
Abstract
There is a high demand for public immunization against Rotavirus (RV), especially in Africa. In Africa, the attenuated RV vaccination is contraindicated in patients with immune diseases and nutrition deficiency. Therefore, the inactivated RV vaccine (IRVV) could be an alternative. In this study, we aimed to develop a pentavalent-IRVV using the most circulating RV strains in Egypt and evaluate it against the commercially available Rotarix® vaccine. Trial-IRVV was developed with 5% sucrose, 2% polysorbate-80, and adsorbed on Alum to potentiate the vaccine immune response. Then, it was injected subcutaneously into mice groups at 0-, 21-, and 35-time intervals. In parallel, Rotarix was administered twice on 0 and 28th day. The success of the pentavalent-IRVV/monovalent-Rotarix vaccine immunity rested on achieving immunoglobulin G (IgG) exceeding 1:6,400 that implies less susceptibility to RV infection (RVI). IRVV stimulating IgG >1:6,400 could be an alternative vaccination approach to reach a reasonable protective immunization level against RVI. In addition, Alum adjuvant incorporation effectively provoked a triple elevation of the immunization pattern.
• Global Rotavirus (RV)-related deaths represent about 80% in low-income countries.
• Life-virus related events impede the global adoption of attenuated RV vaccines.
• Pentavalent, inactivated RV vaccine (IRVV) design using Egyptian circulating strains.
• Alum adjuvant was included as rational attempt for enhancing the immunogenicity.
• IRVV is alternative approach, creating a RV-infection reasonable immunization level.
Introduction
R
Globally and in Egypt, RV is the leading cause of severe gastroenteritis in infants and young children (6,25,26,47). The most common RV strains are G1P[8], G2P[4], G3P[8], and G4P[8] globally, which are responsible for about 88.5% of all RV diarrheal illnesses among children, besides the upcoming G9 strain (37). The Egyptian circulating RV strains are comparable to the globally known strains (1,6,10,25,32,44).
The natural RV infection (RVI) is partially protective against reinfections (9,42). Thus, vaccination can effectively prevent RVI and subsequent RV-related mortalities and morbidities. Currently, there are several attenuated RV vaccines available in the market; however, adoption of such vaccines in developing countries faces some challenges as limited immunogenicity in African developing countries (18,27,28,49) and relatively high expense in the African/Egyptian context (35,40). Moreover, they come with some serious side effects such as intussusception (24,4) and accidental reassortment events (15,16). Hence, new vaccination approaches are needed to bypass the existing limitations.
The inactivated vaccination approach is able to overcome such challenges (28). In addition, the inactivated RV vaccine (IRVV) showed sufficient immunogenicity in various animal models, and advanced immunogenicity was observed with the use of an adjuvant (13,20,21,23,45,48,50).
Consequently, we aimed to design a pentavalent IRVV containing the most common RV epitopes in the Egyptian environment and reveal the effect of the adjuvant (Alum) incorporation on the immunogenicity pattern. The IRVV and a commercially available attenuated RV vaccine (Rotarix®) humoral immunity were compared using a mice animal model. Immunoglobulin G (IgG) antibodies were estimated as an indicative proxy for appraising the IRVV/Rotarix immunity, where the RV-immunity/IgG level correlation was previously established by Velázquez et al. (43).
Materials and Methods
Stool sample collection
From November 2013 to February 2014, 18 stool samples were collected from hospitalized children <2 years with a severe gastroenteritis in the Pediatric department of El-Mattaria hospital, Cairo, Egypt. RV gastroenteritis typically reaches its peak in these months in Egypt (10). An oral consent was taken from children's primary caregiver before collecting stool samples.
RV antigen detection and genotyping
The RV detection and genotyping were performed according to the Naval Medical Research Unit-3 (NAMRU-3) protocols at their laboratories in Cairo, Egypt. Initially, a clear 20% stool-phosphate buffered saline (PBS) supernatant was exposed to RNA extraction using QIAGEN Commercial Kit (QIAamp Viral RNA Mini Kit, Qiagen®). The purified RNA was used as a template in OneStep reverse transcription polymerase chain reaction (RT-PCR) process to detect RV-VP6 using OneStep RT-PCR Commercial Kit (QIAGEN OneStep RT-PCR Kit). Samples with identified strains were subjected to VP7/VP4-genotyping process. In RV-identification and VP4/VP7 genotyping, the applied PCR programs and the selected primer sets were optimized as explained in NAMRU-3 protocols (Tables 1 and 2).
NAMRU-3, Naval Medical Research Unit-3; RT-PCR, reverse transcription polymerase chain reaction; RV, Rotavirus; VP, viral protein.
The VP6 band in positive-RV samples was estimated to appear at 380 base pair (bp). In the G genotyping, the G1, G2, G3, G4, and G9 bands were assessed to be detected at 158, 244, 466, 403, and 119 bp, on turn. In contrast, VP4 genotyping was projected to show the bands of P4, P8, and P6 at 483, 345, and 267 bp, respectively.
RV isolation and in vitro propagation
RV epitopes of G1, G2, G3, G9, and P[8] were selected for the bulk virus-lysate production due to their clinical importance in Egypt (1,6,10,25,32,44). Fecal RV strains of G1P[8], G2P[8], G3P[8], and G9P[8] were implicated for IRVV production. The virus isolation was performed using monkey kidney epithelial cells (MA104) and human colonic adenocarcinoma cells (Caco-2) provided by the Egyptian Holding Company for Biological Product and Vaccines labs-Egypt (VACSERA, Giza, Egypt). The isolation protocol includes two processes as follows: (i) initial RV isolation from stool specimen on MA104 cells (38,41); and (ii) RV titer elevation on Caco-2, due to the existence of cell integrin that correlated with RV adherence as α2β1, β1, and αXβ2 (17,30).
The tumor-origin cell lines as Caco-2 cells were currently appraised to be applied for vaccine trials only (7). The African green monkey kidney cell line (Vero) [CCL-81] from NAMRU-3 was selected for bulk RV strain propagation, as recommended by the World Health Organization for human vaccine production (46).
The media designed for maintenance of MA104, Caco-2, and Vero cell lines were DMEM, RPMI, and DMEM, respectively. The trypsin concentration for virus activation and cultivation was 20 and 1 μl/ml, respectively. In Caco-2 cells, a lower trypsin concentration of 0.5 μl/ml was used in the cultivation process. Virus activation and cultivation were performed at 37°C (38,41). Five consecutive passages of virus cultivation were performed on Caco-2 cells following the initial two runs on the MA104 cells. Then, 7–10 propagation passages were performed for bulk antigen production on Vero cells for each antigen pool.
Vaccine preparation and evaluation
Two bulk virus-lysates were produced after exposing maximally RV-infected cells to three cycles of freezing and thawing for virus releasing and harvesting, centrifugation of cell suspension for deposition of cell debris, and supernatant filtration. Bulk virus-lysates were: (i) the first virus-lysate mixture of G2P[8], G3P[8], and G9P[8] strains (multivalent-strain lysate) and (ii) the second virus-lysate of G1P[8] strain (monovalent-strain lysate). The final RV antigen pool was formulated by the admixture of the multivalent- and monovalent-strain lysates by 2:1 ratio, which was supplemented with 5% sucrose and 2% polysorbate 80 (Sigma-Aldrich Co.) as stabilizing agents and sterile filtrated using 0.2 μm Stericup filter (Millipore).
RV-lysate pool titer was estimated using plaque assay with Caco-2 cells (39), before the inactivation process. The virus strains were thermally inactivated in a water bath at 60°C for 2 h producing the IRVV, as previously described (13). To modify the IRVV immunological characteristics, sterile Alum (VACSERA) was added to yield a final concentration of 740.74 μg/ml. The inactivated vaccine-Alum mixture was kept overnight on stirrer at 4°C that resulted in antigen adsorption on Alum. Post inactivation, confirmatory tests were performed as verification of the virus-inactivation using tissue-culture technique and vaccine sterility by the direct inoculation method.
Immunization
To evaluate the immune response toward the thermally inactivated and formulated RV vaccine, 32 BALB/c mice (6- to10-week-old) of 16–18 g bodyweight were included. Mice were housed under healthy conditions at fixed temperature of 27°C and handled according to international guidelines in the Animal House of VACSERA. Mice were prepared and organized into separate groups before starting the immunization schedule, where a larger mice group was involved in IRVV-Alum trial. The designed mice groups were divided into: (i) reference positive-control group (Rotarix mice group; four mice); (ii) negative-control group (four mice); and (iii) IRVV group.
The negative-control mice group included two subgroups as follows: the first subgroup was vaccinated with propagation media (four mice); and the second subgroup was aimed for randomly negative sera collection from 10% of all nonvaccinated mice before starting the immunization schedule (four mice). The study-IRVV group was divided into two subgroups: the first subgroup was vaccinated with the IRVV without Alum (4 mice); and the second subgroup was vaccinated with IRVV adsorbed on Alum (20 mice). In all groups, mice were exposed to orbital sinus bleeding after at least 14 days of the given dose, so sufficient IgG antibodies could be developed.
The reference Rotarix group was vaccinated twice orally with minimum 4 weeks between the two doses (on 0 and 28th day), as recommended by the manufacturer, and blood was collected on the 27th and 42nd days. Three 0.5 mL doses of the IRVV/Alum-IRVV or propagation media were injected subcutaneously to the corresponding mice groups at 0-, 21-, and 35-time intervals, and blood was collected on the 49th day to measure the immunity results after the third dose (13). For evaluating Alum-IRVV immunity that results after the second dose, bleeding was done for only 20% of mice group on the 35th day. For IRVV/Alum-IRVV subgroups, extra two bleeding times on 56th and 70th days were designed to evaluate the immunity persistence (13). The immunization and bleeding schedule could be graphically illustrated in a simplified sketch (Supplementary Figs. S1–S5; Supplementary Data are available online at
Sera preparation and measurements
Sera were collected, exposed to heat inactivation at 56°C for 30 min, and stored at −20°C before being tested. The IgG-antibodies were determined as discussed before (11,50) with minor modifications. Enzyme-linked immunosorbent assay plates (Costar®) were coated overnight with 0.02 M ethylenediaminetetraacetic acid (EDTA)-lysate of G1 strain in 0.05 M carbonate buffer, nonreacting antigen removed by washing with PBS-0.05% Tween 20 buffer (PBS-T solution), and blocked for 1 h with 4% bovine serum albumin in PBS buffer at 37°C. The monovalent coating layer was representing an unchanging antigenic layer, where EDTA treatment aimed to detach the variable outer layer and expose the stable VP6 layer, so verifying the even reactivity. Washing was performed three times with PBS-T solution.
Incubation was performed with serum dilutions for 1 h at 37°C, where the starting serum concentration was 1:100 and followed by twofold dilution. A 1,000-fold diluted HRP-conjugated goat anti-mouse IgG was used for plate coating for another 1 h, and the resulted Ag-Ab complex was detected by TMB substrate at 450 nm followed by stopping the substrate-conjugate reactivity by 1 N HCL. The IgG level was estimated in duplicates for each sample.
Calculations and immunity appraisal
The optical density (OD) values, being greater than 2.1 times of the estimated negative sera, were considered positive, as previously described (50). The highest positive OD reading and its corresponding serum dilution reciprocal in the performed ELISA technique were tabulated among variant points and mice groups. For assessment of the pentavalent-IRVV immunity, the data were managed and analyzed using Microsoft Excel.
The averages of serum-dilutions reciprocal, which were considered as the highest positive, were plotted against the corresponding OD value average for the IRVV and IRVV-Alum system at 49, 56, and 70 time intervals. The relationship aimed to reveal the vaccine immunogenicity along the immunization intervals, besides simplifying the role of Alum-adjuvant in immunity. For the IRVV-Alum, the relationship between the serum-dilutions reciprocal and the corresponding OD values on 35th, 49th, and 56th days was plotted directly without subtracting the blank-OD value, where a direct link was approved to connect the OD value with the actual antibody titer, as established before (3).
Due to the antigenic content and vaccination dose difference, the comparative evaluation between the prepared pentavalent IRVV and the live-attenuated Rotarix vaccine was dependent on achieving IgG-antibody level >1:6,400 that implies less susceptibility of RVI (43).
Results
Strain detection and genotyping
The VP6 detection method indicated that all of the 18 collected samples were RV positive, except sample Nos. 5 and 18. The existed G/P epitopes in each RV-positive sample depend on the detected bands on the stained gel after electrophoresis. In G/P genotyping, the gel bands could reveal the present strains (Supplementary Figs. S1–S5).
Vaccine production and evaluation
The specimen no. 3, 12, 13, and 16 that had epitopes of high importance (G1, G2, G3, G9, and P[8]) were selected as an antigen source for the vaccine production. RV cultivation and propagation stages were successfully performed using the Caco-2 and Vero cell lines (Supplementary Figs. S1–S5). The ultimate virus-lysate titer was evaluated to be 2.1 × 107 PFU. Post heat-inactivation, the virus was approved to be nonreactive in cell culture. The vaccine was also successfully confirmed to be sterile.
Appraised immune response
The recorded data revealed the immune pattern postadministration of three doses of alum-adsorbed and nonadsorbed IRVV (Fig. 1), where an elevated immune response was obtained. The average of the highest positive-serum dilution was estimated to be about 1:1,600 and 1:22,400 followed by a sharp declining phase recording 1:3,700 in the IRVV group at 49, 65, and 70 time intervals, respectively. In the IRVV-Alum group, the highest positive-serum dilution average was 1:27,520, then continuously increased till it reached 1:67,200, followed by a declining phase recording 1:12,800 on the same intervals. Therefore, the Alum adjuvant effectively provoked a longer and triple IgG elevation. The OD curve on 35th, 49th, and 56th days of Alum-IRVV was progressively elevated (Fig. 2). The OD curve reflects the elevation of RV-specific IgG antibody titer by vaccination with the formulated pentavalent IRVV.
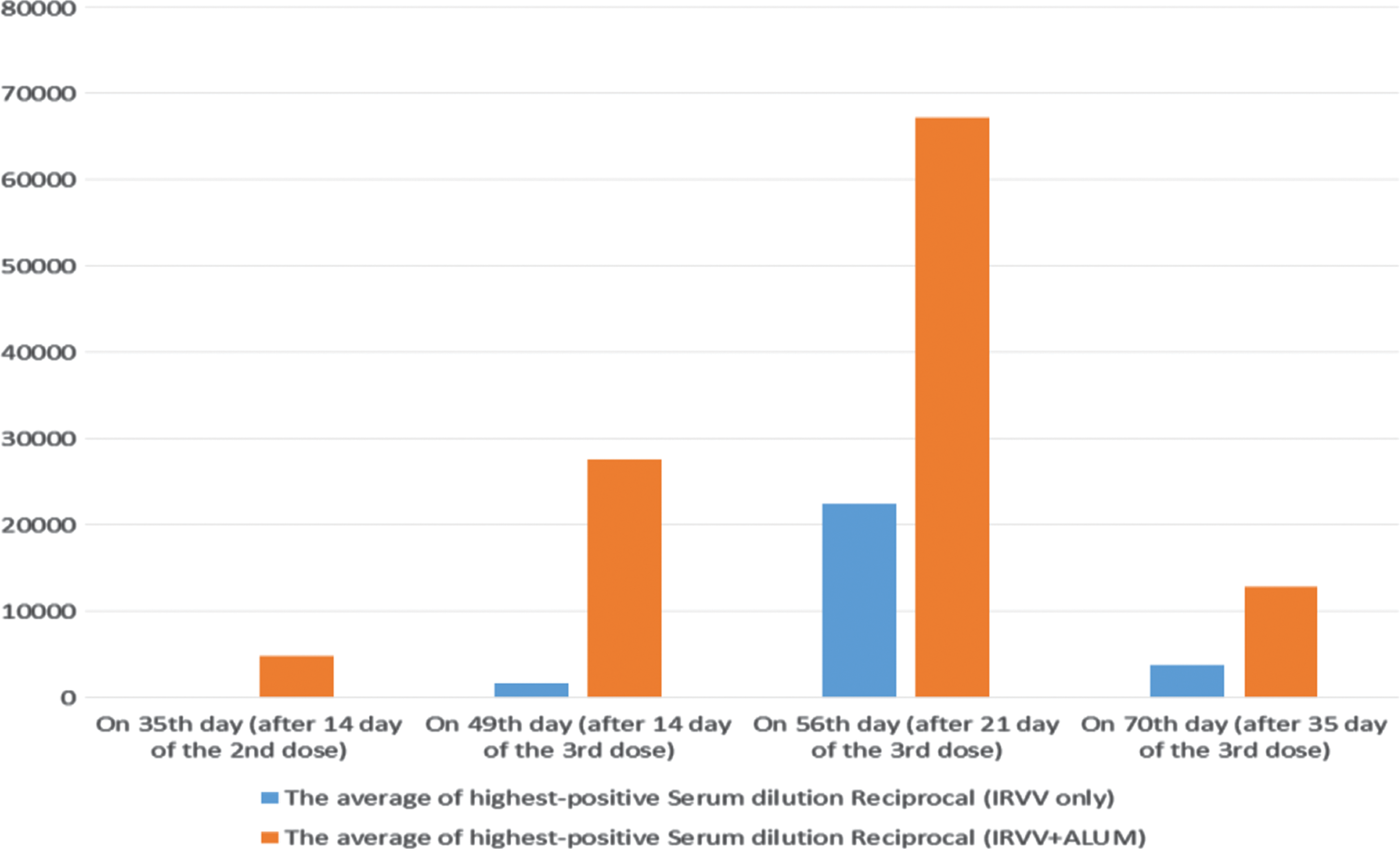
Average IgG antibodies from mice group vaccinated with IRVV (blue bars) and those vaccinated with IRVV-Alum (orange bars) after different time intervals of immunization. IgG, immunoglobulin G; IRVV, inactivated RV vaccine; RV, Rotavirus. Color images available online at
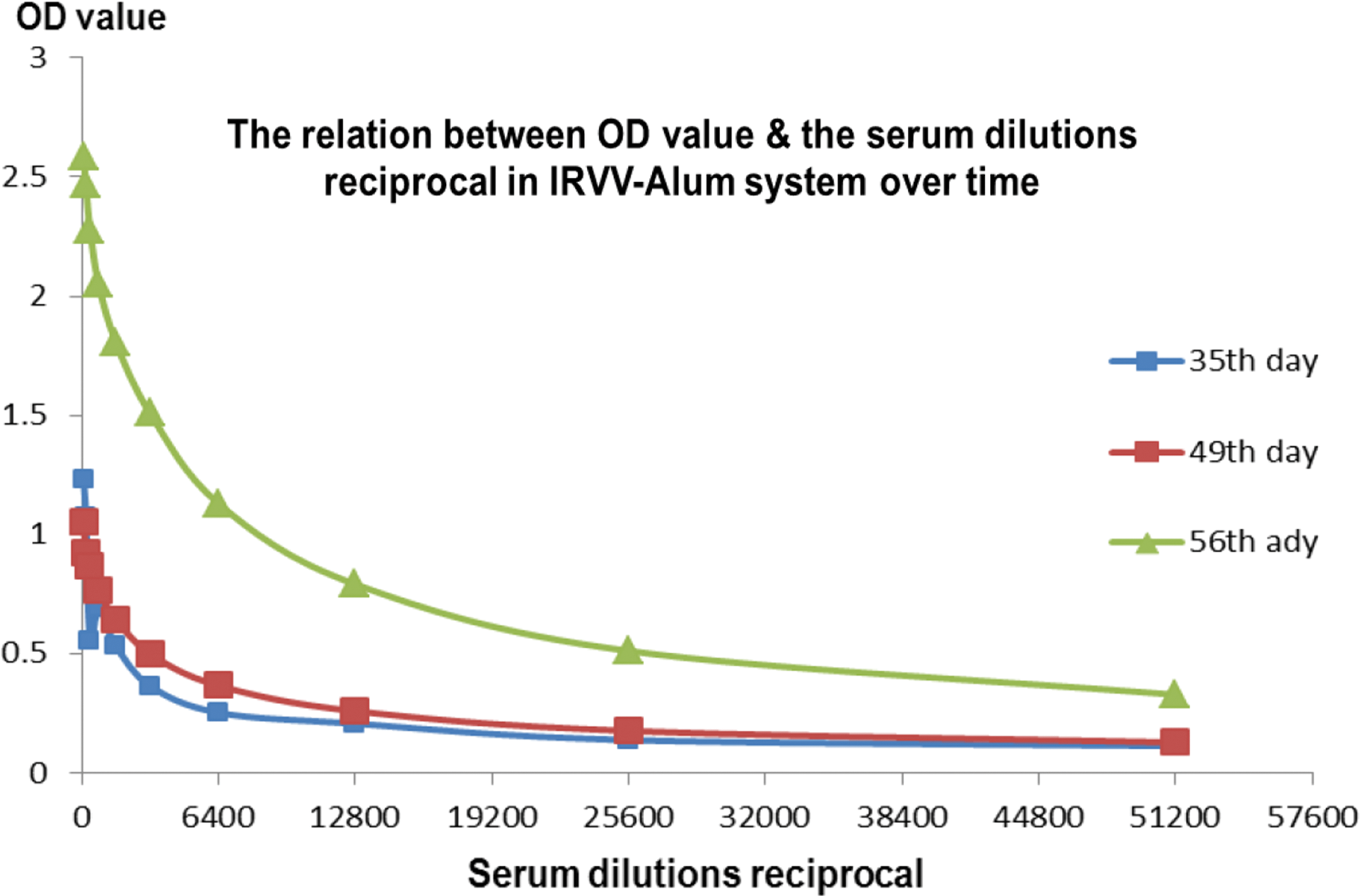
OD curves of IRVV-Alum vaccination on 35th, 49th, and 56th days. OD, optical density. Color images available online at
In Rotarix vaccinated group, the average of resulted IgG antibodies was initially 1:7,040 after the first vaccine dose and massively aroused after the second one. By comparing the pentavalent IRVV-Alum immunization data to the attenuated Rotarix vaccine, a comparable IgG level exceeding the protective value of 1:6,400 was established for both alternative vaccines.
Discussion
Despite the presence of commercial RV vaccines, some obstacles suppress their global adoption as the limited immunogenicity in poor countries (18,27,28,49) that might be attributed to the vaccine neutralization by exaggerated maternal antibody in serum and breast milk, severe malnutrition, and interference by flora and viruses in the gut (22,28). The Rotarix vaccine efficacy was documented to be 85%, 76.9%, and 49.4% in developed countries, South Africa, and Malawi, respectively (18,36). Moreover, the accidental vaccine/wild-type RV strain reassortment, which results in uncommon combinations as G1P[4], G2P[8], and G9P[4], was reported by the Australian Rotavirus Surveillance Program (15,16), besides the obstacle of high cost per vaccine dose (35,40) and the threatening intussusception risk (4,24).
These frailties, which are mostly related to the active virus presence, could be bypassed with virus inactivation. The IRVV (12) is capable of: (i) ensuring consistent efficacy among low/high-income countries; (ii) avoiding intussusception/reassortment events associated with oral vaccines; and (iii) providing less costly RV vaccination to be more suitable for routine/public immunization, especially in poor African countries. The RV vaccination cost-effectiveness could be duplicated by increasing the vaccine coverage among populations and reducing the vaccination price to $9 instead of $20 (40).
Therefore, the current study aimed to develop a promising pentavalent-IRVV from epidemiologically important RV epitopes using a mice animal model. As a result of difference in antigenic content and doses, the assessment of the trial-IRVV compared to the attenuated Rotarix vaccine was depended on yielding IgG level >1:6,400, representing less susceptibility to have a RVI (43). In the study, the IgG level was assessed to be 1:22,400 and 1:67,200 after 21 days of the third dose of the IRVV and IRVV-Alum formula, respectively. Accordingly, a satisfactory immunogenicity was achieved with both of IRVV and IRVV-Alum trials. However, the Alum incorporation produced a higher and long-lasting protection.
The adjuvant advanced immunogenicity could be attributed to the following: (i) the antigen presentation, delivery, and targeting distribution; (ii) potentiation/modulation of the antigen immunogenicity; and (iii) prevention of rapid antigen degradation and elimination. Therefore, the adjuvant incorporation would duplicate antibody titers, diminish the antigen content/dose, reduce the total number of doses, and diminish the immunization time. The Alum adjuvant was considered to act specifically by antigen slow-delivery, antigen-retention so more interaction with the immune system, macrophage/complement activation, and inflammatory factor induction (5,29,34).
Similar findings with regards to IRVV immunogenicity were observed in other studies (13,14,20,21,23,45,48,50). Skin, intramuscular, and subcutaneous IRVV mice immunization with various adjuvant systems as lipid adjuvants (MPL and L3) or Al(OH)3) was approved to be immunogenic (13,14,23,50). Gnotobiotic piglet's vaccination with three doses of IRVV was approved, similarly, to provoke a high IgG titer and protect against further RV shedding (45). Thus, there is enough evidence supporting the plausibility of the RV inactivated vaccination. However, large-scale research studies are warranted to explore the heterotopic and localized intestinal immunization that could be developed by the IRVV. In addition, there is a need to search for other alternative adjuvants that can provoke long-lasting immunization or modulate other immunity elements of the adaptive/innate system.
IRVV efficacy could be elevated by controlling the additives, especially the adjuvant system; immunization route; and virus epitopes involved in the vaccination. For example, the inactivated triple-layered virus particles were estimated to provide a higher immunization level than the double-layered particles due to the VP4 and VP7 epitopes present in the RV outer shell. Moreover, certain adjuvant incorporation was related for mucosal immunity induction, besides systemic immunogenicity (20,45). One of the adjuvant systems that should be extensively studied for immunization as a marvelous strategy is the mucosal adjuvants, including toxins, where the killed antigen could produce a reasonable immunity after oral inoculation. Mucosal IRVV could solve the main IRVV problem of lacking of the local immunity response (31), besides avoiding the killed antigen painful injection route.
Conclusions
We developed an IRVV that can be an alternative RV vaccination approach to reach a reasonable protective immunization level against RVI. In addition, Alum adjuvant incorporation effectively provoked a triple elevation for the immunization pattern.
Footnotes
Acknowledgments
The authors thank the Bacterial and Parasitic Disease Research Program laboratory at NAMRU-3, under direction of Dr. Hanan El-Mohammady for assistance in RV-strain identification and genotyping with the technical support of Dr. Rania Abdel-Khalek and Dr. Mireille Kamel. The authors thank Arbovirus Unit, Viral and Zoonotic Disease Research Program (VZDRP) at NAMRU-3 for providing Vero cells. The authors thank VACSERA laboratories for their support and thank Mahmoud Hashim (Researcher at Ingress-health, Germany) for statistical and grammatical editing.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.