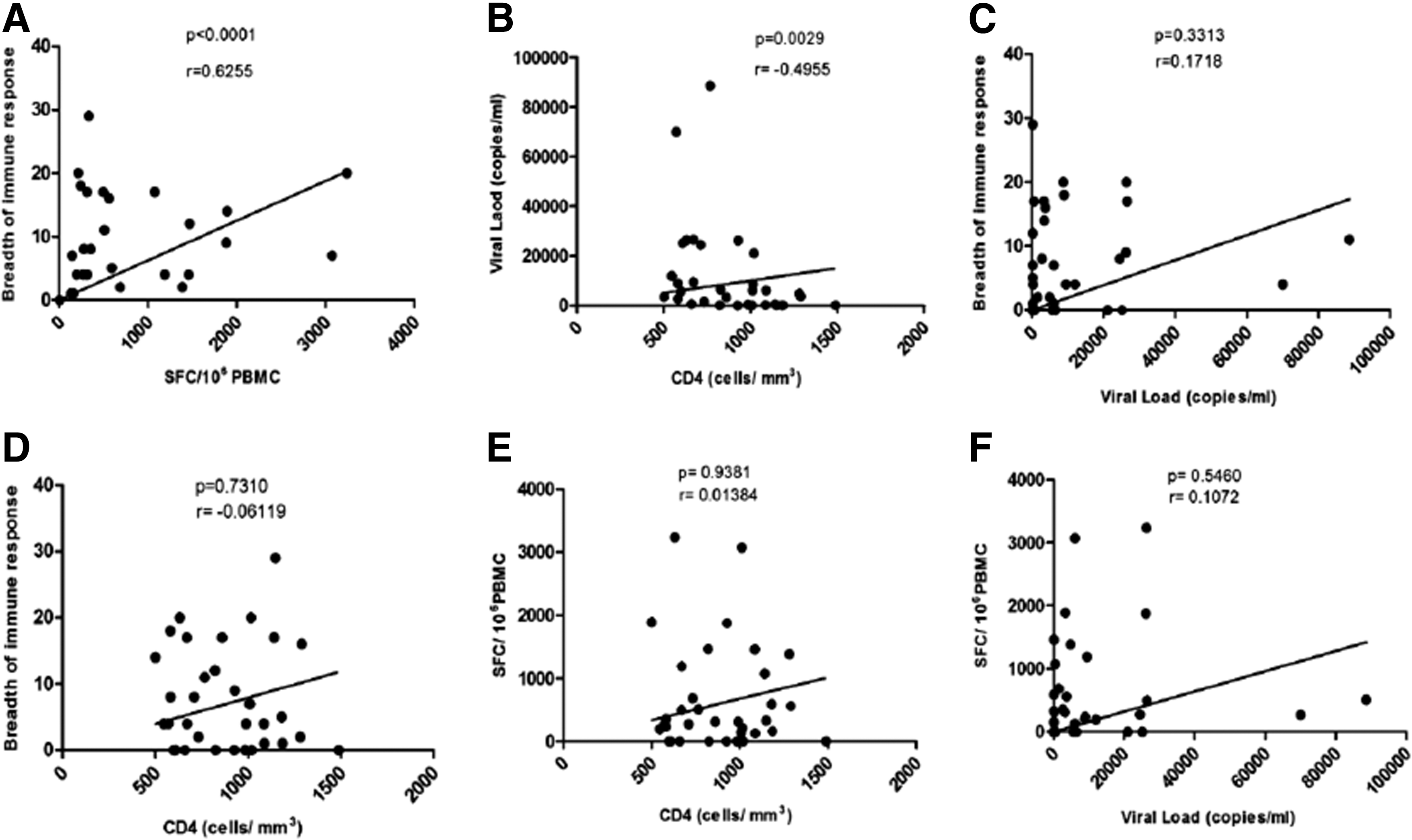
Correlation between the number of PTE peptides recognized, magnitude of immune response, CD4 T cell count and plasma viremia:
Discussion
We detected robust T cell immune response against immunodominant regions within HIV-1 Gag, one of the most conserved HIV-1 proteins with highest cross clade epitope density (1,21,25). The study revealed that T cells from HIV-1-infected Indian individuals are capable of recognizing PTE peptides along with their variants, although the magnitude of immune response exhibited (299.4 SFC/106) is comparatively lower than 2,169 SFC/106 as reported by Malhotra et al. (1). However, the breadth of response was found to be higher with 4.5 peptides/subject compared to 2.7 epitopic domains (20). The difference in both the magnitude as well as breadth of immune response could be explained by the fact that the PTE peptide set used by Malhotra et al. represented sequences from subtype B and the study was performed in subtype B-infected population only. The higher magnitude of response may be because of close proximity of peptide and viral sequences (20). Furthermore, it is interesting to note that the magnitude of response reported in this study was also found to be comparatively lower than we reported (21) earlier using the consensus HIV-1 subtype C sequences. The reason for this variation may be due to the use of subtype C-specific peptide sequences and regional variation of the study participants as this study involves participants from South India only.
Strikingly, the magnitude of immune response followed a differential pattern to different variants of the same peptide. An inverse trend was observed when a peptide and its variant were targeted by different individuals, while they followed the same pattern of immune response when targeted by the same individual. Our results are in contrast to previous findings (1,8,30), wherein no significant difference in the magnitude of immune response to peptide variants was observed. Furthermore, they advocated that both T cell receptors of HIV-specific T cells as well as their respective HLA molecules can withstand some degree of amino acid substitution in their epitopes without total loss of epitope recognition or binding in line with other studies. Our results indicate that variations or even small amino acid mismatches are not well tolerated by our study population, which could be explained by the HLA profile of study participants that plays an important role in efficient binding of these peptides to their respective Class I and class II MHC molecules. Variations introduced in peptide sequences may abrogate effective binding to respective HLA, resulting in poor immune response. Hence, it is imperative to take into consideration the distribution pattern of HLA alleles within local populations for the development of immunogen design having broad population coverage. The limited amount of sample we had restricted HLA typing of study subjects, which would have allowed us to better see the impact of HLA on targeting of peptides and their variants. These findings suggest that mosaic antigens representing Group M viruses may not be effective against all the circulating subtypes as regional and host genetic factors might impede the roadway to a universal vaccine.
A total of 18 immunodominant peptides were identified with p24 dominating the magnitude and breadth of responses. The reasons for this high frequency and magnitude of response are likely due to amino acid sequence conservation in this protein subunit. The findings are in concordance with previous reports from South Africa and India, wherein they have shown that p24 Gag subunit is highly targeted by subtype C-infected population and is a potential vaccine target (12,21,25). In addition, preferential targeting of peptides was mainly observed in p24 Gag subunit followed by p17 and p15 subunits. Our results are in concordance with Jia et al. study wherein they reported preferential targeting of conserved regions of Gag, and p24 was found to be associated with better control of viral replication (15). These findings are further supported by Hertz et al. that, HLA class I molecules also preferentially target evolutionarily conserved regions of human and viral proteomes (14). Recently published data from Cameroon, a country with greatest HIV-1 diversity, have also reported that individuals from diverse epidemics with different HLA backgrounds have T cells that recognize certain regions of HIV that are conserved across group M viruses (28). These findings clearly indicate that host genetic factors, especially the HLA molecules, play a crucial role in recognition of viral epitopes as people from different ethnicities express specific HLA molecules.
Next, we compared all the reactive peptides identified in our study with previously reported beneficial peptides by other groups. We found that only three peptides, all of them belonging to p24, were reported by three different studies. Gag PTE 13271–285NKIVRMYSPVSILDI has been reported as the most immunodominant peptide recognized in a cohort of HIV-1 subtype B or C infected Thais by Masahiko et al. in 2011 (23). Similarly, Kulkarni et al. showed that in rhesus macaques immunized with p24CE DNA vaccine, the maximum immune response was elicited against PTE Gag 3286–300RQGPKEPFRDYVDRF, and this particular peptide expresses the epitopes most efficiently recognized by cellular immunity (19). Dorrell et al. found a fourfold increase in T cell immune response against HQAAMQMLKDTINEE T cell sequence in HIV-1-infected individuals immunized with Recombinant Modified Vaccinia Virus Ankara/HIV-1 Gag Vaccine, which is also one of the immunodominant peptide PTE Gag24194–208 identified in our study population (4). However, we do not find any similarity in peptide recognition pattern in South African population with subtype C infection31. The difference in pattern of recognition could be due to the involvement of different host and viral factors, including the HLA profile of enrolled participants between South Africa and India (30). Other peptides have not been reported previously and are being documented for the first time in subtype C-infected subjects.
Next, cross-reactivity, defined in terms of number of variants recognized for a particular peptide, was analyzed in the study population. Although many studies have assessed cross-reactivity in different ethnic groups, however, the T cell immune response to Gag peptide variants has not been investigated in subtype C-infected Indian population. The PTE peptide set used in this study contains Gag sequences for subtype A, B, C, and non-ABC, represented by peptide numbers 163, 89, 145, and 152, respectively. However, we observed that the immune response was found to be directed against other sequences in the peptide set, except for two, PTE Gag16340→54 ELERFALNPSLLETT and PTE Gag 89239→253 TTSTLQEQIAWMTSN belonging to clade A and B, respectively. These results are in concordance with other reports suggesting that individuals infected with a particular clade have the ability to recognize peptides of a different subtype/clade unless it does not have variation that might interfere with TCR binding (31). The highest cross-reactivity was observed in p24 Gag subunit followed by p17 and p15 Gag region. Peptides targeted by study participants in p24 Gag region were found to be highly conserved followed by p15 region. However, peptides belonging to p17 region had high entropy score, which indicates that the probability of acquiring mutations in this particular region is very high, hence are not suitable candidates for an immunogen design. The results are in concordance with findings from South Africa, wherein highly cross-recognized peptides were from the p24 region (30). A significant positive correlation between magnitude and breadth of immune response clearly states the fact that a healthy immune system (most of the study participants were asymptomatic for any opportunistic infection at the time of sample collection) is able to mount a good immune response and has the ability to recognize more antigenic peptides compared to symptomatic and chronic individuals. The findings are in line with Gray et al. study where they also reported nonsignificant association of breadth and magnitude of IFN-γ responses with viral load, indicating that specificity or quality of the response would be crucial for viral control (13).
For the design of a successful global HIV vaccine, it is imperative that only those immunogens should be considered where both viral as well as population diversity are addressed. In summary, this study found limited cross-reactivity of HIV-1-infected Indian population against multiple variants of Gag PTE peptides, representing different subtypes and circulating strains of HIV-1 worldwide. Hence, further refinement of immunogens based on consensus sequences, taking into account human diversity, is required for the development of a globally relevant vaccine.
Footnotes
Acknowledgments
We would like to acknowledge Prof. Clive Gray and Lycias Zembe for sending us the PTE peptide sets and also for their kind help in data analysis. We thank Department of Science and Technology for providing the research infrastructure and financial support.
Author Disclosure Statement
No competing financial interests exist.