
Abstract
In this study, we evaluated the immunogenicity and efficacy of mucosal delivery of a recombinant modified vaccinia Ankara virus (MVA) expressing the secreted version of bovine herpesvirus type 1 (BoHV-1) glycoprotein D (MVA-gDs) without addition of adjuvant in two animal models. First, we demonstrated the capability of MVA-gDs of inducing both local and systemic anti-gD humoral immune response after intranasal immunization of mice. Then, we confirmed that two doses of MVA-gDs administered intranasally to rabbits induced systemic anti-gD antibodies and conferred protection against BoHV-1 challenge. Our results show the potential of using MVA as a vector for the rational design of veterinary vaccines capable of inducing specific and protective immune responses both at local and systemic level.
Introduction
B
In countries with a high prevalence of this infection, vaccination is crucial to control the disease. Today, vaccination against BoHV-1 infection is performed with live attenuated or inactivated vaccines. The live attenuated vaccines are considered more effective, but can revert to a virulent strain and can also induce latency. In contrast, the inactivated vaccines are safe, but poorly immunogenic, and usually require repeated applications and coadministration of adjuvants. Furthermore, due to commercial restrictions of products derived from seropositive animals, the availability of vaccines that allow differentiation between infected and vaccinated animals has gained importance (36). Thus, the development of vaccines based on nonreplicative viral vectors could become a feasible alternative.
Modified vaccinia Ankara virus (MVA) has been extensively used as vector for producing human and veterinary vaccines because of its safety profile and its capability to induce durable and protective (humoral and cellular) immune responses. In addition, MVA vaccines are stable, can be lyophilized, and are suitable through different routes of immunization (9,26). Several researchers have evaluated MVA in clinical trials for HIV, tuberculosis, and malaria (16,19,22,44) and also as a potential veterinary vaccine against rabies, blue tongue, influenza, bovine tuberculosis, and bovine respiratory syncytial virus (1,4,5,6,47,48).
The local immunity at mucosal surfaces is known to be essential to control viral replication and spread. Indeed, secretory immunoglobulin (Ig) A is the predominant Ig isotype in mucosa and is a crucial effector by neutralizing pathogens present in mucosal surfaces (34,35). The upper respiratory tract is the main route of cattle infection by BoHV-1 and Israel et al. (23), in fact, demonstrated that antibodies in the respiratory mucosa are capable of inhibiting BoHV-1 replication.
The main target of the adaptive immune response against BoHV-1 and virus-infected cells is viral envelope glycoproteins B, C, and D (gB, gC, and gD). In particular, gD is essential for attachment and penetration of the virus into cells (25) and is also considered as an independent neutralization antigen (12,29).
Recently, several authors developed viral vectored vaccines expressing BoHV-1 gD and reported the induction of mucosal IgA after intranasal (IN) immunization. However, these vaccines induce poor levels of protection against BoHV-1 infection (27,28,37,50). Previous research has documented that IN inoculation of MVA elicits mucosal and systemic immune responses without generation of inflammatory reactions in the central nervous system or in the upper and lower airways (3,8,13,17,21,38).
In a previous study, we demonstrated that a recombinant MVA expressing a secreted version of gD from BoHV-1 (MVA-gDs) induces a potent and durable systemic anti-gD humoral response in rabbits when administered by the intramuscular route. In addition, this specific response confers protection against IN challenge with infectious BoHV-1 (15). Furthermore, IN vaccination with MVA-gDs plus cholera toxin (CT) as adjuvant induced a local immune response measured as anti-gD IgA antibodies in nasal and bronchopulmonary washes in a mouse model (15). Although CT is a strong mucosal adjuvant commonly used in research experiments, its toxicity has precluded its use for human trials and no information is available concerning veterinary vaccines with this toxin.
In this context, the aim of this work was to evaluate immunogenicity and efficacy of MVA-gDs administered by the mucosal delivery route without adjuvants.
Materials and Methods
Cells and viral stocks
Primary cultures of chicken embryo fibroblasts (CEFs) and Madin-Darby bovine kidney (MDBK) cells (American Type Culture Collection) were used as described before (15).
Viral stocks of MVA (wild type) and recombinant MVA-gDs (15) were propagated in CEFs, purified by centrifugation through a sucrose cushion and titrated as described elsewhere (14). The BoHV-1 strain Los Angeles (LA) was grown using MDBK cells at a low multiplicity of infection (10). BoHV-1 inactivation was performed with formaldehyde 0.2% for 24 h at 37°C.
Animal experiments and sampling procedure
All experiments were performed following internationally recognized guidelines with the approval of the Institutional Committee for Care and Use of Experimental Animals, CICUAE-CICVyA, INTA, Argentina. Mice and rabbits were housed in animal facilities at the Instituto de Biotecnologia, INTA.
IN immunization of mice
Specific pathogen free female BALB/c mice, which were 8–10 weeks old, were purchased from the Animal Services Laboratory, Faculty of Veterinary Medicine, University of La Plata. After 1 week of adaptation, the animals were slightly anesthetized with Isoflurane (2-chloro-2-(difluoromethoxy)-1,1,1-trifluoro-ethane) and vaccinated twice (day 0 and 21) by the IN route. Each dose consisted of 1 × 107 Plaque forming units (PFU) of purified MVA (n = 6) or MVA-gDs (n = 6) viruses in 50 μL TMN buffer (0.01 M Tris-HCl pH 7.5; 1.5 mM MgCl2; and 10 mM NaCl). Fourteen days after receiving the last immunization, mice were euthanized and bronchopulmonary washes were taken as described previously and kept at −20°C until used for enzyme linked immunosorbent assay (ELISA) tests (15). Serum samples were taken at days 1, 20, and 34 postvaccination and assayed for the presence of anti-gD antibodies by ELISA. Two independent experiments were performed.
Rabbit vaccination and challenge assay
Three-month-old New Zealand female rabbits were purchased from a breeding farm located in La Plata, Buenos Aires, Argentina. Three groups with four animals each were locally anesthetized with 2% xylocaine in spray and then immunized with MVA, MVA-gDs, or TMN buffer twice by the IN route. Each dose of viral vaccines contained 2 × 107 PFU of purified virus and was administered separately by 14 days. All groups were intranasally challenged with 1.2 × 107 TCID50 of BoHV-1 LA at 14 days post booster (dpb) using a protocol described previously (46). This method was selected because it mimics the natural route of BoHV-1 infection. Seventeen days post challenge (pch), the animals were euthanized and trigeminal ganglia were obtained to evaluate latency.
Serum samples were taken from all groups at different time points, and the presence of anti-gD antibodies was determined by ELISA. After the BoHV-1 challenge, nasal swabs were collected daily, processed individually, and inoculated in 10-fold serial dilutions over MDBK cells. Viral titers were calculated by the Reed and Muench method (40) and expressed as log TCID50/mL. Three blind passages in MDBK monolayers were performed with negative samples for BoHV-1 isolation.
Antibody measurement by ELISA
The presence of anti-gD antibodies in samples from vaccinated mice and rabbits was determined using a modified ELISA described previously (51). For the detection of serum antibodies, anti-mouse IgG + IgM peroxidase-conjugated (Axell) or anti-rabbit IgG peroxidase-conjugated (Sigma-Aldrich) antibodies were used. For the determination of IgA antibodies in nasal and bronchopulmonary washes from mice, anti-mouse IgA peroxidase-conjugated (Southern Biotechnology) antibody was used. The reaction was developed by adding 0.4 mg/mL 2-20-azino-bis-3-ethylbenzthiazoline-6-sulphonic acid (ICN Biochemicals) and 0.0015% H2O2 in 50 mM citric acid buffer (pH 5) and then reading at 405 nm in a Multiskan spectrophotometer (Labsystems). The optical density (OD) plotted corresponds to the mean OD value per group minus the background OD.
Statistical analysis
Statistical analysis was performed using GraphPad Prism version 5.00 for Windows. For mice experiments, we used Mann–Whitney U test (two tailed), Friedman test, and Dunn's multiple comparison test. One-way analysis of variance (ANOVA, repeat measures) with Tukey's Multiple Comparison Test and two-way ANOVA with Bonferroni posttest were carried out with rabbit data. p-Values <0.05 were considered significant.
Results
MVA-gDs administered intranasally induces both mucosal and systemic humoral response in mice
To evaluate the capability of MVA-based viral vector to induce local immune response without adding an adjuvant, we vaccinated mice with MVA or MVA-gDs by the IN route as described in the “Materials and Methods” section. Fourteen days after the booster, we assessed IgA antibodies (humoral mucosal response) in nasal and bronchopulmonary washes. Two doses of MVA-gDs administered by the IN route induced significantly higher levels of specific anti-gD IgA in respiratory mucosa than two doses of MVA (Fig. 1A, B). In addition, the IN immunization of two doses of MVA-gDs elicited a strong anti-gD systemic response (Fig. 1C).
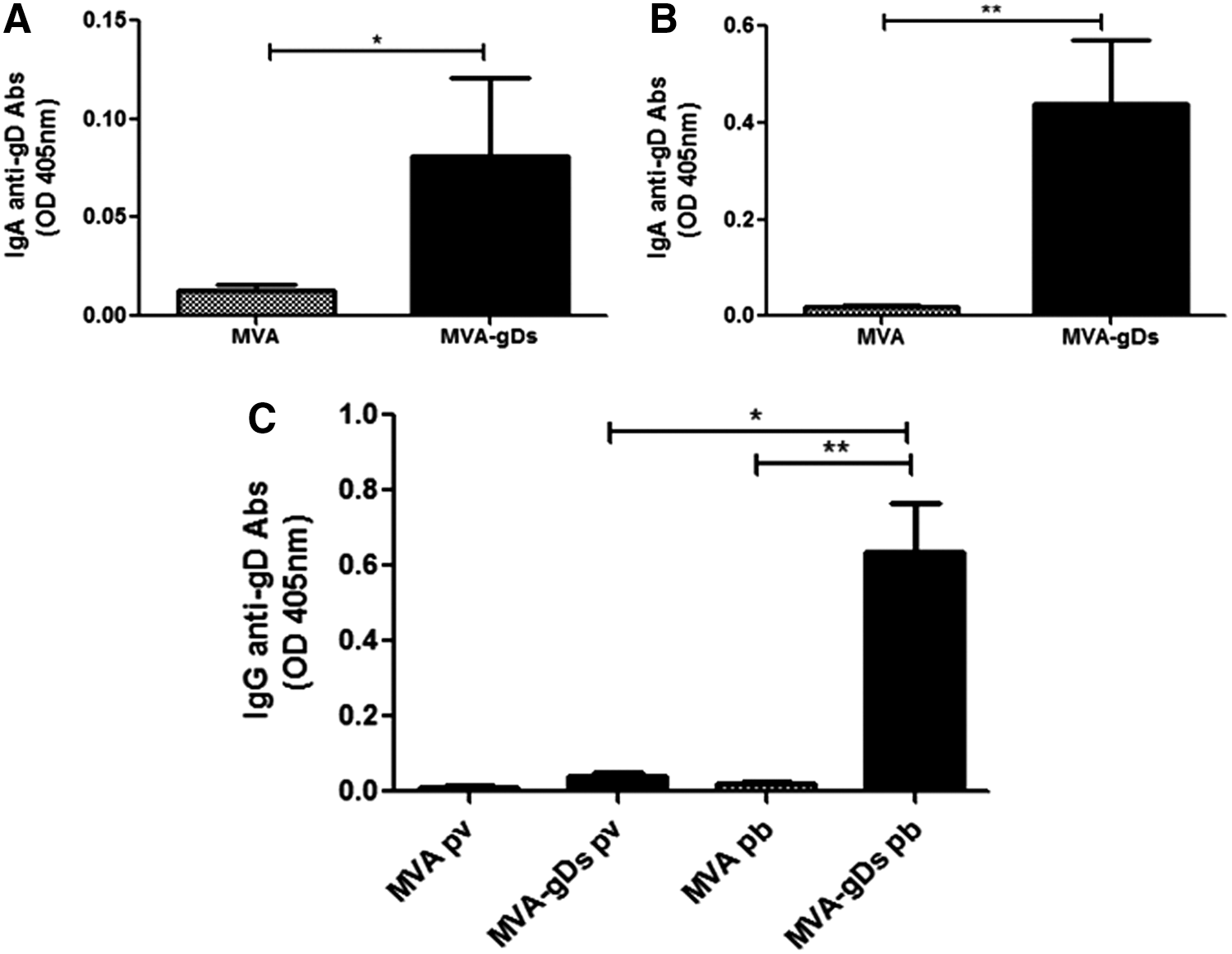
Mucosal and systemic anti-gD humoral responses induced by intranasal administration of MVA-gDs in mice. The Balb/C mice were immunized with two doses (days 0 and 21) of MVA or MVA-gDs. Nasal
IN administration of MVA-gDs induces partial protection against BoHV-1 in rabbits
The rabbit is a useful and well-established animal model to study both pathogenesis of BoHV-1 and the efficacy of experimental vaccines against this virus (15,41,43,46). After demonstrating that IN administration of MVA-gDs is capable of inducing specific mucosal and systemic immune responses in mice, we decided to evaluate the efficacy of the recombinant virus to protect rabbits against a BoHV-1 challenge. As described in “Materials and Methods” section, rabbits were IN vaccinated twice with MVA or MVA-gDs, and 14 dpb all the animals were IN inoculated with BoHV-1.
In the systemic anti-gD IgG response analysis, no significant differences were detected between wild type and recombinant MVA-based vaccines. However, the anti-gD antibodies in the MVA-gDs vaccinated group tended to rise after the booster (Fig. 2A) and a quick response was evident 17 days pch. These results demonstrate that vaccination was effective in priming the anti-gD response.
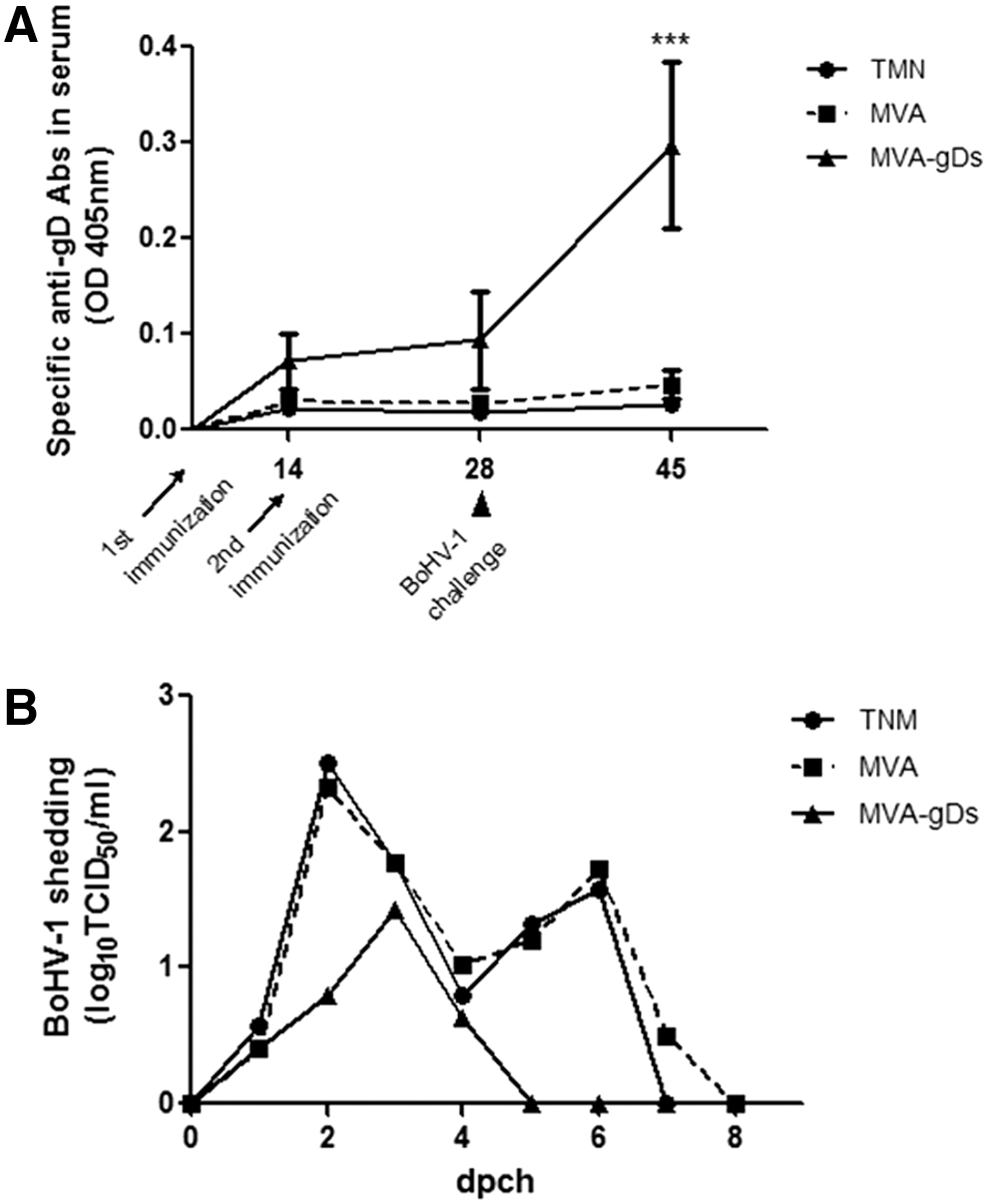
Evaluation of immunogenicity and efficacy of MVA-gDs in the rabbit model.
We finally assessed BoHV-1 excretion after challenge. Both TMN (negative control) and MVA vaccinated animals shed virus for 6 or 7 days (from day 1 to 7 pch) and reached the peak of excretion on day 2 pch with mean levels of 2.5 and 2.3 log TCID50/mL, respectively (Fig. 2B). No significant differences were detected between groups throughout the evaluated period. In contrast, the MVA-gDs vaccinated group presented a shorter period of shedding, with a maximum of 3 days, than the TMN and MVA groups. In addition, the peak of excretion was delayed 1 day (3 pch) and showed reduced titers (mean value of 1.4 log TCID50/mL). The MVA and MVA-gDs means were significantly different (p < 0.05).
Considering these results and taking into account that efficacy of vaccines against BoHV-1 is evaluated in terms of reduction of virus shedding after challenge and also by the duration of virus excretion (36), our data suggest that the MVA-gDs vaccinated group was partially protected against challenge.
Discussion
BoHV-1 infection is distributed worldwide, and the main strategy of control used in countries with a high prevalence of infection is based on vaccination. The licensed vaccines that are currently used to tackle this infection are live attenuated or inactivated; however, they present several disadvantages related to safety and efficacy, respectively. To overcome these problems, vaccine research is focused on the development of rationally designed vaccines based on recombinant vectors expressing immunogenic antigens also allowing the differentiation between vaccinated and infected animals.
MVA has several properties that make it an excellent candidate to develop human and veterinary vaccines (9). Indeed, MVA has an excellent safety profile because it is unable to productively replicate in mammals. In addition, it is genetically and physically stable and can be produced at large scale. Finally, MVA induces both humoral and cellular immune responses even without the coadministration of adjuvants.
Recovery from BoHV-1 relies mainly on cell-mediated immunity while humoral immunity prevents and controls reinfection (2). The antibody effector functions include virus neutralization, complement fixation, and antibody dependent cell-mediated cytotoxicity. BoHV-1 infects the host through the mucosal surfaces and protection from reinfection is correlated to levels of mucosal antibodies (23,37,52).
Several studies have reported that MVA-based vaccines administered intranasally are effective to induce a protective response against respiratory virus such as influenza (3), parainfluenza (13), and respiratory syncytial virus (49). However, our previous work (15) is the only one reporting the evaluation of a recombinant MVA as vaccine candidate against BoHV-1. This previous work reports a recombinant MVA expressing a secreted version of BoHV-1 gD, which was immunogenic (local and systemic) in mice by IN administration with CT and protective against BoHV-1 infection in rabbits vaccinated by the parenteral route.
In the present study, we assessed the immunogenicity and efficacy of the recombinant candidate vaccine MVA-gDs after IN inoculation without the coadministration of adjuvants in two animal models. We observed that the administration of two doses of MVA-gDs elicited similar magnitudes of anti-gD specific antibodies in serum and respiratory mucosa of vaccinated mice than MVA-gDs coadministered with CT (15). This result may be due to the immunostimulatory capacity of MVA (33). In addition, these results support previous studies in which a systemic immune response could be induced after IN immunization with MVA-based vaccines (17,26).
Even though antivector immunity could affect revaccination response with the same vectored vaccine, Draper et al. (11) reported that MVA antibody induction using homologous or heterologous immunization schemes is clearly antigen dependent. Our results demonstrated that a homologous prime-boost immunization with MVA-gDs increased (local and systemic) specific humoral immune response against BoHV-1 gD protein, thus indicating that antivector immunity did not interfere with anti-gD specific immune response (15).
Indeed, Gherardi et al. (17) observed that two doses of MVA-env administered intranasally induced higher levels of anti-gp160 serum antibodies than single immunization. Meseda et al. (31) and Breathnach et al. (5) also reported high titers of anti HSV-2 gD and anti-equine influenza virus antibodies in serum and nasal secretions in MVA homologous prime-boost immunization schemes. However, other authors documented better humoral responses with heterologous vaccination schemes using protein antigens, DNA vaccines, or adenovirus vectors as prime and recombinant MVA as boost (18,24,42).
Considering the immunogenicity induced by our immunogen after IN administration in mice, we decided to assess its efficacy in a BoHV-1 challenge test. Rabbits are an appropriate model since it mimics the pathogenesis of BoHV-1 infection in cattle. Indeed, after IN inoculation, rabbits shed virus for ∼5 to 7 days, develop typical respiratory clinical signs, and BoHV-1 is able to establish latency in trigeminal ganglia (7,30,32,46). Thus, to test MVA-gDs efficacy, we immunized rabbits intranasally twice and then challenged them with infectious BoHV-1 14 days post revaccination.
In agreement with our results, several studies reported that control animals show periods of viral excretion of 5–7 days after BoHV-1 challenge and the peak of excretion was on day 2 pch (15,32,43,46). Importantly, the vaccine candidate curtailed the period of viral excretion and the level of virus shedding compared with the control groups. However, in our study rabbits showed no respiratory clinical signs and we could not detect viral DNA in trigeminal ganglia (data not shown). These discrepancies with other authors could be due to the age of the animals and/or to the viral strain used in our study (30,32,43). In addition, after BoHV-1 challenge, the MVA-gDs vaccinated animals showed an increase in serum anti-gD IgG (anamnestic response), which suggests that animals had been primed for gD-specific antibody responses.
Current vaccines only partially protect against disease and not against infection; therefore, improved vaccines are required based on new immunogens, delivery, and formulations, which will protect against both infection and latency. Besides, since latently infected animals are considered reservoirs of the virus, new vaccines should also prevent the establishment of latency. Since we were unable to detect viral DNA in the trigeminal ganglia of the three vaccinated and challenged groups (data not shown), we could not confirm that vaccination with the recombinant vaccine impairs the establishment of latency after challenge.
Overall, our data suggest that IN administration of two doses of MVA-gDs conferred partial protection to rabbits against BoHV-1 challenge.
Several live vector vaccines have been evaluated as vaccine candidates against BoHV-1 in animal models and cattle. Bovine adenovirus 3 (28,39,50), human adenovirus 5 (20,37), and Newcastle disease virus (27) expressing gD elicited specific humoral immune response, but conferred low levels of protection against BoHV-1 infection in coincidence with our observations of immunogenicity and partial protection induced by MVA-gDs in the rabbit model.
BoHV-1 gD is the major target of neutralizing antibodies and cytotoxic T lymphocytes and is considered the principal antigen in the development of rationally designed vaccines. However, the immune response elicited by gD seems to be insufficient to fully protect animals after challenge. A feasible approach to improve vector immunogenicity could be the incorporation of sequences of other relevant BoHV-1 proteins such as gB and gC or cytokines. Another possibility could be the implementation of heterologous immunization schemes combining MVA-gDs with DNA or adenovirus-based vaccines.
Finally, this study shows the capability of MVA-gDs of inducing both specific (local and systemic) humoral response in mice and protection against BoHV-1 infection in a rabbit model.
Conclusion
In the last years, the rational design of recombinant vaccines that overcame the limitations of conventional vaccines has gained importance. MVA-based vaccines are considered as excellent candidates. In this study, we demonstrated that a recombinant MVA expressing a secreted form of bovine herpesvirus-1 gD administered intranasally without the addition of adjuvant is capable of inducing local and systemic humoral immune response in mice. In addition, we tested this candidate vaccine in the rabbit model and observed that mucosal delivery conferred protection against BoHV-1 challenge.
Footnotes
Acknowledgments
This work was supported by grant PNBIO 1131032 from the Instituto Nacional de Tecnología Agropecuaria (INTA). The authors are grateful to Danilo Buscafusco and Sebastian Di Giacomo for their help in rabbit's euthanasia and acknowledge Mr. S. Díaz for animal care and Mrs. M.J. Mónaco for technical assistance.
Author Disclosure Statement
No competing financial interests exist.