
Abstract
Hepatitis B and C infections can be either acute or chronic. The chronic infection can culminate in liver cirrhosis and hepatocellular carcinoma. Influence of the host genetic makeup on conversion of acute to chronic infection, development of cirrhosis, and hepatocellular carcinoma is an interesting area of research. Variability in different immune system genes may account for such differences in the outcome of infection. This article discusses single nucleotide polymorphisms in different host immunomodulator genes that have been frequently reported to influence the outcome of infection and severity of disease. The genetic variability could be utilized for the prediction of disease outcome and treatment responses.
Introduction
V
Two hundred forty million people are chronically infected with HBV and it claims 780,000 lives annually. In contrast, 150 million people are chronically infected with HCV and more than 500,000 people die due to complications of hepatitis C, each year (66). However, not everyone who contracts HBV or HCV gets chronic infection. Rather, 90% of HBV and 10% of the HCV-infected individuals resolve acute infection spontaneously during initial 6 months (92). About 15–30% of the chronic hepatitis patients go on to develop cirrhosis over the period of 20 years (47,92,117). Development of cirrhosis during chronic hepatitis predisposes the patients to hepatocellular carcinoma (85). Aforementioned findings raise some intriguing questions such as, which immunogenetic factors of the host are responsible for the spontaneous clearance of the infection or progression toward cirrhosis and hepatocellular carcinoma.
The objective of this article is to highlight and elaborate immunogenetic factors associated with spontaneous recovery or exacerbation of the infection. This article reviews our current understanding about polymorphisms in different genes encoding immunomodulators and their impact on the efficiency of immune system to deal with the HBV and HCV infection.
Mannose Binding Lectin
Mannose binding lectin (MBL) is an initiator component of the complement system through lectin pathway. This protein recognizes mannose residues on the surface of pathogens and facilitates their phagocytosis. The deficiency of MBL is very common and predisposes an individual to different infectious diseases (35,111).
This protein is encoded by mbl2 gene located on the q arm of chromosome 10 at position 11.2. The serum levels of MBL are strongly dependent on a single nucleotide polymorphism (SNP) at position −221 (allele X or Y) of the promoter region. Allele Y is associated with high, while allele X is associated with low expression of the gene (15). Three SNPs in the first exon of this gene are also correlated with low MBL serum levels. First, SNP occurs at codon 52 (allele D), second at codon 54 (allele B), and the third at codon 57 (allele C). Wild-type allele is referred as A, which is associated with normal MBL serum levels. These alleles form five haplotypes because of linkage disequilibrium. These haplotypes are YA, XA, YB, YC, and YD. High and low expression promoters with wild-type structural allele are represented by YA and XA, respectively. In contrast, YB and YD are mutant haplotypes linked with minimal detectable MBL serum levels (12). Haplotype YD is less common compared to haplotype YB, whereas haplotype YC is rare.
Envelope proteins S, L, and M of HBV and E1 and E2 of HCV are glycosylated, which are recognized by MBL (96,125). It is well known that low serum levels of MBL are associated with development of chronic hepatitis B leading to cirrhosis and hepatocellular carcinoma. Low serum levels of MBL have dose-dependent relationship with cirrhosis and hepatocellular carcinoma. Individuals who suffer from chronic hepatitis B and its complications, that is, cirrhosis and hepatocellular carcinoma have low expression promoter (X) and/or mutant allele (B) (12,15). Similarly, low serum levels of MBL are also associated with hepatitis C progression and poor response to antiviral treatment. Nevertheless, correlation between hepatitis C and MBL is not as clear as its correlation with hepatitis B (67,96).
Type I Interferon Receptor 1
The interferon type 1 (IFN-α/β) keeps check on viral replication and activates host immunity for clearing viral infection in acute stage (34). The signaling of IFN occurs through IFN receptors called IFN-1 and IFN-2. The polymorphism of different loci at the promoter and coding regions of interferon receptor-1 (IFNR1) is associated with susceptibility and protection from chronic hepatitis B and its advance stages. Polymorphisms at the promoter region (−568, −408, −77, and −3) and in coding regions (C/G at intron 3 and C/G in exon 4) can lead to susceptibility or protection from developing chronic HBV infection or its advance stages (38,53,128,129). The G and C at promoter positions −568 and −408 of IFNR1, respectively, and their related haplotype I were less susceptible to chronic HBC infection, while the C and T, at positions −568 and 408 and their related haplotype III, were associated with higher risk to chronic HBV infection (128).
However, there are no evidences of relationship between the disease outcome and severity of HCV with polymorphism of IFNR1. In addition, mutations in other genes (IFNL3 and IFNL4) affect the expression of IFNR1 during HCV infection and may lead to severe HCV disease and poor response to interferon therapy (10).
Human Leukocyte Antigen Complex
Human leukocyte antigen (HLA) complex is the oldest factor known to influence the outcome of HBV and HCV infection. It is human version of the major histocompatibility complex (MHC). Genes of the MHC are located close together on the short arm of chromosome 6. These genes are divided into three main classes: MHC class I, MHC class II, and MHC class III. Humans have three main MHC class I genes, known as HLA-A, HLA-B, and HLA-C, and six main MHC class II genes, known as HLA-DPA1, HLA-DPB1, HLA-DQA1, HLA-DQB1, HLA-DRA, and HLA-DRB1. Functions of MHC class III genes are very different from class I and class II, except they are located on the same region of chromosome.
Several hundred variants (alleles) of these genes are known and many different alleles can occur even within the same ethnic group. Therefore, studies from different regions of the world have different findings. Nevertheless, there are some alleles which are widely attributed to the outcome of HBV and HCV infection. Alleles DQB1*0301 and DRB1*1101 are well known to confer protection against chronic hepatitis C, DQB1*0301 being more effective in Africans (43,72,73,99,121). Similarly, alleles DRB1*0101 and DQB1*0501 were found to be protective in an Irish cohort infected with HCV through contaminated anti-D immunoglobulin (4,6,69).
The susceptibility to HCV chronicity and cirrhosis is widely attributed to allele DRB1*0701 among Caucasians (17,27,59,109). Similarly, the risk of hepatocellular carcinoma during chronic HBV infection is associated with two independent loci (rs7574865 at STAT4 and rs9275319 at HLA-DQ) in Chinese population (49). Another study in Chinese population reported two new loci (rs3130542 and rs4821116) associated with chronic HBV infection (44). However, association between HLA class I alleles and the outcome of hepatitis C infection is not as strong (84). Conversely, in different studies, different alleles are found to be associated with viral clearance or persistence (54,57,70,86,105).
Surprisingly, the development of chronic hepatitis B infection is widely attributed to DQB1*0301, the same allele that confers protection against chronic hepatitis C (50,104). Alleles DRB1*1301 and DRB1*1302 are known to confer protection against chronic hepatitis B among Europeans (18,42); however, the protective alleles among Chinese population are DQA1*0301 and DQA1*0102 (62). HLA-DP polymorphisms rs3077 and rs9277535 are found to eradicate infection in some Asian populations (127). However, there is a study which associated HLA-DPA1*0103-DPB1*0402 and HLA-DPA1*0103-DPB1*0401 with protection and HLA-DPA1*0202-DPB1*0501 and HLA-DPA1*0202-DPB1*0301 with susceptibility to chronic HBV infection, among Asians (52). Again, the correlation between HLA class I molecules and outcome of hepatitis B is weak.
HLA molecules encoded from the aforementioned protective alleles could be better candidates for presenting the suitable epitopes to T cells and, thus, drawing a healthy T cell response. These protective HLA molecules could have stronger affinity for suitable epitopes, which can lead to an effective immune response (19,72). Alternatively, some of these protective genes may be in linkage disequilibrium with other genes that promote viral elimination.
Tumor Necrosis Factor Alpha
The polymorphisms in the gene encoding tumor necrosis factor alpha (TNFα) may influence the development of chronic HBV and HCV infection leading to cirrhosis and hepatocellular carcinoma. This pro-inflammatory cytokine is produced by macrophages, monocytes, neutrophils, T cells, and natural killer cells in response to the inflammation, infection, and environmental stress. TNFα stimulates the secretion of other cytokines and expression of adhesion molecules. It also activates the neutrophils and plays a very crucial role in cytolytic and noncytolytic clearance of viral infections.
The TNFα gene belongs to the MHC class III and located on the p arm of chromosome 6 at position 21.3. The amount of cytokine production is significantly influenced by SNPs in the promoter region. So far, seven biallelic polymorphisms have been studied in TNFα gene which are as follows: −163G/A, −238G/A, −308G/A, −376G/A, −857C/T, −863C/A, and −1031T/C (40,51). In general, the polymorphisms that increase the transcription level of TNFα are considered as beneficial for clearance of infection, but harmful for the development of cirrhosis during chronic infections.
In a large number of studies, the presence of high expression −308A and −857T alleles and the absence of low expression −863A allele are found to be significantly associated with the resolution of HBV infection (22,55,90). It was reported in an in vitro study that TNFα accelerates the destruction of HBV mRNA and inhibits its replication (58). An in vivo study further found that TNFα destabilizes nucleocapsid of HBV and reduces the levels of covalently closed circular DNA of HBV in the nucleus (89). So far, no association has been observed between the TNFα promoter polymorphisms and the clearance or persistence of HCV infection (5,16,37). However, the high expression −308A allele does contribute to the rapid development of hepatitis C-related cirrhosis and hepatocellular carcinoma (48,87,124).
Interleukin-10
Interleukin-10 (IL-10) is another factor that contributes to the outcome of HBV and HCV infection, disease prognosis, and response to antiviral therapy (14,23,114,123). IL-10 is an anti-inflammatory cytokine from the interferon family. This is produced mainly by monocytes and to a lesser extent by B and T lymphocytes, which inhibit the activation of macrophages and production of pro-inflammatory cytokines by other immune cells. It also inhibits the activation of type 1 helper T cells (Th1 cells) by discouraging the cell surface expression of MHC class II molecules on the surface of professional antigen-presenting cells.
IL-10 gene is located on q arm of chromosome 1 between positions 31 and 32. Five biallelic polymorphisms that affect the transcription levels occur within the promoter region of IL-10 gene. Three (−1082 G/A, −819 C/T, and −592 C/A) of these five polymorphic sites have been studied in detail (110). Three haplotypes, GCC, ACC, and ATA, are formed due to linkage disequilibrium. Haplotype GCC is associated with high production of IL-10, while haplotype ATA is associated with its low production (9,46). Cell-mediated immunity plays a crucial role in clearing the viral infection. High serum levels of IL-10 are harmful for host as IL-10 suppresses the T cell response. It has been reported in many studies that individuals with low expression promoter, ATA, are better at resolving the HBV and HCV infections (20,56,63,74). Individuals with haplotype ATA are also found better to respond to antiviral therapy (56,63,114,123).
It is worth mentioning in this study that IL-10, being an antifibrinogenic, prevents fibrosis. In two independent studies, IL-10 therapy was shown to improve liver histology with reduced liver fibrosis (79,80). In fact, it was found in other researches that individuals with haplotype ATA and ACC are more prone to cirrhosis, whereas individuals with high IL-10 producing haplotype GCC are less likely to develop cirrhosis (36,56).
However, there are a few studies with contradictory results. It was found in one research that carriers of −592A allele are more prone to chronic hepatitis B (14). Likewise, another research found −1082G allele to be more frequent in patients who recovered from HCV (61). One study in Chinese population described association of −592C and −819C in IL-10 promoter with HBV-related acute liver failure (119). Focusing on SNP instead of the complete haplotype is a major drawback of these reports, which are that they targeted only one. Furthermore, their findings are not even consistent with the well-established fact that low levels of IL-10 are necessary for clearance of a viral infection. A further research in diverse populations infected with HBV and HCV may shed more light on this issue.
Interleukin-18
Interleukin-18 (IL-18) is a pro-inflammatory cytokine produced by monocytes, macrophages, and dendritic cells. It induces the production of Th1 cells and interferon-γ. IL-18 gene is located at position 23.1on q arm of the chromosome 11. Two SNPs in the promoter region of IL-18 gene, C/A polymorphism at position −607 and G/C polymorphism at position −137, are well known to confer protection from chronic infection, cirrhosis, and hepatocellular carcinoma. Both these SNPs disrupt the transcription factor binding sites, resulting in low expression of IL-18 gene (31).
It has been found in various studies that carriers of low expression alleles (−607A and −137C) are more likely to resolve HBV/HCV infection or suffer from only the milder forms of the disease. Whereas individuals with high expression alleles (−607C and −137G) are at higher risk of developing chronic infection, cirrhosis, and hepatocellular carcinoma (2,11,64,103,126).
The molecular mechanism by which IL-18 causes persistence of infection and liver injury is not well understood. There is little evidence that lower levels of this cytokine contribute to clearance of viral infection. In a study, mice inoculated with influenza A virus demonstrated that IL-18 deficiency leads to rapid clearance of virus and more efficient activation of Th1 cells (112). As mentioned above, IL-18 is a pro-inflammatory cytokine; it causes the liver inflammation. It is well-established clinical fact that the expression of IL-18 gets upregulated during chronic HCV infection and causes severe liver injury (98,113). Thus, the individuals who are genetically predisposed to produce low levels of IL-18 (carriers of −607A and −137C allele) are at lower risk of cirrhosis if developed chronic infection. The role of IL-18 in cancer development is much controversial and under investigation. (82). Increased plasma levels of IL-18 have been reported in various cancers such as gastric, renal, and pancreatic carcinoma (8,95,108). Moreover, the chronic inflammation, in this particular context, is caused by IL-18, may itself culminate in carcinogenesis (28,65,97).
Interleukin-28
Interleukin-28 (IL-28) is a cytokine from the interferon family. It upregulates expression of different genes that check viral replication. IL28 gene is located on the q arm of chromosome 19 at position 13.13. IL-28 protein occurs in two isoforms, IL-28A and IL-28B. Two SNPs near the IL28B gene (rs8099917 T/G and rs12979860 C/T) are considered strong predictors of the response to PEGylated interferon and ribavirin therapy against hepatitis C (29,71,75,91,101,106). These SNPs also affect the clinical outcome of HCV infection (94,107). As reported by these studies, individuals with CC genotype for the rs12979860 SNP and TT genotype for the rs8099917 SNP are far more likely to resolve the HCV infection and respond to the antiviral therapy than individuals with CT or TT genotype for the rs12979860 SNP and TG or GG genotype for rs8099917. Since a vast number of studies report that there is strong correlation between IL28B gene and response to the therapy, it is recommended that patients should have their genotype checked before going for therapy.
In contrast, there is little evidence about correlation between protection from hepatitis B and IL-28. It was found in one research that low levels of IL-28 are associated with chronic hepatitis B and poor prognosis of the disease (60). Another study carried with murine hepatocyte cell lines revealed that IL-28 inhibits HBV replication as well (93).
C-C Chemokine Receptor Type 5
C-C chemokine receptor type 5 (CCR5) is a strong possible candidate for the outcome of HCV infection and the course of disease. CCR5 is primarily expressed on Th1 cells. It has also been found on monocytes, dendritic cells, microglial cells, and cytotoxic T cells. The CC chemokines MIP-1α, MIP-1β, and regulated on activation, normal T cell expressed and secreted are the natural ligands of CCR5. Not to mention Th1 immunity is crucial for clearance of HCV infection. In simpler words, CCR5 expressed on Th1 cells interacts with aforementioned cytokines leading to the differentiation and proliferation of Th1 cells.
The CCR5 gene is located at position 21.31on p arm of chromosome 3. A 32 bp mutation, called CCR5Δ32, is frequently found in CCR5 gene. This is a frame shift mutation leading to truncated protein that cannot be expressed on the cell surface. It has been reported that individuals homozygous for this mutation are more prone to develop chronic hepatitis C (76,78,118). In addition, another research disclosed that carriers of this mutation respond poorly to interferon α monotherapy (1). Interferons are known to mediate immune activation by partially upregulating the expression of CCR5 on CD4 and CD8 cells (120). T cells expressing CCR5 may cause immune-mediated hepatocyte damage during chronic hepatitis C (3), and therefore, in some studies it was found that patients of chronic hepatitis C with same mutation show less pronounced hepatic inflammation or fibrosis (32,39,116). However, there are a few studies in which no association was found between CCR5Δ32 mutation and outcome of HCV infection (33,88,115). There has been very little research on correlation between CCR5Δ32 mutation and outcome of HBV infection; one such study reports that CCR5Δ32 does confer susceptibility to chronic hepatitis B (100).
Other Immune Factors
The aforementioned immune factors are widely believed to affect the outcome of HBV and HCV infection and disease prognosis. However, there are a number of other immune factors whose correlation with outcome and severity of infection have not been studied extensively. For instance, the toll-like receptor 2 (TLR2) binds to the HCV core protein and nonstructural protein 3 (NS3). Certain SNPs and deletions in the TLR2 gene (cytogenetic location: 4q32) are correlated with hepatitis C associated hepatocellular carcinoma. These SNPs and deletions are believed to impair the ability of TLR2 to bind with HCV core protein and NS3 (13,81).
Likewise, the role of vitamin D receptor (VDR) in the outcome of HBV infection is also being investigated. Calcitriol, a derivative and the active form of vitamin D, is an important immune regulatory hormone. It activates monocytes, stimulates cell-mediated immunity, and suppresses lymphocyte proliferation, antibody production, and cytokine synthesis. Calcitriol mediates its function by binding to VDR that is expressed on the surface of monocytes and T and B lymphocytes. Four SNPs are known to occur in VDR gene (cytogenetic location: 12q13.11), and two of them are correlated with outcome of HBV infection (7,100). However, these studies are not comprehensive and do not explain how VDR SNPs affect its functioning.
The expression of monocyte chemotactic protein 1 (MCP-1), a small chemokine that recruits monocytes and dendritic cells to the site of inflammation, is upregulated in hepatic stellate cells during HCV infection. A SNP at position −2518 of MCP-1 (cytogenetic location: 17q11.2-q12) is correlated with outcome of HBV infection and HCV-related cirrhosis (77,83).
In addition to IL-10, IL-18, and IL-28, role of interleukin-6 (IL-6) in establishment of the infection and severity of the disease is also accomplished. IL-6 is secreted by macrophages, endothelial cells, and Th2 cells, which induce acute phase response and help B cells to proliferate and produce antibodies. Several SNPs in the promoter region of IL-6 gene (cytogenetic location: 7p21) are known, among them −174G/C polymorphism is found strongly associated with the outcome and severity of the infection. The high producer genotypes, −174G/G, are more frequent in people suffering from chronic infection, cirrhosis, and hepatocellular carcinoma (5,24 –26,30,122). The molecular mechanism by which elevated levels of this cytokine result in persistence of the infection is not well established. The polymorphism of interferon gamma inducible protein CXCL-10 from G to A at promoter region (−201) was also linked with progression to sever diseases in chronic HBV male carriers (21). The role of different SNPs and their association with the outcome of HBV/HCV disease is summarized in Table 1 and Figures 1 and 2.
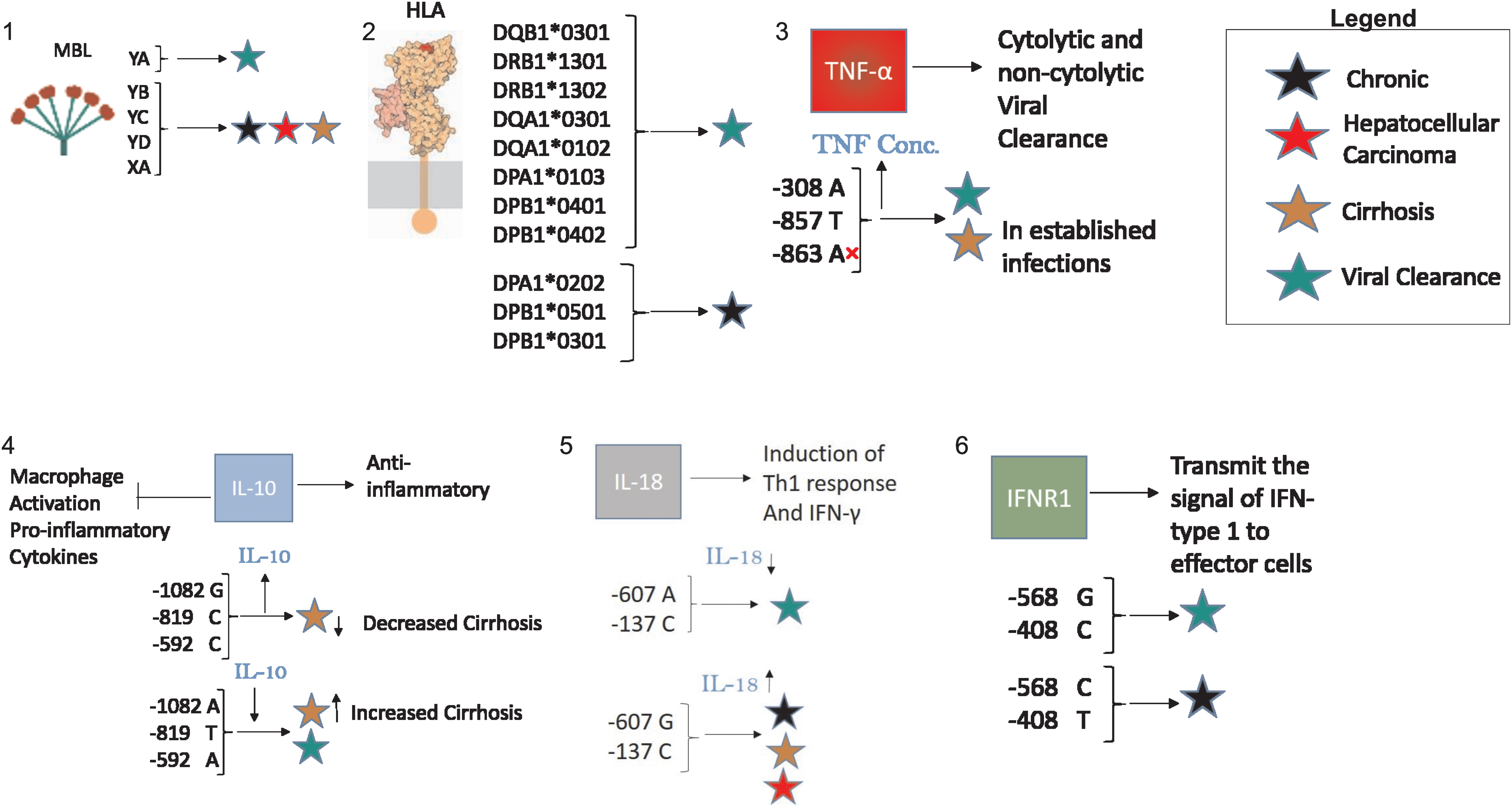
The summary of host immunomodulators' SNPs and their association with the disease outcome in HBV infection. The SNPs of different genes are depicted in numbers 1–6, (→) indicates the function of particular gene, (⊤) indicates the processes inhibited by particular gene, (↑) shows upregulation of particular gene, (↓) points downregulation of particular gene, and ( × ) demonstrates the absence of particular SNP. Details of the SNPs are given in the article text. HBV, hepatitis B virus; SNP, single nucleotide polymorphism. Color images available online at
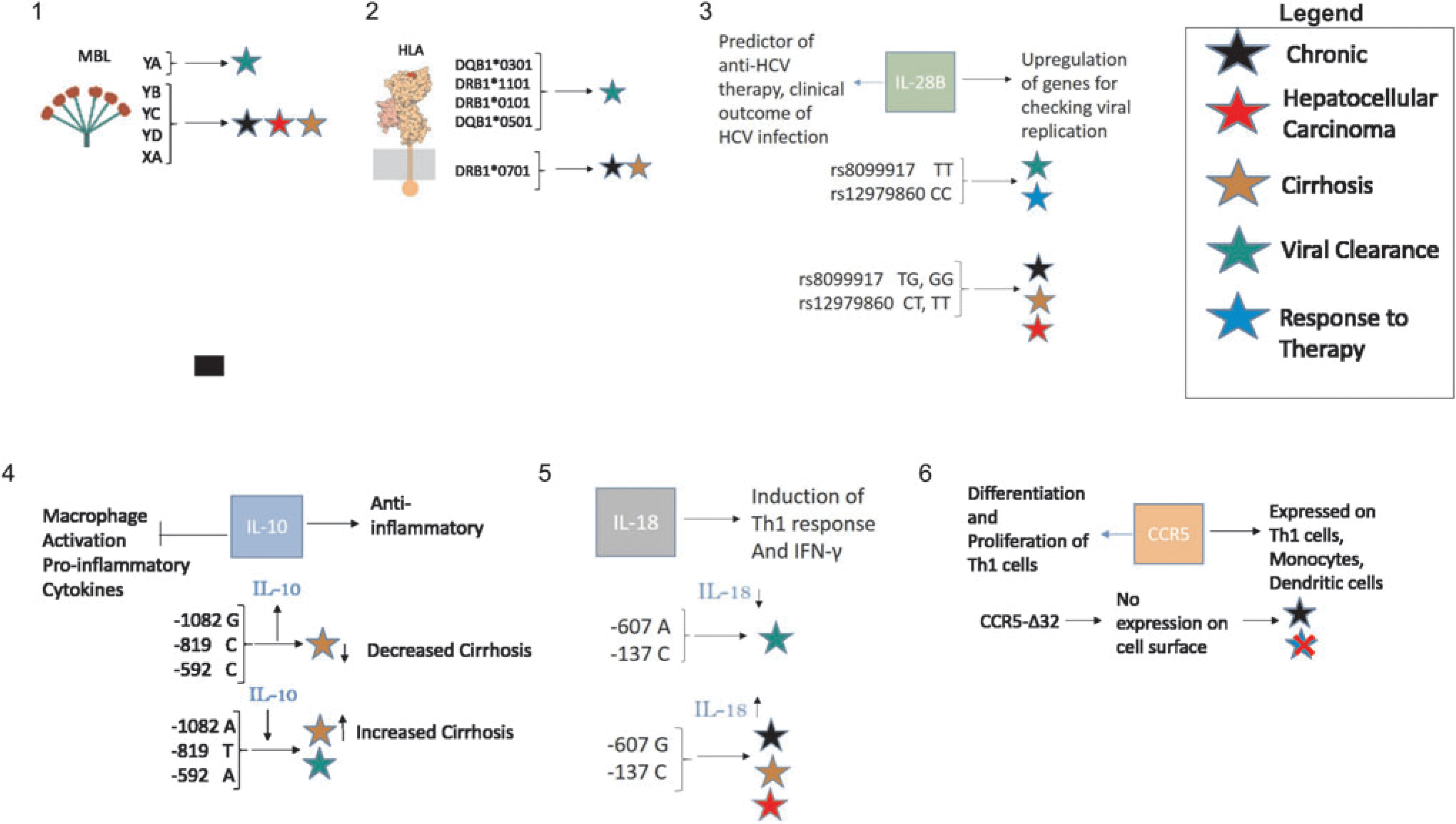
The summary of host immunomodulators' SNPs and their association with the disease outcome and success of interferon plus ribavirin therapy in HCV infection. The numbers 1–6 are explaining different genes, (→) indicates the function of particular gene, (⊤) indicates the processes inhibited by particular gene, (↑) indicates concentration increase of particular gene product, (↓) points concentration decrease of particular gene product, and ( × ) indicates the failure of interferon and ribavirin therapy. For detail of the SNPs, please read the text of the article. HCV, hepatitis C virus. Color images available online at
HBV, hepatitis B virus; HCV, hepatitis C virus; MBL, mannose binding lectin; SNP, single nucleotide polymorphism; HLA, human leukocyte antigen; IFNR1, interferon receptor-1; TNFα, tumor necrosis factor alpha; IL, interleukin; CCR5, C-C chemokine receptor type 5; RANTES, regulated on activation, normal T cell expressed and secreted.
Conclusion
It reviews our current knowledge regarding polymorphisms in the different gene encoding immune factors and correlation of these polymorphisms with the outcome and severity of the infection. Current studies focus on a SNP or a gene and, therefore, may be deficient to present an actual picture of polymorphisms and their influence on the disease prognosis. The allelic variants are carried by the long segments of DNA, which also contain other polymorphisms and inherited together in the form of haplotypes. The linkage disequilibrium forms complex patterns. Hence, thorough investigation is required to differentiate causative SNPs from those that are inherited merely because of linkage disequilibrium. Other modern techniques like microarrays and proteomics may help to further understand the actual role of immune factors in tackling the infection.
Several factors may account for inconsistencies regarding the association of SNP with disease severity. For instance, the studies were conducted in different races, populations, and ethnic groups. Different variants of aforementioned alleles are known to occur in different population and one population may have the preferential use of different alleles in clearance of infection (41,68). Racial heterogeneity may be considered for the future studies. Besides, genetic, environmental, viral, and even behavioral factors may play an important role in disease prognosis. Many studies involved a smaller sample size, which may decrease their power along with precision of the measurements of an association. Finally, reproducibility is another vital factor in this subject. To sum up, larger, well powered, and interdisciplinary studies involving epidemiology, immunology, virology, and molecular genetics may lead to more authentic findings. The advancement in the field will further unravel complex physiological and genetic pathways that may serve as potential drug targets, ultimately facilitating the development of more efficacious therapies.
Footnotes
Author Disclosure Statement
No competing financial interests exist.