
Abstract
The mucosal immune response against the porcine epidemic diarrhea virus (PEDV) is very important in piglets. To develop a PEDV vaccine suitable for inducing high levels of intestinal IgA in piglets, recombinant yeast expressing the PEDV S1 gene was constructed and tested by oral immunization of mice and piglets. The S1-specific IgG and IgA were tested at 0, 14, and 28 days postimmunization (dpi) in mice. Compared to the control group, the mice treated with S1 expressing yeast, demonstrated significantly higher levels of IgG and IgA against PEDV from 14 dpi onward. The recombinant yeast inducing a fecal IgA response in piglets was also tested. PEDV-specific IgA could be detected at 7 dpi and increased to 28 dpi. We demonstrated that whole recombinant yeast can be used as a PEDV vaccine vector for inducing high levels of IgA against PEDV in piglets. This could be a good vaccine candidate for PEDV control in piglets.
Introduction
P
PEDV possesses a 28 kb (approximately), positive-sense single-stranded RNA genome, which codes at least seven open reading frames (ORFs) (2). These ORFs code for four structural proteins: spike (S), envelope (E), membrane (M), and nucleocapsid (N); and three nonstructural proteins (replicases 1a and 1b, and ORF3). Among them, S, M, and N are related by inducing an immune response (11). One immunodominant region, S1, plays a significant role in infection and inducing an immune response (18).
PEDV initiates its infectious cycle at the mucosal surfaces, especially on the intestinal mucosal epithelial surface (9,10). To inhibit the entry of the virus into the body through the mucosal surfaces, an ideal PEDV vaccine would induce a high level of mucosal immune response (4,25). However, to date, both the commercial PEDV vaccine and the immune effect evaluation system (IgG test with enzyme-linked immunosorbent assay [ELISA]) are focused on an immune response at the systemic level. The development of a new vaccine, which induces high levels of mucosal immune response and the development of new mucosal immune response detection methods, may provide useful tools for PEDV control.
The yeast expression system is one of the widely used methods to produce recombinant proteins (22). Recently, some researchers have reported that animals immunized orally with whole recombinant yeast expressing target antigens could induce a high level of antigen-specific T cell immune response (17,26,28). A possible explanation for this is that yeast could activate the Dendritic cells (DC) cell by receptor-mediated conjugation (26). As DC cell plays an imported role in activation of systemic immune response, as well as local immune response, we designed a recombinant yeast expressing PEDV S1 in hope of inducing an active intestinal immune response against PEDV.
Materials and Methods
Vector construction
The Pichia pastoris codon-optimized full-length S1 gene of PEDV strain CV777 was synthesized by Shanghai Sunny Biotechnology Co., Ltd. The S1 gene was cutout of the PUC-57-S1 vector using BamH I and EcoR I, which had been sequenced into the 5′ and 3′ ends of the S1 gene. The yeast vector pPIC9K was purchased from Invitrogen and linearized with BamH I and Eco R I (the alpha factor signal sequence was removed). After ligation of the linearized vector and S1 gene, the recombinant vector pPIC9K-S1 was transformed into Escherichia coli DH5α. After identification by restriction endonuclease digestion, the recombinant vector pPIC9K-S1was sent for sequencing at the Shanghai Sunny Biotechnology Co., Ltd.
EGFP gene was amplified from the vector pEGFP-N1 and cloned into pPIC9K using the methods described above. The recombinant vector pPIC9K-EGFP was identified by sequencing.
Yeast transformation
The recombinant plasmids, pPIC9K-S1 (8 μL) and pPIC9K-EGFP (8 μL), were digested with Sal I, and the linearized vectors were separated using QIAquick Gel Extraction Kit. The competent P. pastoris GS115 cells were prepared according to the manual published by Invitrogen.
The linear plasmids pPIC9K-S1 (0.5–10 μg) and pPIC9K-EGFP (0.5–10 μg) were mixed with competent GS115 (80 μL, 30% W/V) cells on the ice. After 5 min, the plasmids were transformed into the competent GS115 cells using a Bio-Rad Gene Pulser, (1,500 V, 25 μF, and 200 Ω) then streaked and cultured on plates of minimal dextrose medium without histidine at 30°C for 3 days. These plates were screened for pPIC9K-S1 and pPIC9K-EGFP positive clones by polymerase chain reaction. The positive clones were cultured in 3 mL of buffered glycerol-complex medium (BMGY) overnight at 28°C.
Induction of EGFP and S1 expression
Fifty microliters of GS115 was inoculated into 50 mL BMGY and cultured at 28°C until the optical density at 600 nm (OD600) reached 4. The recombinant yeast cells were centrifuged and resuspended in methanol-complex medium at OD600 of 1. The cells were continuously cultured at 28°C in 5 mL/L methanol for 3 days. The methanol was supplemented at every 24 h. The expression of EGFP was observed under a fluorescence microscope and the S1 expression was determined using Western blot.
Ten milliliters of samples were collected at 24, 48, 72, and 96 h after the recombinant yeast cells were transferred into methanol-complex medium.
Western blotting
To examine S1 expression in yeast, whole yeast cell extracts were prepared by LiAc/NaOH extraction method using 10 mL of yeast culture (29). Protein samples were separated by sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) and transferred onto nitrocellulose membranes (Millipore). The membranes were prepared by incubating in PEDV-specific antibody (1:500) obtained from vaccinated and then infected piglets in our previous infection experiment (the titers were about 1:20,000 when analysis with commercial ELISA Kit [Biovet]) and then horseradish peroxidase (HRP)-conjugated goat anti-pig IgG antibody (ab112746; Abcam). To ensure equal loading of protein samples, Rabbit polyclonal anti-GADPH antibody (HA101; Tiangen) was used to reprobe the same membrane for S1 Western blotting. Signals were visualized using enhanced chemiluminescence (BoShiDe).
Mouse immunization and antibody detection
Immunization of mice
Immunogenicity of recombinant yeast expressing S1 protein was evaluated using 5-week-old BALB/c mice. Thirty mice were divided into three groups, with 10 individuals in each group. The first group of mice was given phosphate-buffered saline (PBS) and used as the negative control. The second group of mice was given same quantity of recombinant yeast expressing EGFP. The third group of mice was immunized with 0.5 mL of the recombinant yeast (in media OD600 = 1, 1 × 109 CFU/mL) expressing S1 protein in PBS buffer. Immunization was performed using a 1 mL syringe fitted with a gavage needle. After 2 weeks, five mice in each group were given booster immunizations using the same methods.
Serum IgG detection
Before the primary immunization, peripheral blood was collected by tail vein puncture bleeds and at 14 and 28 dpi, five mice in each group had taken blood by removal eyeball. We constructed pET-28a-S1 vector for expression histidine-tag fusing recombinant protein S1 in E. coli. The protein was purified with NI-column and used for detection antibodies against S1.
A concentration of 50 μg/mL purified recombinant PEDV S1 protein in 0.05 M NaHCO3 was used to coat ELISA plates. An ELISA was performed as described in Ref. (24). The coated plates were blocked with serum samples diluted to 1:100 with 1% skim milk in PBST buffer for 2 h at 37°C. The secondary antibody, HRP-conjugated goat anti mouse IgG (ab97265; Abcam), was diluted in PBST (1:5,000) and 100 μL per well was added. The plates were finally incubated with the chemiluminescent substrate, TMB peroxidase substrate solution (Pierce), for 10 min at 37°C. The reactions were terminated with 50 μL 2 M H2SO4 and the wells read at the optical density 450 nm.
Mucosal IgA
The method to detect IgA extracted from feces and diluted in saline was adapted from a previous report (13). Approximately, 0.1 g (wet weight) of the intestinal contents was obtained from decapitated mice at 14 and 28 dpi. The extraction buffer consisting of 0.01 M PBS pH 7.4, 0.5% Tween 20, and 0.05% sodium azide was then added to each tube at a ratio of 1 mL of buffer to 0.1 g of sample. The samples were thoroughly homogenized by manual shaking and mechanical homogenization with a vortex mixer. The suspensions were then centrifuged at 1,500 g for 20 min at 4°C.
Supernatant (0.2 mL) was transferred to a sterile microcentrifuge tube containing 2 μL of a protease inhibitor cocktail (Sigma). The tube was then vortexed briefly. The samples were centrifuged at 10,000 g for 10 min and the supernatants were used to detect IgA.
The mucosal IgA at 0, 14, and 28 dpi was tested using an S1 coated ELISA. The process was performed as that used for IgG detection, but the secondary antibody was HRP-conjugated goat anti-mice IgA (ab97235; Abcam). The results are expressed in terms of the OD450 of the extraction buffer (100 μL).
Immunization piglets and intestinal IgA detection
The experiment was conducted in a local industrialized pig farm. The offspring of three healthy young sows (3–4 parity) who had not been exposed to PEDV were used in this experiment. Thirty-one piglets were separated into three groups and used to conduct the immunization assay. Yeast culture media (2 mL; OD600 = 1) containing PBS (Group 1), yeast with GFP (Group 2), or yeast with S1 (Group 3) was given orally to the piglets soon after birth and on the day 2 with a 2 mL syringe. We conducted two times of vaccination in the aim of increasing yeast attachment on the gut mucosa and about 4 μg of the recombinant S1 was contained in the yeast.
Fecal samples (≥5) were collected in the morning at 3, 5, 7, and 14 dpi. Fecal IgA was extracted from 0.5 g feces using the same methods as used for extracting mice intestinal content IgA. IgA was tested using the same methods, with only changing second antibody to an HRP-conjugated goat anti-pig IgA (ab112746; Abcam).
The animal study protocols were approved by the Institutional Animal Care and Use Committee of the Shaanxi Province, and all experiments were conducted in accordance to the Guide for the Care and Use of Laboratory Animals.
Statistics
One-way ANOVA followed by a Tukey posttest was used for multiple comparisons. Statistical analysis was performed using SPSS 22 software. Differences were considered significant at p ≤ 0.05.
Results
Generation of recombinant yeast and analysis of antigen expression
The expression of recombinant protein S1was identified by Western blot. As shown in Figure 1B, the protein extraction from recombinant yeast expressing S1 reacted with the PEDV-specific serum and one band with molecular weight of about 59.2 KDa was seen in lane 3. No bands were observed in the lanes 1 and 2, which ran the protein extractions from yeast and yeast expressing EGFP, respectively.
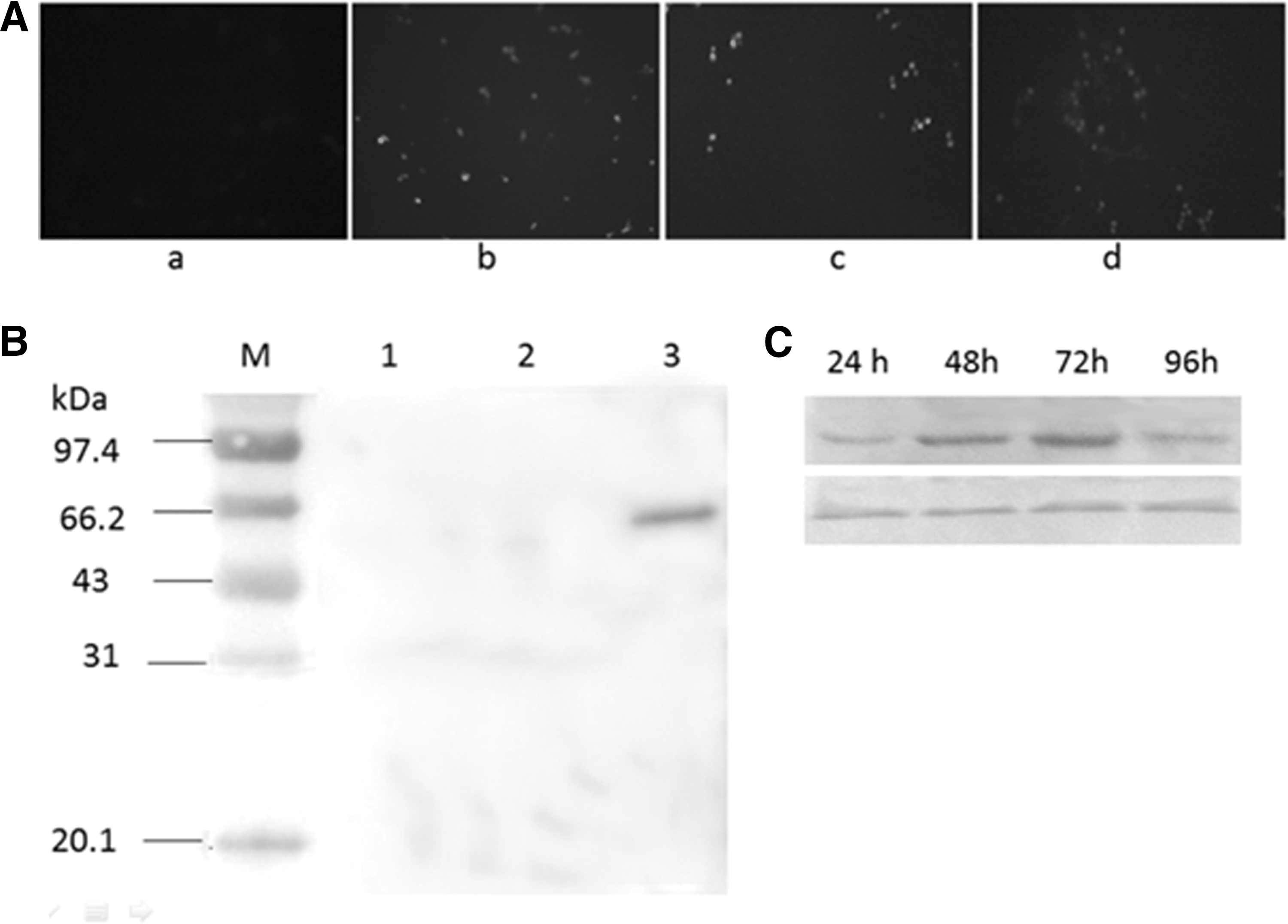
Identify S1 and EGFP expression.
The expression of EGFP was observed under florescent microscopy. As shown in Figure 1A, the maximum expression of EGFP was seen at 72 h, after which it reduced. The maximum expression of S1 was also seen at about 72 h (Fig. 1C), consistent with EGFP expression.
Antibody response of mice
To assess the immunogenicity of the recombinant yeast expressing S1 antigen protein, BALB/c mice were immunized orally with the whole yeast or recombinant yeast, which was induced with 5% methanol for 72 h. The chemicals (methanol and other chemicals in media) were removed by washing in ice cold PBS and recollected by centrifugation. Serum samples collected at different times were tested using ELISA. To reduce the effect of nonspecific reactions, sera samples were mixed with E. coli lysates at 4°C for 24 h before the S1-specific antibody testing. As shown in Figure 2A, the S1-specific IgG can be detected at 14 dpi and was significantly different (p < 0.05) to the control. After receiving the booster vaccine, the IgG immune response in mice was significantly increased (p < 0.01). No significant difference was seen in the control groups during the experiment.
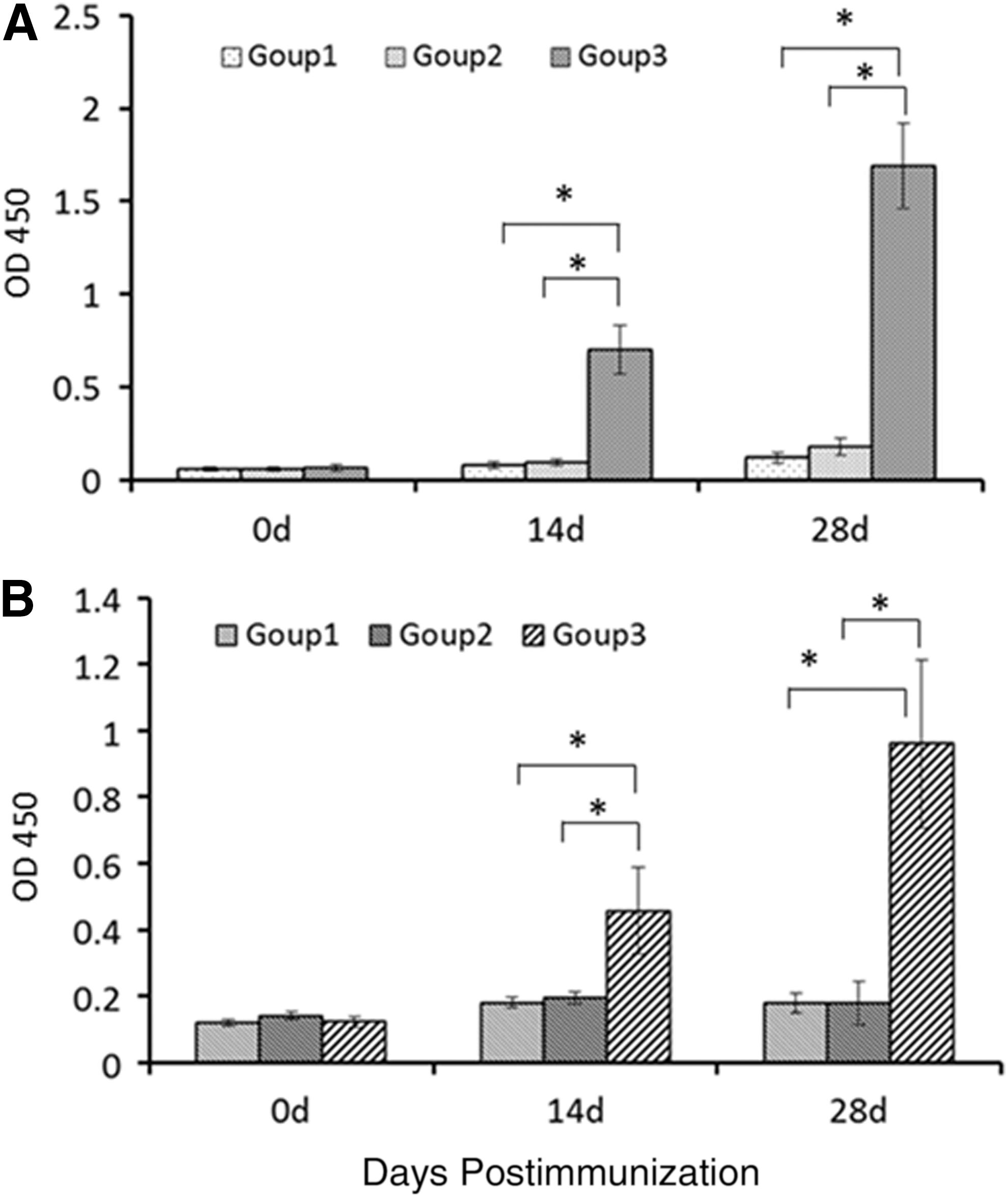
Mice systemic and mucosal immune response detection.
As IgA levels directly correlate with the immune protection of PEDV, anti-PEDV-specific IgA levels were determined in the intestinal contents of the mice. As shown in Figure 2B, PEDV-specific IgA was detected in mice fed with yeast expressing S1 protein after a booster immunization (p < 0.01). The IgA level of mice immunized with S1 was significantly higher than the control treated mice. During immunization, none of the mice died, and weight gain was the same across all groups (data not shown).
Piglets' IgA response to S1
A total of 78 fecal samples were collected from piglets and tested using ELISA. PEDV-specific IgA immune response was detected at 7 dpi in Group 3 and this response increased to 14 dpi. No PEDV-specific IgA was detected in the other two groups (Fig. 3).
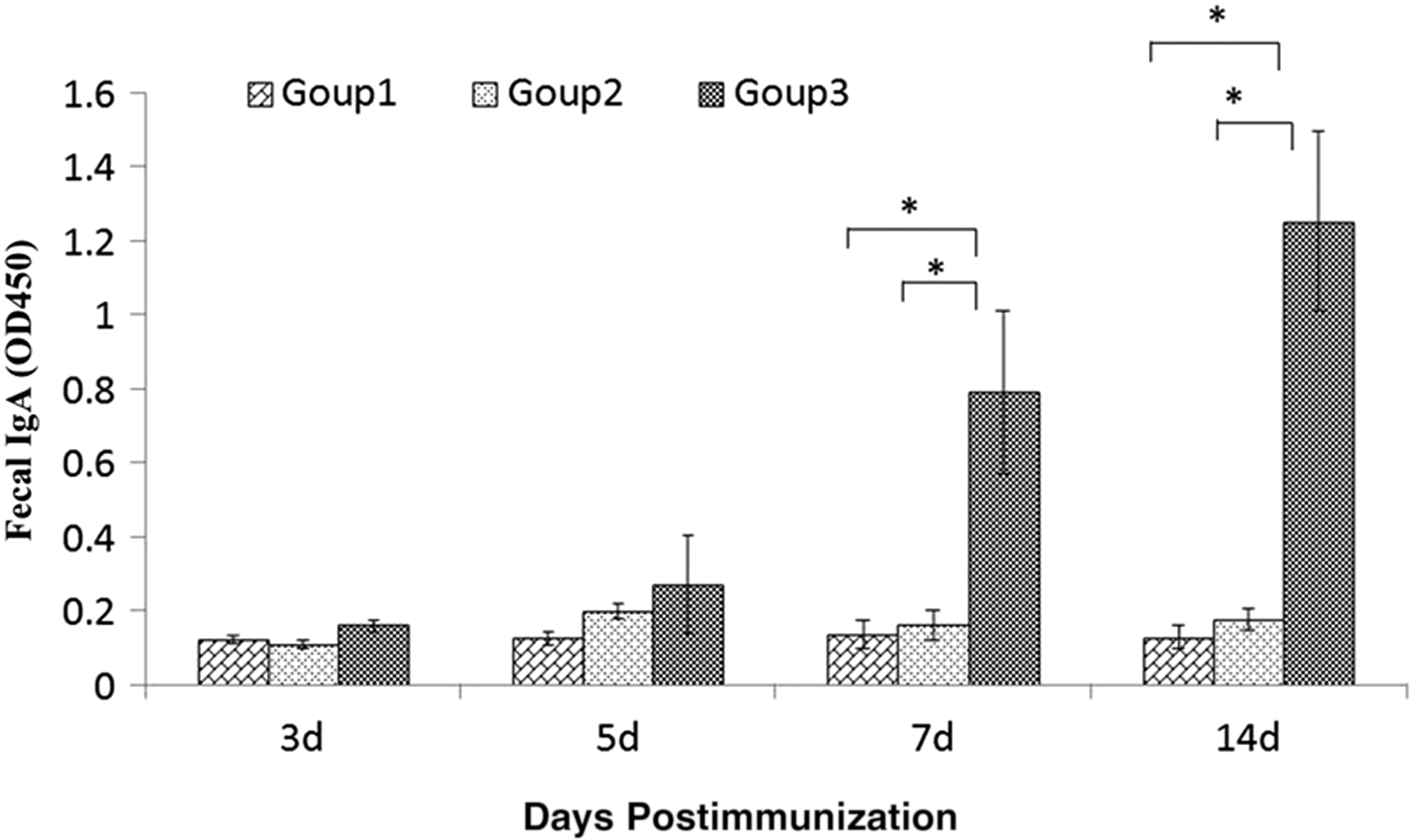
Piglets fecal IgA detection. A total of 78 fresh fecal samples were collected at 0, 5, 7, and 14 dpi and their extractions were diluted 20 times to test for IgA. OD450 values were used to measure the immune response. *Means significant.
During the experiment, no piglets developed diarrhea or other signs of alimentary disease. Only one or two piglets suffered a slight respiratory disease, but recovered within a day.
Discussion
The properties of the yeast P. pastoris make it an important tool for the expression of proteins (22). It is used widely for the production of pharmaceutical proteins (19). Different to E. coli, it does not have toxic cell wall pyrogens, and mammalian cells may contain oncogenic or viral DNA. Yeast can be grown to a high cell density using simple media and it can be manipulated genetically almost as readily as E. coli. Yeast is also suitable for high level production of soluble cytosolic proteins (22).
The yeast P. pastoris is widely used to produce vaccine antigens and several recombinant antigens have been commercialized (7,16,23). The immune studies have demonstrated good immunogenicity and immune protection using these recombinant antigens. Whole recombinant Saccharomyces cerevisiae yeast expressing tumor or HIV-1 antigens can potently induce antigen-specific cytotoxic T lymphocytes (CTL) responses, including those mediating tumor protections, in vaccinated animals (26). This report provided us with a new concept around the development of whole yeast vaccines. Several recombinant yeast systems expressing different antigens are reported and their protection effects have been confirmed by animal immunization (3,28,29).
Recombinant yeast represents a novel vaccine strategy for the induction of broad-based cellular immune responses. It exerts a strong adjuvant effect (28), augmenting DC presentation of exogenous whole-protein antigens to major histocompatibilty complex (MHC) class I- and class II-restricted T cells (26). Due to the unique way that the whole yeast vaccine is being used, it is developed into a new field of immunology: the intestinal mucosal immunology. The intestinal mucosal immunology is quite complex as the mucosal surface is not only a predominant structural and immunological barrier against the microorganisms but also plays an important role in absorbing water and nutrients during the digestive process (5). Our focus in these studies was the production of IgA, which is a key intestinal strategy to generate immune protection in a noninflammatory manner. DCs, as well as scattered M cells, extensively infiltrate the epithelium and capture antigens in the intestinal lumen (5,6,12). Following antigen uptake, these DCs may present the antigen to lamina propria CD4+ T cells, including antigen-specific Treg and Tr1 cells, which may thereafter initiate local IgA responses through the production of CD40 L and cytokines, including IL-10 and TGF-β (5,8,15,21). In our experiments, we demonstrated a high level of IgA immune response. As early as 14 dpi, a relatively high level of IgA in the intestinal content of the immunized group was detected. The immunity of the recombinant yeast in piglets was also confirmed. The PEDV-specific IgA was detected at 7 dpi and the levels increased throughout the experiment. Due to commercial constraints, we were not able to study the IgA levels at longer time intervals from immunization. However, our data did indicate that it is possible to develop a vaccine, which will induce an active immune response against PEDV in newborn piglets. To our knowledge, this is the first report to develop a newborn piglet PEDV vaccine.
Footnotes
Acknowledgments
The project was supported by the National Natural Science Foundation of China (Grant No. 31302122) and Programs for Science and Technology Shaanxi (2014K02-06-01).
Author Disclosure Statement
No competing financial interests exist.