
Abstract
Human immunodeficiency virus type-1 (HIV-1) infection represents one of the biggest public health problems worldwide. The immune response, mainly the effector mechanisms mediated by CD8+ T cells, induces the selection of mutations that allows the virus to escape the immune control. These mutations are generally selected within CD8+ T cell epitopes restricted to human leukocyte antigen class I (HLA-I), leading to a decrease in the presentation and recognition of the epitope, decreasing the activation of CD8+ T cells. However, these mutations may also affect cellular processing of the peptide or recognition by the T cell receptor. Escape mutations often carry a negative impact in viral fitness that is partially or totally compensated by the selection of compensatory mutations. The selection of either escape mutations or compensatory mutations may negatively affect the course of the infection. In addition, these mutations are a major barrier for the development of new therapeutic strategies focused on the induction of specific CD8+ T cell responses.
Introduction
S
The activation of CD8+ T cells depends on the interaction of the T cell receptor (TCR) with a protein segment of the virus, which is presented through the human leukocyte antigen class I (HLA-I) (38). However, due to the high genetic variability of the virus and the strong selective pressure exerted by the CD8+ T cell response, viral populations with mutations in epitopes, affecting its presentation, can be selected in the host (10,73). Thus, the selection of these mutations by the immune response restricted to HLA-I leads to the accumulation of escape mutations in viral antigens, and represents a mechanism of immune evasion, facilitating the adaptation of the virus to host immune responses, which can be highly predictable based on the frequency of HLA-I alleles in a population (50). In this review, we describe the selection process of immune escape mutations, the main selective forces, and the viral evolution associated with CD8+ T cells restricted to HLA-I molecules. Furthermore, we expose the effects of these mutations in the viral replicative capacity, their importance in the clinical course of the infection, and in the development of new therapeutic strategies based on the induction of specific CD8+ T cell responses by vaccination, or in the elimination of the viral reservoirs.
Selection of Escape Mutations by CD8+ T Cells
One of the most striking features of HIV is its high genetic diversity. The high mutation rate of the viral reverse transcriptase, estimated at 2.4–3.4 × 10−5 mutations per site per replication cycle (56,72), along with an approximate recombination rate of 2.8 crossovers per replication cycle per genome (94), contribute to a large generation of viral quasispecies within hosts. However, all these diverse viral populations do not expand at the same rate in the patient, since a certain viral genotype dominates as the result of a positive selection due to phenotypic constraints expressed by the individual (27). In the case of HIV-1, the immune response, especially the CD8+ T cell responses, exerts a high selection pressure on viral populations. CD8+ T cells are crucial for the partial control of HIV replication in the acute phase, as it has been shown that the expansion of HIV-specific CD8+ T cells correlates with a decrease of the plasma viral load (47). These cells recognize infected cells through the interaction of the TCR with a protein segment of the virus presented in the context of an HLA class I molecule. It has been reported that certain immunodominant epitopes derived from HIV-1 are related with the development of strong CD8+ T cell responses. In fact, CD8+ T cells, restricted to epitopes derived from highly conserved regions of HIV-1 Gag, have a greater functional capacity, eliminating infected cells before de novo synthesis of viral proteins (20,77). This CD8+ T cell activation depends on the stability of the interaction between the epitope presented and the HLA-I molecule; for instance, an epitope with a low affinity for HLA-I leads to a decreased stability of the complex epitope/HLA-I, thus blocking the correct epitope presentation and the activation of CD8+ T cells (87).
The CD8+ T cell response leads to continuous viral evolution, allowing the selection of viral strains with the ability to escape immune recognition (10). This phenomenon known as immune escape, was described since 1991 (70), and has been observed in both acute and chronic phases of HIV infection (60). Phillips et al. described for the first time the selection of mutations within CD8+ T cell epitopes restricted to HLA-I molecules, resulting in decreased recognition of infected cells with the mutated virus, leading to a diminished production of IFN-γ by CD8+ T cells (70). From this study, a large number of mutations associated with immune escape restricted to HLA-I molecules have been described (Table 1). In general, these mutations are located in critical sites within the epitope that is recognized by CD8+ T cells, where an amino acid change leads to a diminished affinity of the epitope/HLA-I complex, affecting recognition by the TCR and the induction of a specific response (60).
The letter X indicates that in a given position, several amino acid substitutions have been described as escape mutations.
HLA-I, human leukocyte antigen class I; TCR, T cell receptor.
In the acute phase of HIV infection, at the time where the viral load reaches its peak, there is homogeneity of the viral sequences, which can be associated with lack of selective pressure in this phase. During this phase of infection, the CD8+ T cell responses are restricted mainly to epitopes derived from Env, Nef, and Tat, whereas during the chronic phase, the responses are restricted to epitopes derived from Gag (14,59,92). When the viral load starts to decrease, viral strains with escape mutations begin to predominate. Initially, the first viral strain populations with escape mutations correspond to those affecting recognition by the TCR; as infection progresses, this population is replaced by viral strains with mutations that affect the presentation by HLA-I molecules (18). The rate of selection of escape mutations tends to decrease as infection progresses to the chronic phase. Ganusov et al. reported that 1 year postinfection, the rate of viral escape was 10 times lower than the rate during the acute phase, which could be attributable to a lower elimination of infected cells by CD8+ T cells, a fitness cost of the selected mutations, and/or a higher production rate of virions in the acute phase compared with the chronic phase (36).
Specific CD8+ T cell responses associated with the selection of immune escape mutations are correlated with the cytotoxic profile of these cells; however, recent studies have shown that the noncytotoxic response of CD8+ T cells, specifically the response mediated by macrophage inflammatory protein-1β (MIP-1β), also has the ability to induce the selection of escape mutations (7,33,85). Although the rate of viral escape from the CD8+ T cell responses varies among hosts, certain factors play a pivotal role in the timing of the selection of escape mutations. It has been reported that immunodominant epitopes, that is, those epitopes that dominate the recognition events in a particular HLA-I molecule and, therefore, induce a strong CD8+ T cell response (1), tend to accumulate escape mutations faster than subdominant epitopes, which do not induce strong CD8+ T cell responses (53). Moreover, the Shannon entropy, which is defined as a measure of the population diversity at a given sequence in a specific position or within a cluster of bases/amino acids, is important for the selection of escape mutations, considering that epitopes with a high Shannon entropy tend to accumulate escape mutations faster than epitopes with low Shannon entropy (30,53). Furthermore, those epitopes with low entropy are typically subdominant, highlighting the strong effect that these two parameters have in the selection of immune escape mutations (53).
Types of HIV-1 Immune Escape Mutations
HIV-1 immune escape mutations can be classified according to the mechanism of evasion in three groups (Fig. 1).
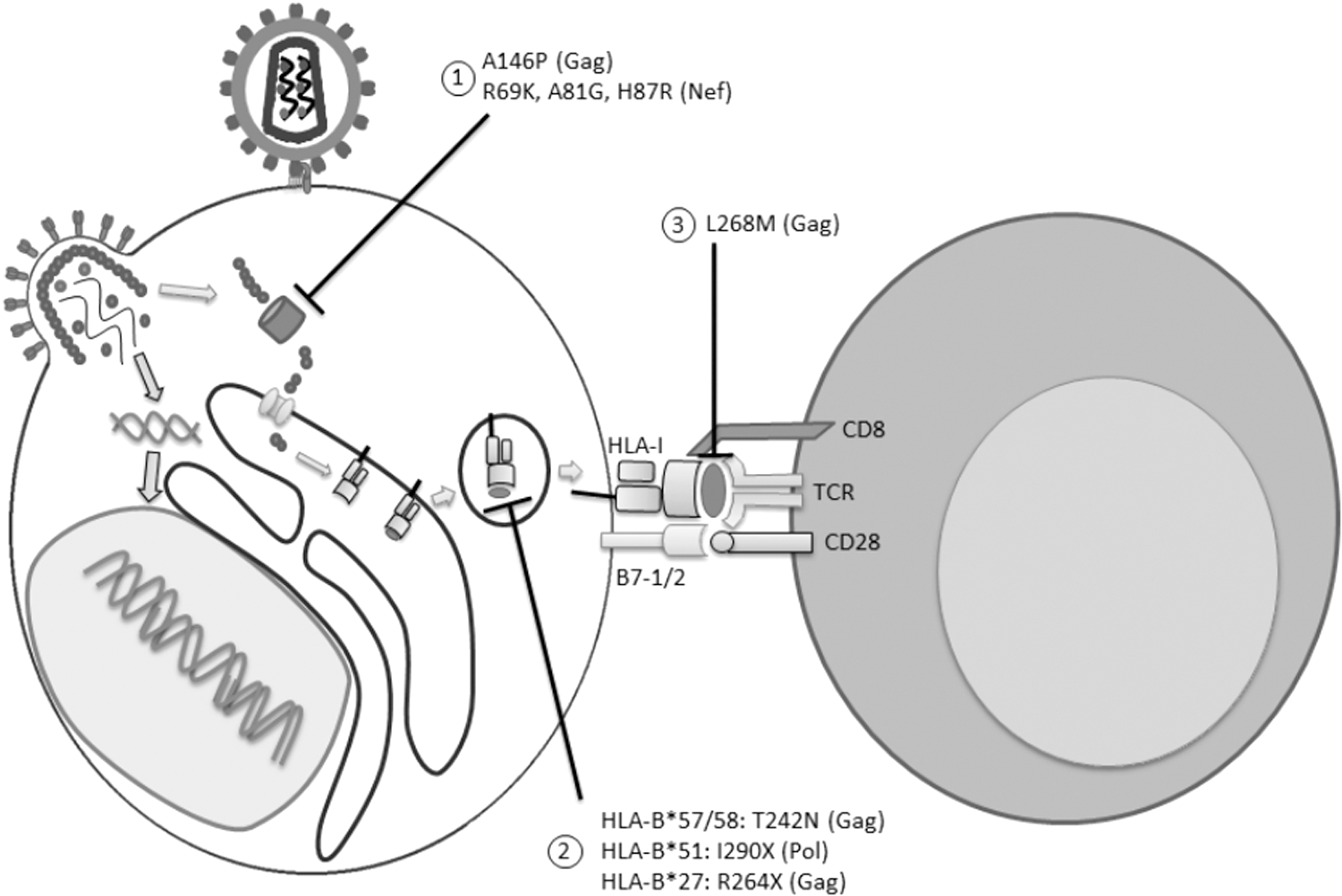
Types of HIV-1 immune escape mutations. Viral proteins present in the infected cell are processed through proteasome, leading to the generation of protein fragments that are transported to the endoplasmic reticulum through Transporter associated with antigen processing. Once in the endoplasmic reticulum, viral proteins are further processed by aminopeptidases into epitopes, which are loaded to the HLA-I molecule and transported to the cell surface for antigen presentation to CD8+ T cells. In several steps of this process, HIV-1 escape mutations can be selected to evade the immune response.
Mutations that affect the cellular processing of the peptide
These mutations generally flank epitopes restricted to HLA-I, which affect the cellular processing of the peptide in the cytoplasm or the endoplasmic reticulum, impeding the correct formation of the peptide in the infected cell (63,93). Several studies have described the selection of these mutations. Draenert et al. showed that in patients with the protective allele HLA-B*57:01, which presents the epitope ISPRTLNAW derived from Gag (IW9, residues 147–155), a mutation in the amino acid located immediately before the N-terminal portion of the epitope is selected (31). This mutation consists in a change of an alanine for a proline at position 146 (A146P), blocking recognition of the IW9 epitope by the aminopeptidase I in the endoplasmic reticulum, affecting its processing and preventing the formation of the epitope/HLA-I complex (31). Cellular processing of viral peptides influences the type of epitope that is going to be presented. Immunodominant epitopes recognized by CD8+ T cells suffer a specific processing in several domains that allows the formation of the epitope, whereas subdominant epitopes are most likely degraded in the cytoplasm, impeding the docking with the HLA-I molecule in the endoplasmic reticulum. Moreover, these subdominant epitopes are susceptible to be degraded by the proteasome, as the inhibition of this cellular machinery leads to an increased presentation of subdominant epitopes by dendritic cells (25). This preferential processing of immunodominant epitopes can be explained by the presence of specific N-terminal motifs in the precursor protein of the epitope, as when these motifs are integrated adjacent to a subdominant epitope, it leads to a higher production of the epitope (49).
Given that the machinery involved in the generation of epitopes restricted to HLA-I is highly conserved in humans, the adaptation of HIV-1 through a mutation that allows the escape of this processing would be favored, as the same mutations could contribute to the evasion of the immune response in different hosts, whether or not they express the same HLA-I allele (18). However, the frequency of such mutations at the population level is low (82), which can be explained by the type of epitope that is presented by the HLA-I molecule, as each of these molecules presents a different epitope that derives from a different protein precursor. The mutations affecting this processing of the epitope tend to revert when a virus with this type of mutation is transmitted between HLA-I discordant individuals, as the protein precursor that results in the formation of the epitope is different in the recipient, resulting in lack of selective pressure (81).
Mutations that affect the presentation by HLA-I
Most of the HIV-1 immune escape mutations have been associated with a decrease in the affinity between the epitope and the HLA-I molecule that arises by the selective pressure exerted by CD8+ cells restricted to HLA-I molecules. They are located mainly in the second position or in the C-terminal domain of the epitope, since these positions show the main interactions with the HLA-I molecule, being recognized by B and F pockets, respectively (55). For protective HLA-I alleles, some amino acids bind to these pockets and are related with the protective effect of the allele. The HLA-B*27 allele presents peptides with arginine in position 2 and hydrophobic amino acids in the C-terminal domain, limiting the peptide presentation to those derived from highly conserved protein regions, where a mutation in these peptides results in a functional alteration (65). The HLA-B*57/58:01 allele preferentially presents peptides with tryptophan in the C-terminal domain (55). The preference of protective alleles to present certain peptides is highly related to the stability between the peptide and the HLA-I molecule that leads to correct induction of a specific CD8+ T cell response, with the ability to control viral replication (4).
HIV-1-infected patients who express HLA-B*57:01 mount a specific CD8+ T cell response against the TSTLQEQIGW epitope derived from Gag (TW10, residues 240–249). Leslie et al. demonstrated that the change of threonine to asparagine at position 242 (T242 N) drastically decreases the stability of the epitope/HLA-I molecule complex. When a viral strain with the T242 N mutation is transmitted to a recipient without the HLA-B*57:01 allele, this mutation reverts to a wild-type phenotype (50). Similarly, HIV-1-infected individuals expressing HLA-B*27:05 that presents the epitope KRWIILGLNK derived from Gag (KK10, residues 263–272), select viral strains with the mutation R264G/K/Q, consisting in a change from arginine to glycine, lysine, or glutamine at position 264, resulting in a decrease in the affinity of the epitope/HLA-I complex that reverts once a viral strain with any of these mutations is transmitted to an HLA-discordant host (5,6). These observations suggest that the protective effect of these types of HLA-I alleles is mediated in part by the selection of mutations in highly structurally conserved regions of HIV-1.
Protective HLA-I alleles also induce the selection of escape mutations in subdominant epitopes. Payne et al. described the selection of three different mutations in the KRKGGIGGY epitope derived from Pol (KY9, residues 901–909), which is restricted by HLA-B*27:05. The R902K (substitution of an arginine for a lysine at position 902), K903R (substitution of a lysine for an arginine at position 903), and G908E (substitution of a glycine for a glutamic acid at position 908) mutations were enriched in patients expressing HLA-B*27:05, and lead to a higher dissociation rate of the epitope for the HLA-I molecule (69). It is noteworthy that the selection of this type of mutations within subdominant epitopes seems to be restricted to protective HLA-I alleles, as several studies performed with different HLA-I alleles not associated with disease progression have not found a selection of escape mutations in the corresponding epitopes (30,79), which display highly conserved sequences at the population level (54,89).
It is important to highlight that the associations of HLA-I alleles with HIV-1 progression or with HIV-1 escape mutations can be specific, depending on the predominant viral subtype of the population. Historically in Japan, where the predominant viral subtype is B, the HLA-B*51:01 allele has been associated with slower progression of infection, whereas in Sub-Saharan Africa, where HIV-1 subtype C prevails, this allele has been associated with fast progression (19). Similarly, in a Kenyan cohort of typical progressors expressing the protective alleles HLA-B*57:01 or -B*58:01 and infected with HIV-1 subtype A, there was a selection of the P243T (change from proline to threonine at position 243) and I247 L (change from isoleucine to leucine at position 247) mutations in the TW10 epitope, whereas in patients with the same alleles, but infected with HIV-1 subtype B, the T242 N mutation was selected in the same epitope (58). This can be explained by the fact that the TW10 epitope is different in HIV-1 subtype A, in comparison with HIV-1 subtype B or C, where, in subtype A, there is a proline at position 243, whereas in subtypes B and C there is a leucine at the same position. The T242 N mutation in the TW10 epitope in HIV-1 subtype A is not viable in terms of viral fitness, explaining why other mutations are selected, in particular, those that do not carry a major negative impact in viral fitness, allowing the virus to escape the immune response (58).
Mutations that affect recognition by the TCR
There is another type of HIV-1 immune escape mutations that does not affect the processing of the peptide or its affinity for the HLA-I molecules, but that does affect the recognition by the TCR. In the KK10 epitope restricted to HLA-B*27:05, the mutation L268 M (change of a leucine for a methionine at position 268) is selected. This mutation does not affect the affinity of the epitope for the HLA-I molecule; instead, it affects recognition of the epitope/HLA-I complex by the TCR (40). Nevertheless, these types of mutations are not always associated with immune escape from the CD8+ T cell responses. It has been reported that there are several clones of CD8+ T cells with different TCR rearrangements that can recognize mutated epitopes (29,83). Likewise, there are CD8+ T cell clones that carry TCR molecules allowing recognition of wild-type as well as mutated peptides, without affecting the functional capacity of these cells (16); this is the case reported for HIV-1 controllers (71).
Viral and Clinical Implications of HIV-1 Immune Escape Mutations
The association of escape mutations with viral fitness and the selection of compensatory mutations
Even though most of the immune escape mutations provide an evolutionary advantage for the virus, many of them also have a negative impact in viral fitness (51). One example is the T242 N mutation selected in the TW10 epitope, restricted to HLA-B*57:01 or -B*58:01 that destabilizes the helix 6 of p24 (57). However, to efficiently evade the immune response without affecting viral fitness, other mutations are selected to restore viral infectivity. These mutations are epitope specific and can be selected within the epitope or close to it. The substitution of glycine for alanine at position 248 (G248A) in the TW10 epitope is only selected in HIV-1-infected patients that express HLA-B*57:01 or -B*58:01. The G248A mutation alone is not associated with immune escape nor with a compensatory effect; however, the combination of mutations T242 N and G248A in the TW10 epitope leads to total loss of the epitope affinity for HLA-I, without affecting viral fitness (57). Likewise, the R264K mutation in the HLA-B*27:05-restricted epitope KK10 has a negative impact on viral fitness, which is compensated by the selection of the S173A mutation (change of serine for alanine at position 173), upstream the epitope (84).
HIV-1 compensatory mutations associated with immune escape mutations are not unique; as for each epitope, several mutations can be selected to partially or fully restore viral fitness. Brockman et al. determined that the mutations, H219Q (change of histidine for glutamine at position 219), I223 V (change of isoleucine for valine at position 223), and M228I–L (change of methionine for isoleucine or leucine at position 228), have the same ability to compensate the deleterious effect caused by the T242 N mutation in the TW10 epitope (13); this suggests that there are different mechanisms of selective pressure for the selection of a given compensatory mutation. The selection of compensatory mutations may also explain why some patients with protective HLA-I alleles progress in a similar way to typical progressors. It has been reported that in typical progressors expressing the protective alleles HLA-B*57 or -B*58:01, viral strains with compensatory mutations in epitopes restricted to those alleles appear with a high frequency (18). The selection of immune escape mutations along with compensatory mutations highlight the ability of HIV-1 to adapt to the different sources of selective pressures exerted by the host, which can be related to the ability of the virus to adapt at the level of a population.
The role of HLA-I in the adaptation of HIV-1 at a population scale
As discussed so far, the specific CD8+ T cell response restricted to HLA-I plays a crucial role during HIV-1 adaptation to the host. However, if a given HLA-I allele is highly prevalent in a population, it is plausible to infer that escape mutations selected by that HLA-I allele would also be highly prevalent in the population. Moreover, were there to be a high prevalence of protective HLA-I alleles in a population, the circulation of preadapted viral strains with escape mutations for that specific allele would decrease the protective effect. In Japan, where the prevalence of the HLA-B*51:01 allele in HIV-1-infected individuals is about 22%, there is a high frequency of the I135X mutation in the TAFTIPSI epitope derived from Pol (TI9, residues 283–290). This has led to a change in the protective effect of the HLA-B*51:01 allele, since it was related with lower viral loads and, therefore, with a slower progression of the infection between 1997 and 2008; however, in the recent years, this association was lost (43). Similar studies have been performed in different cohorts, such as in Barbados and South Africa, where the high prevalence of protective HLA-I alleles has favored the selection of viral strains with immune escape mutations in the populations, negatively affecting the protective effect of specific HLA-I alleles (41,68). Regarding protective HLA-I alleles targeting subdominant epitopes, it has been demonstrated that the high frequency of such alleles in a population also leads to the selection of escape mutations and the loss of the protective role. A study from a South African cohort demonstrated that the high frequency of HLA-B*15:03, estimated at 14% in this population, has led to the accumulation of escape mutations in epitopes derived from Gag, Nef, Tat, and Pol at the population level, and that patients expressing this particular allele have significantly higher viral loads compared with patients not expressing it. However, in the United States, this allele has been associated with lower viral loads, as the low frequency of the allele, estimated at around 1%, has not allowed the selection of such mutations (35). Similar results have been observed in HLA-A*68, which is the most prevalent HLA-A allele in South Africa and that is associated with higher viral loads in this population, as a result of the selection of escape mutations in subdominant epitopes (45).
In order for this phenomenon of fixation of escape mutations in a population to occur, the distribution of HLA-I alleles should be homogenous within the population. For example, in the United States, where the population is highly heterogeneous, the frequency of escape mutations has slightly increased in the last three decades, which have not favored viral adaptation in this population throughout that time (22).
Considering that some escape mutations selected by the pressure of a protective HLA-I allele carry a negative impact on viral fitness, one can stipulate that viral strains adapted to a highly frequent protective HLA-I allele in a population could be less virulent than those viral strains in populations where the frequency of the same HLA-I allele is not high. Payne et al. demonstrated that in Botswana, the circulation of HIV-1 strains with escape mutations associated with HLA-I-protective alleles has led to a significant decline in viral fitness in that country (68). Nevertheless, a meta-analysis performed by Herbeck et al. showed that since 1984 the markers associated with disease progression (viral load and CD4+ T cell count) have not changed significantly (39), suggesting that the virulence of HIV-1 has been constant since the introduction of the virus into the human population in 1920. So far, there are no studies showing the frequency of compensatory mutations associated with escape mutations at the population level, which could clarify the role of these mutations in HIV-1 adaptation.
HIV-1 immune escape mutations and their implications in the clinical course of the infection
The selection of escape mutations restricted to HLA-I in HIV-1 may have a negative impact in the clinical course of the infection. Goulder et al. described that the selection of the R264X mutation in the HLA-B*27:05-restricted epitope KK10 derived from Gag, was associated with a decrease in the time of progression to AIDS in patients expressing the allele (37). Likewise, in a 10-year follow-up of an HIV-1 controller patient expressing HLA-B*27:05, it was shown that the CD8+ T cells specific for the KK10 epitope had a drastic decrease in frequency around the 7th year of the study, a phenomenon that was associated with the presence of the R264X mutation in the viral strains of the patient (32). It has been reported that the transmission of a virus with escape mutations between individuals with the same HLA-I is associated with faster progression to AIDS in the recipient, as viral strains are preadapted to the CD8+ T cell responses (17,23). Although these results suggest an impact of HIV-1 immune escape mutations in the clinical course of the infection, the emergence of these mutations only explain about 6% of the variation in the plasma viral load, as determined by a mathematical model (42).
The Impact of HIV-1 Immune Escape Mutations in the Development of New Therapeutic Strategies
HIV escape mutations in the development of CD8+ T cell-based vaccines
Considering that the selection of escape mutations in HIV-1 by CD8+ T cell-specific responses may lead to a decrease in viral fitness, novel vaccine strategies have been implemented based on immunodominant and conserved HIV-1 epitopes to induce such specific responses. However, the high genetic variability of HIV-1 has been the biggest hindrance for this kind of therapy, as HIV-1 strains with escape mutations for the CD8+ T cell response generated can be selected in the vaccinated individual (3). Several vaccine strategies have been developed so far that induce specific CD8+ T cell responses. Many of them have been implemented using expression vectors coding for HIV-1 proteins, since using an attenuated HIV-1 virus represents a high risk of infection for the individual (80). The Poxvirus-based vaccines have been used based on their capacity to adapt high amounts of foreign genome (91). The NYVAC (derived from a highly attenuated Vaccinia virus strain) and the modified vaccinia Ankara (MVA) vaccines expressing the Gag, Pol, and Nef proteins of SIVmac239 have been tested in primates, where NYVAC induces predominantly CD4+ T cell responses, whereas MVA induces mainly CD8+ T cell responses (64). In a United Kingdom cohort, it was demonstrated that the MVA-HIVconsv vaccine, which expresses 14 highly conserved peptides derived from HIV-1 subtype A, B, C, and D, was safe and induced strong CD8+ T cell responses (11). However, the efficacy of Poxvirus-based vaccines has been questioned, since in a Thai cohort at high risk of acquiring HIV-1 infection, where the ALVAC (Attenuated Canarypox virus, expressing Gag and Pol from HIV-1 subtype B) vaccine was tested, in only 26% of the vaccinated individuals was a protective effect achieved (75). This low efficacy of Poxvirus-based vaccines can be explained by the fact that, once a foreign genetic material is introduced in the vector, it loses its immunogenicity; therefore specific CD8+ T cell responses are rarely generated. For this reason, Poxvirus-based vaccines have not gone through phase IIb/III clinical trials (80).
Another vaccine strategy is the use of human adenoviruses as expression vectors of HIV-1 proteins, especially human adenovirus 5 (Ad5). Merck developed the trivalent Ad5-HIV gag/pol/nef vaccine, which elicited strong CD8+ T cell responses in phase I clinical trials (66). However, the use of Ad5 as an expression vector can be affected by the seroprevalence of Ad5 in the population, as the immunogenicity of the vector would be affected in a vaccinated individual who has neutralizing antibodies from a previous Ad5 infection (80). Another issue that this vaccine faces is safety; a study in America and Australia determined that people vaccinated with Ad5-HIV gag/pol/nef had a higher risk of contracting HIV-1 infection than those individuals who received the placebo, although this effect could be attributed to the presence of neutralizing antibodies against Ad5 in vaccinated individuals (15). Methodologically, one big issue that Ad5- and Poxvirus-based vaccines face is the introduction of large foreign genomic sequences, as it can affect the replicative capacity of the vector. Also, the introduction of full genomic sequences of HIV-1, such as gag or pol, can diminish the effectiveness of the vaccine, due to the high variability of the virus in certain regions of the genome (52). For this reason, new vaccine strategies have been developed based only on highly conserved epitopes of HIV-1, as escape mutations in these epitopes would decrease viral fitness (46). Nevertheless, using just a narrow set of epitopes would result in a modest protection at the population level, since it would only benefit those individuals able to present the epitopes in their HLA-I repertoire. To solve this problem, mosaic vaccines were developed; these are based on the in silico prediction of the most significant conserved epitopes that circulate in a population, to generate broader CD8+ T cell responses and cover the genetic diversity of HIV-1 in the population (46). Barouch et al. showed that in Rhesus macaques a mosaic vaccine based on Adenovirus 26-expressing peptides derived from Gag, Pol, and Env induced a broader CD8+ T cell response than vaccines based on consensus sequences (8). Nevertheless, the conserved feature of a protein does not always associate with a decrease in viral fitness once an escape mutation is selected (76), pointing to the need for further studies to create a functional vaccine that induces broad CD8+ T cell responses.
Role of HIV escape mutations in the elimination of viral reservoirs
Another therapeutic strategy targeted to eliminate HIV from infected hosts consist in the reactivation of latent HIV harbored in cellular reservoirs, which are the responsible of the long-life maintenance of the virus (86). The main cellular reservoirs of HIV are resting CD4+ T cells, although monocytes, macrophages, hematopoietic stem cells, and other cells, can act as reservoirs of the virus (2). These cells maintain HIV in a latent state mainly by chromatin condensation, low availability of transcription factors, and posttranslational mechanisms (28). The identification of these mechanisms has led to the development of new therapeutic strategies that disrupt latency, which include histone deacetylase inhibitors, activators of protein kinase C, and cytokines such as IL-2 and IL-15 (74). This strategy has been denominated “shock and kill,” as once the virus is reactivated, the infected cell can be eliminated by viral cytopathic effects, through CD8+ T cell-mediated killing, and/or by limiting the infection of neighbor cells using HAART (12). However, if the cellular reservoir were seeded with viral strains with escape mutations, the CD8+ T cell response would not be able to target these infected cells, rendering this strategy ineffective. Moreover, the reservoir size is variable among patients, and the most important factor limiting the size of the reservoir is HAART, as an early initiation of this therapy substantially decreases reservoir size (12). Deng et al. have recently demonstrated these features, as they found that in patients initiating HAART during the chronic phase, there was presence of viral strains with escape mutations, which were replicative competent upon reactivation of resting CD4+ T cells (24). Similar results were obtained by Papuchon et al., who showed that in patients under effective HAART, there was presence of escape mutations in the cellular reservoir. Interestingly, although in some patients during the acute phase there was a heterogeneous viral population in plasma, containing both escaped virus and wild-type virus, only the virus with escape mutations seeded the cellular reservoir (67). In order for CD8+ T cells to kill these latently infected cells, a broadly functional profile of CD8+ T cells able to recognize other nonmutated epitopes have to be employed. This strategy has been published recently, showing an improvement of killing of latently infected cells using broadly functional CD8+ T cells, compared with CD8+ T cells specific for mutated epitopes (88).
Altogether, these results demonstrate that for the “shock and kill” strategy to be efficient, a thorough analysis of the sequences of HIV harbored in cellular reservoirs is necessary, to improve the effectiveness of CD8+ T cell-mediated killing of these cells.
Concluding Remarks
Diverse selective forces of the immune system, particularly the CD8+ T cell responses restricted to HLA-I molecules, select HIV-1 immune escape mutations. Although these mutations represent a barrier for the effective killing of infected cells, they have also helped to identify conserved regions in HIV-1 that may offer key insights in the development of new therapeutic strategies based on the induction of specific CD8+ T cell responses. Moreover, the identification of viral strains with escape mutations in the cellular reservoirs is crucial for novel strategies based on the elimination of latent virus. Nevertheless, so far these new strategies have not proven to induce a polyfunctional CD8+ T cell profile or to completely eliminate the viral reservoir, pointing the requirement of further studies to define a functional and sterilizing therapy for HIV-1.
Footnotes
Acknowledgments
The authors thank CODI (2014-919) of the Universidad de Antioquia, UdeA, for its financial support. The authors wish to thank Anne-Lise Haenni for her constructive comments.
Author Disclosure Statement
No competing financial interests exist.