
Abstract
Nucleic acid recognition by toll-like receptor 9 (TLR9) initiates signaling pathways that regulate the production of proinflammatory cytokines or type I interferons, as well as many other molecules required to initialize the immune response. The use of synthetic oligodeoxynucleotides (ODNs) has been crucial to emulate the recognition of DNA sequences by TLR9. Furthermore, ODN administration to mice has shown to confer protection against a wide range of viral, bacterial, and parasitic pathogens. In contrast, oncogenic DNA viruses like hepatitis B virus, Epstein–Barr virus, and human papilloma virus inhibit TLR9 expression, thus contributing to the establishment of chronic viral infections. In this review, we will focus on TLR9 signals initiated by ODN recognition, on the inhibition of TLR9 expression mediated by DNA oncogenic viruses, and on TLR9 expression as a relevant event in the progression to cancer, considering other functions of this receptor, aside from viral recognition.
Introduction
T
Highly relevant among TLRs is TLR9, member of a receptor subgroup that recognizes nucleic acids (29). Canonically, TLR9 recognizes unmethylated CpG DNA sequences, but recent evidences indicate that TLR9 is able to recognize other types of nucleic acids, which means that the recognition of nucleic acids by TLR9 is more sensitive than what was initially thought.
TLR9 Structure and Ligand Binding
TLR9 is a protein with an N-terminal ligand recognition domain shaped for a leucine-rich repeat (LRR) domain, a 20-amino acid helical transmembrane domain, and a cytoplasmic signaling Toll/IL-1 receptor (IL-1R) homology domain (29). In TLRs, the LRR domain is a 22–29-aa sequence containing hydrophobic residues and forming a loop structure; when assembled as a full protein, tandem LRR copies are arranged as a solenoid in a curved configuration (horseshoe shape), which is constituted by 25 LRRs in TLR9 (6).
Since no crystallographic structure has been reported yet for TLR7, 8, and 9, the location of the ligand binding site to the LRR domain is not known. However, Wei et al. developed a three-dimensional structure of the LRR domain of TLR7, TLR8, and TLR9 based on homology modeling; according to their results, ligand binding region in TLR9 is located in the proximity of LRR 17 (81).
In vivo analysis with single-molecule fluorescence tools showed that TLR9 is predominantly located in endosomal compartments, remaining as a homodimer before and after stimulation with CpG-B oligodeoxynucleotide (ODN) in HEK293 cells (9). Results obtained by Chen et al. suggest that a conformation change is necessary for TLR9 activation, and this change occurs after binding CpG sequences (9).
The study of the signaling pathways activated by TLR9 after DNA recognition involves the extensive use of synthetic ODNs with a phosphothioate (PS) instead of a phosphodiester (PO) backbone to improve cellular uptake and nuclease resistance. Nonetheless, the cell response to both kinds of ODNs is different; for instance, PS-ODNs showed an effect on macrophage viability and the activation of the IL-12 promoter at lower concentrations than PO-ODNs; on the other hand, the CpG PO-ODN activity was inhibited by non-CpG PS-ODN, whereas non-CpG PO-ODN failed to show such effect (65). In the same way, while stimulus with 1,668 PS-ODN induced higher IL-6 production in murine B cells than 1,668 PO-ODN, an inverted CpG sequence in 1,668 PS-ODN did not abrogate IL-6 production (59). Similarly, Vollmer et al. showed that non-CpG PS-ODN induced proliferation of human B cells and the secretion of IL-6 and IL-10 (78). Thus, both CpG and non-CpG ODNs are potent IL-6 and IL-10 inductors in B cells.
With respect to the ODN methylation status, methylated CpG PS-ODNs (mCpG ODN) and unmethylated CpG PS-ODNs were reported to be taken up similarly by spleen and lymph node cells (13). Importantly, when each kind of ODN was colocated with TLR9 in the late endosome of mouse spleen-derived dendritic cells (DCs), both of them induced the production of interferon γ (IFNγ), IL-6, and MCP-1 (13).
Coch et al. reported that PO-ODN complexed with DOTAP (N-[1-(2,3-dioleoyloxy)]-N,N,N-trimethylammonium propane methyl-sulfate) induced the production of IFNα and tumor necrosis factor (TNF) in plasmacytoid dendritic cells (pDCs) and elimination of the CpG motif abolished IFNα production, whereas stimulus with CpG-methylated ODN reduced IFNα production in the same cells (11). Furthermore, pDCs recognized genomic self and bacterial DNA complexed with cationic molecules, but activation was lower when DNA was methylated.
These antecedents suggest that stimulus with PS-ODNs has delayed, at least partially, our understanding on the effect of the recognition of naturally occurring DNA by TLR9. In this regard, a very elegant work by Haas et al. showed that TLR9 recognizes DNA sugar 2′ backbone deoxyribose ODN, and this recognition drives cytokine production in the same extent as PS-ODN, disregarding CpG sequences (23). They reported that the addition of a 3′ PD poly-G extension (24 Gs) improved ODN cellular uptake and prevented nuclease sensitivity. It is remarkable that 3′ PD poly-G ODN induced TLR9 activation and cytokine production in a CpG sequence-independent manner, and this activation was not an effect of the poly-G sequence (23).
Interestingly, a recent work has shown that TLR9 is able to recognize DNA:RNA hybrids (58). Stimulation of plasmacytoid and conventional dendritic cells with a DNA:RNA hybrid containing a guanosine–uridine-rich RNA strand motif induced the expression of CD40 and CD86 in both types of cells, and it also induced the production of IFNα and IL-6 in a TLR9-MyD88-dependent manner; this recognition occurs in endosomal compartments (58). Since ssDNA has no cytokine stimulating effect, the authors concluded that TLR9 is able to sense the intact DNA:RNA hybrid. In agreement with the work of Haas et al., these results may also suggest that duplex ODN with at least one DNA PO backbone could trigger TLR9 signaling (23).
Altogether, these works support the idea that the endosomal location of TLR9 avoids self-DNA recognition, although the controversy about the selectivity of non-mCpG DNA sequences versus mCpG DNA sequences remains. The differences in cytokine production levels in different cells could be explained by the differential expression of TLR9 isoforms and the machinery inherent to each cell type. With respect to the TLR9 selectivity for DAMPs, TLR9 seems to act as a promiscuous molecule, recognizing self DNA, foreign DNA, PS-ODN, PD-ODN, and even DNA:RNA hybrids, as well as CpG motifs or motifs different to CpG sequences (11,13,58,78).
TLR9 Signaling
TLR9 is expressed in intracellular compartments of pDCs, monocytes, B lymphocytes, T lymphocytes (CD4+ and CD8+), endothelial cells, keratinocytes, and melanocytes (19,31,50,84); also, it is expressed on the surface of tonsil and peripheral B lymphocytes, splenic DCs, gastric, intestinal, and cervical epithelial cells (3,15,22,40,55,64).
Before stimulation, TLR9 resides in the endoplasmic reticulum, and it is translocated to the Golgi complex and lysosomes after stimulus with ODNs (either CpG or non-CpG), where it colocates with MyD88 (10,39,42). TLR9 translocation from endoplasmic reticulum to the endolysosome is dependent on the interaction with UNC93B1 (Unc-93 homolog B1); therefore, when UNC93B1 is not functional, there is no cytokine production in response to CpG ODNs (7,36,70). It should be noted, as well, that UNC93B1 trafficking is dependent on the adaptor proteins (AP) 1 and 2, by the C-terminal tyrosine-based sorting motif (Yxxφ) (57). TLR9 is a 150 kDa protein that undergoes proteolytic cleavage in the endolysosome, generating an 80 kDa protein; this modification is dependent on asparagine endopeptidase and cathepsin family members (16,57,66). Cleavage generates an N-terminal and a C-terminal TLR9 fragments, which remain associated; such association is required for DNA sensing and signaling transduction (55). Thus, the formation of the TLR9N+C complex and the exclusion of TLR9 from the cell membrane are mechanisms to avoid potential recognition of self DNA and autoimmune diseases (4,55).
TLR9 ligand binding induces two different signaling pathways with common protein components. In both pathways, TLR9 recruits the AP MyD88, which interacts with IL-R-associated kinase 4 (IRAK4) through its death domains and activates IRAK; in turn, this induces the recruitment of TNF receptor-associated factor 6 (TRAF6) (28,30,44,46).
In the MAPKs pathway, TRAF6 is ubiquitinated by E1 and E2 (Ubc13/Uev1A), promoting the recruitment of TAB2 and TAB3, which mediate the activation of transforming growth factor (TGF)-associated kinase 1 (TAK1) (14,33). TAK1 activates IκB kinase β (IKKβ), inducing the translocation of NFκB to the nucleus, and activates MKK3/6, leading to the activation of the JNK-p38 kinase pathway (14,80). In addition, TRAF6 activation leads to the activation of IRF5, which in turn induces the expression of IL-6, IL-12, and TNF (71).
In the type I IFN pathway, PYKfive, the complex TRAF3, AP-3, and IRF7, along with the DOCK2 protein (which activates IKKα), osteopontin, and viperin, are activated during IFNα production in plasmacytoid cells (21,24,27,32,63,68,74).
TLR9 Expression in Oncogenic DNA Virus Infection
Human papilloma virus
TLR9 activation in keratinocytes induces the expression of TNF, IL-6, and IL-8 (69). However, Hasan et al. demonstrated that VPH16 E6 and E7 proteins inhibit the transcription of TLR9 in keratinocytes, and as a consequence, there is no secretion of IL-8 or MIP3a in response to CpG ODN (25). Moreover, TLR9 inhibition mediated by human papilloma virus 16 (HPV16) E6 and E7 was confirmed in a C33 cell line infected with HPV16 quasivirions (26). The mechanism underlying this event was elucidated by chromatin immunoprecipitation, showing that TLR9 inhibition was mediated by the recruitment of a complex that included the NFκB transcription factor (p50–p65), the histone deacetylase HDAC1, and the JARID1B demethylase to the TLR9 promoter (26).
In a similar manner, beta HPV38 abolished TLR9 expression, as demonstrated in RPMI8226 human myeloma cells and in human foreskin keratinocytes (HFK) transiently transfected with HPV38 E6 and E7 (56). Accordingly, IL-8 and MIP3a secretion were undetectable in HFK expressing HPV38 E6 and E7, and stimulated with CpG ODN (56). Chromatin immunoprecipitation analysis showed that an inhibitory complex formed by ΔNp73α (p53 antagonist), IKKβ, and the polycomb protein enhancer of zeste homolog 2 (EZH) is recruited to the promoter of the TLR9 gene (56).
Although chronic HPV infection is associated with the development of epithelial lesions and cervical cancer, the dynamics of TLR9 expression during HPV infection and the progression of intraepithelial lesions to cervical cancer remain to be studied.
Daud et al. found that TLR9 expression in cervical cells was increased in women (American population) within 4 months after clearing an HPV16 infection, but TLR9 expression decreased in women with persistent infections (classified as such after two positive HPV16 results in an 8-month period) (12). Remarkably, TLR9 basal expression levels were higher in samples from women with persistent infections with respect to women who cleared the infection (12).
Evaluation of TLR9 expression in cervical cells from women positive for high-risk (HR) and low-risk (LR) HP, with a previous HPV-positive test, showed that women positive for LR HPV had significantly higher TLR9 levels with respect to negative HPV samples, while TLR9 expression in HR HPV-positive samples was marginally higher with respect to the control group (Italian population) (8). Importantly, when TLR9 expression was evaluated in samples from women with HPV persistent infections (6–18 months), TLR9 expression levels were higher in patients who had a persistent infection. These apparently contradictory results can be explained by the fact that oncogenic DNA viruses such as HPV16, hepatitis B virus (HBV), and HSV-2 inhibit TLR9 expression in a transient manner (85).
At the protein level, TLR9 was detected by immunohistochemistry in 53 cervical biopsy, conization, or hysterectomy samples from Tunisian women. Five cases of CIN I, 6 cases of CIN II, 7 cases of CIN III, and 22 cases of cervical carcinoma were compared with 7 cases of condyloma and 6 normal cervical squamous epithelium samples (18). The results showed higher TLR9 levels in samples from CIN and cervical carcinoma patients with respect to normal cervical squamous epithelium (18).
Another study performed in a Korean population showed a statistically significant increase of TLR9 expression in cervical carcinoma tissue with respect to normal cervical epithelium (41). Furthermore, an increase in TLR9 expression was detected by immunohistochemistry, in good agreement with the severity of the disease; this is, low-grade intraepithelial lesion < high-grade intraepithelial lesion < invasive squamous cell carcinoma; 70% of the invasive cell carcinoma showed a moderate to strong immunoreactivity score for TLR9 (41). Likewise, TLR9 assessment by immunostaining in a Indian population (n = 9) showed higher TLR9 protein expression in agreement with the severity of the disease; specifically, tumor tissue samples from squamous cervical carcinoma patients expressed higher TLR9 levels than those from CIN II and CIN III patients, with respect to control samples (20). Accordingly, a recent case–control study showed that TLR9 protein levels was increased in women with cervical cancer (n = 30) in comparison with controls (n = 30) (Hindu population) (2).
A recent study demonstrated that TLR9 expression in peripheral blood mononuclear cells (PBMCs) is higher in patients with squamous intraepithelial lesion and cervical cancer and higher TLR9 expression levels in the same cells are significantly associated with a greater risk for developing squamous intraepithelial lesions and cervical cancer in a Mexican population (Martínez-Campos, unpublished), suggesting that the increase in TLR9 expression in PBMCs could be correlated with the augment of TLR9 expression in tumors. Furthermore, TLR9 serum concentration increased in patients diagnosed with cervical cancer (n = 20) with respect to healthy donors (n = 20) (Mexican population) (Martínez-Campos, unpublished).
These works suggest that although oncogenic HPV proteins inhibit TLR9 expression during infection, TLR9 increases during the progression of cervical cancer. Remarkably, TLR9 could represent a new biomarker for cervical cancer progression since changes in the TLR9 protein can be detected in the periphery.
Hepatitis B virus
HBV is an oncogenic virus, capable of diminishing TLR9 expression in pDCs and B cells, blocking IFNα and IL-6 production (77). In agreement with this, TLR9 relative expression in PBMCs was significantly lower in patients with chronic HBV infections (n = 60) with respect to healthy donors (n = 60) in an Iranian population (61). The TLR9 protein was not detectable when assessed in PBMCs from five chronic HBV carriers and six HBV-associated hepatocellular carcinoma (HCC) patients, in comparison with TLR9 protein expression levels in controls by Western blot (Asian population) (77). On the contrary, flow cytometry analysis of TLR9 protein expression in PBMCs from Chinese HCC patients (n = 11) showed higher TLR9 expression levels with respect to healthy controls (n = 11). In this study, TLR9 protein levels in PBMCs were increased in agreement with disease severity (HCC > HBV related liver cirrhosis > chronic HBV infection > healthy donors) (83). Furthermore, other studies have reported that TLR9 is overexpressed in tumor cells from HCC patients (16,45,53). Interestingly, Liu et al. found that TLR9 is activated in response to mitochondrial DNA released under hypoxic conditions, and this activation induced the proliferation of tumor hepatocellular cells (45).
Hence, as it was the case for HPV and cervical cancer, HBV downregulates TLR9 during infection, but TLR9 is overexpressed in tumor cells, at least in part, because of hypoxic conditions, suggesting that TLR9 inhibition is no longer necessary once the cell is transformed. Again, TLR9 increase in the periphery could be an indicator for a cell transformation event.
Epstein–Barr virus
In vitro, human B cells infected with Epstein–Barr virus (EBV) acquire the capacity to proliferate indefinitely, generating the lymphoblastoid cell line (LCL). TLR9 quantification in LCLs has shown that this cell line had lower TLR9 expression levels than EBV-free primary B cells; consequently, TNF and IL-6 production after stimulation with CpG ODN (17) were abolished in LCLs. In EBV-infected RPMI8226 and primary B cells, the molecular mechanism of TLR9 modulation involved the EBV latent membrane protein 1 (LMP1), including the activation of the NFκB transcription factor, mediated by IKKα and IKKβ, since an overexpression of dominant negative mutants of these proteins restored the TLR9 promoter activity (17). Remarkably, LMP1 activated JNK1, and in turn activated p73, which positively activated ΔNp73α (1), suggesting that ΔNp73α is recruited to the TLR9 promoter, as observed for HPV38 (56).
In a similar way, another study reported that EBV downregulated TLR9 at mRNA and protein levels and the early lytic protein BGLF5 contributed to this downregulation during a productive EBV infection (76).
While EBV is associated with Burkitt's lymphoma, Hodgkin's disease, posttransplant lymphoma, nasopharyngeal carcinoma, and some forms of gastric carcinoma, TLR9 expression in patients with these types of cancer remains to be assessed.
Merkel cell polyomavirus
Early proteins of Merkel cell polyomavirus (MCPyV) inhibit TLR9 expression in a promoter-dependent manner (66). Specifically, transient transfection experiments with a reporter TLR9 promoter plasmid showed that large T antigen (LT) and small T antigen (sT) early expression MCPyV proteins were responsible for this inhibition; thus, a knockdown of these proteins restored significantly the expression of TLR9 in MKL1 (MCPyV-positive MCC line) (66). Keratinocytes transduced with the early genes of MCPyV and stimulated with a CpG ODN failed to secrete IL-6, IL-8, and MIP3α, as normal keratinocytes do. In contradistinction to EBV, MCPyV did not abolish TLR9 expression through the NFκB signaling pathway, but inhibited the transcription of TLR9 by downregulating the C/EBPα and C/EBPβ proteins (66). Moreover, an evaluation of TLR9 regulation by other members of the polyomavirus family showed that BK polyomavirus and JC polyomavirus efficiently diminish TLR9 expression, as shown in keratinocytes transduced with recombinant retroviruses expressing the early proteins LT and sT from each virus (66).
As observed in HPV and HBV infection, MCPyV proteins decrease TLR9 expression; however, based on the evidence discussed herein, it is likely that TLR9 expression is augmented in tumor cells of Merkel cell carcinoma as a result, at least in part, of the hypoxic conditions in solid tumors.
TLR9 in Cancer
The regulation of TLR9 expression has also been assessed in other cancer types, unrelated to oncogenic viruses. In this regard, two different TLR9 expression profiles have been reported in cancer, probably due to differences in variable stratification for statistical analysis. For instance, TLR9 serum levels were measured in 60 cases of breast cancer and compared with the same number of samples from benign breast disease patients and healthy controls in an Indian population (34); the results showed a reduction in TLR9 levels as the severity of the disease increased. Similarly, a higher tumor TLR9 expression in patients with renal cell carcinoma (Finnish population, n = 138) was associated with a better prognosis with respect to patients with lower TLR9 expression levels, as evaluated by immunohistochemistry (60).
On the other hand, a study in an Austrian population reported that TLR9 expression in breast tumor correlates with the estrogen receptor (ER) protein expression status. Although no significant differences were found, there was a trend suggesting that TLR9 mRNA expression was higher in ER-negative samples from breast cancer patients (n = 44) in comparison with ER-positive samples from breast cancer patients (n = 79) and healthy controls (n = 10) (5). A differential evaluation of TLR9 by breast cancer stage showed higher TLR9 mRNA levels in samples from patients with grade III tumor (n = 24) with respect to healthy controls (n = 10). A similar trend was found in ovarian cancer patients, showing that TLR9 expression was higher in stage III ovarian tumor samples (n = 56) in comparison with nonneoplastic samples (n = 30) (5).
TLR9 evaluation in samples from prostate cancer patients and samples from benign prostate hyperplasia patients (control) showed that TLR9 was significantly higher in prostate cancer patients with respect to controls (Finnish population) (75). Even though no TLR9 was detected in stromal cells in samples from benign prostate hyperplasia patients, it was detected in stromal cells in samples from prostate cancer patients (75). TLR9 stratification into “low expression” (n = 47) and “high expression” (n = 22) in tumor samples from prostate cancer patients in a Chinese population (47) showed that subjects with high TLR9 protein expression have a higher probability of lymph node metastasis and a poorer prognosis.
In addition, TLR9 is overexpressed in tumors from patients with glioma (Chinese population) (79). In this work, TLR9 evaluation by immunohistochemistry in normal samples showed a lower degree of TLR9 immunoreactivity in normal brain tissue (n = 13), but TLR9 protein levels were significantly augmented in high-grade glioma tumor tissue (n = 128). Even more, a higher mean progression-free survival (from the date of surgery to the first MRI-confirmed recurrence or death) was related to a lower TLR9 protein expression (n = 38) with respect to patients with higher levels of TLR9 expression (n = 31) (79).
Finally, a study assessed serum TLR9 protein level in 76 Chinese patients with noncell small lung cancer (NCSLC), showing significantly higher TLR9 levels in serum from patients with NCSLC with respect to 50 healthy donors (38). As mentioned before, taking into account that changes in TLR9 expression could be detectable in PBMCs or serum in cancer patients, these works suggest that TLR9 could be used as a biomarker for the progression of many types of cancer.
TLR9, Cell Stress and Invasion
The role of TLR9 in cell transformation is not well understood, and remains to be elucidated whether TLR9 itself promotes cell transformation or TLR9 upregulation in cancer is a consequence of transformation instead; available evidence suggests both scenarios; some works have correlated the upregulation of TLR9 caused by cellular stress and many others have shown that TLR9 signals enhance proliferation and tumor cell invasion.
With regard to cell stress, experiments performed in mice showed that chronic physical stress augments TLR9 expression in macrophages, leading to an increased production of the IL-1β, IL-10, and IL-17 cytokines in plasma (82). Even more, TLR9 induces enhanced corticosterone (stress hormone) levels with respect to TLR9 knockout mice. In the same system, TLR9 induces the expression of TNF, IL-4, IL-6, and MCP-1 in splenocytic lymphocytes stimulated with concanavalin A (43). Thus, chronic stress could be associated with immunosuppression, imbalance of cytokine production, and cancer development in a TLR9-mediated way. In addition, DNA damage and p53 expression regulate TLR9; in particular, chemotherapy agents such as doxorubicin and 5-fluorouracil, ionizing radiation, and ultraviolet light induce TLR9 expression in cancer cells in a p53-dependent manner (51,67). Also, treatment of cancer cells with DNA derived from chemotherapy-killed cancer cells promotes proliferation and invasion (73).
Furthermore, Kirillov et al. showed that a prolonged stimulation to normal human fibroblasts (NHF) with TGF-β induces TLR9 expression. Also, the authors found that a long CpG stimulus to NHF-induced higher matrix metalloproteinase-14 (MMP14) expression levels and an increased capacity of invasion with respect to nonstimulated NHFs. Interestingly, CpG-stimulated NHFs had a higher survival capacity under hypoxic conditions (37).
TLR9 expression is induced by hypoxia in D54MG and U373MG glioblastoma cell lines, and this event correlates with an increased invasive capacity. Moreover, treatment with a TLR9 siRNA diminishes the invasive capacity of D54MG and U373MG cells, as well as the expression of MMP2 and MMP9 (62).
Glioma cell lines and tumor cell samples showed higher proliferation and invasion capability in response to CpG ODN (79). On the other hand, an increase in TLR9 levels correlated with glioma progression and diminished patient progression-free survival (79). Stimulation of breast cancer cells (MDA-MB-231) with CpG ODN increases its invasive capacity in a dose-dependent manner, by MMP13 activation; interestingly, non-CpG ODN induced invasion almost in the same extent as CpG ODNs (52).
Results by Tanaka et al. evidenced the expression of TLR9 in the surface of HCC cell lines and suggested an increase in cell proliferation when cells were stimulated with ODN CpG (72). Also, TLR9 was detectable on the cell surface of colon adenocarcinoma samples, and stimulation of the SW480 colon adenocarcinoma cell line with CpG ODN increased the viability of these cells (54).
Taken together, these works showed that TLR9 is activated by mitochondrial, tumor-derived, and self DNA; hence, DNA recognition by TLR9 is not restricted to viral or bacterial genetic material. Furthermore, TLR9 upregulation was associated with cellular stress and its activation promotes cell proliferation and invasion (Fig. 1).
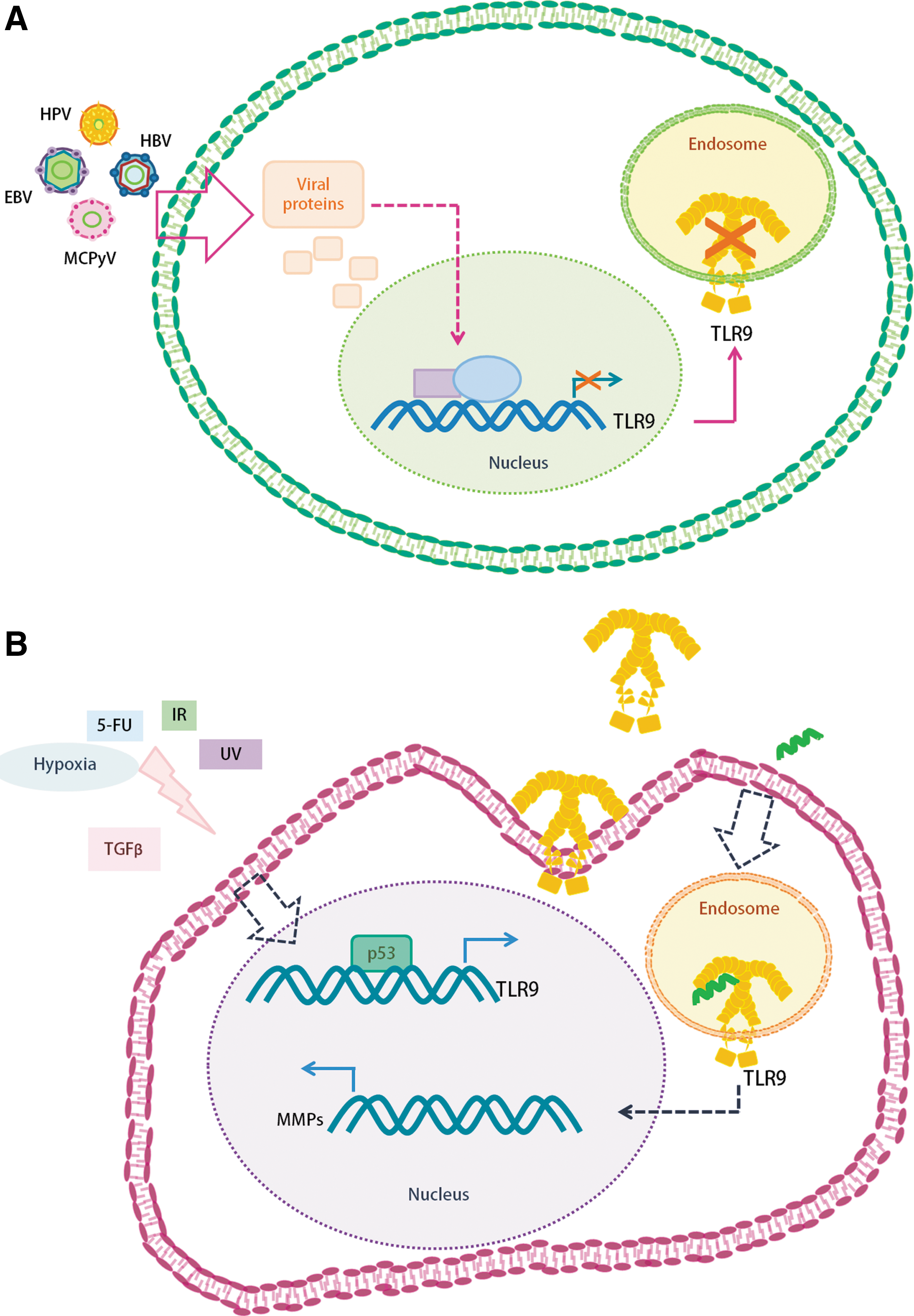
DNA oncogenic viruses show different strategies to inhibit TLR9 expression, probably as a mechanism to evade the immune response
Conclusions
It is well accepted that TLR9 is a TLR that recognizes DNA and has a crucial role in initiating and enforcing the immune response, and therefore many oncogenic DNA viruses have developed strategies to inhibit its expression. Nevertheless, TLR9 is associated with other cellular processes such as cell stress, proliferation, and invasion. In addition, TLR9 is involved in cellular processes not mentioned herein, like cell energy modulation and autophagy, and it is also related to cancer progression. TLR9 regulation depends on many factors and it is not known whether TLR9 expression is related to cancer as a cause or as a consequence, and probably both cases are true. Disregarding the multiple roles of TLR9, the evaluation of this protein in precancerous lesions, in PBMCs, and in plasma could represent a new marker for cancer progression. Certainly, an extensive research in this regard remains to be done.
Footnotes
Author Disclosure Statement
No competing financial interests exist.