
Abstract
Porcine reproductive and respiratory syndrome virus (PRRSV) is an economically important global swine pathogen. PRRSV infects porcine dendritic cells (DCs), but the effects of the interactions with DCs are largely unknown. Current research focuses on the production and regulation of interferons and selected inflammatory cytokines in DCs, which may play key roles in immune modulation. In addition, PRRSV also downregulates swine leukocyte antigen class I (SLA-I), SLA-II, and CD80/86 costimulatory molecules in DCs. In this study, we aim to evaluate the PRRSV immunomodulatory effects on monocyte-derived DCs (MoDCs) through interactions with porcine DC-SIGN (pDC-SIGN) receptor. We demonstrated that blocking the PRRSV and pDC-SIGN interactions in MoDCs with recombinant hICAM-3 did not affect the regulatory effects of PRRSV on SLA-I, SLA-II, or CD80/86 molecules. The hICAM-3 did not affect the morphological changes on MoDCs associated with their activation and maturation after PRRSV infection, and did not impair the virus infectivity in these cells either. The mRNA levels of tumor necrosis factor alpha (TNF-α), IL-12p35, IL-1β, and IL-6 were upregulated after hICAM-3 treatment or PRRSV infection, but in the presence of the blockage of pDC-SIGN in MoDCs with hICAM-3, PRRSV did not modulate the expression of these genes. However, in the presence of an anti-pDC-SIGN monoclonal antibody (mAb), we showed that PRRSV infection significantly reduced the mRNA expression levels of TNF-α and IL-1α, but enhanced the expression of IL-12p35 in MoDCs. Both hICAM-3-Fc and pDC-SIGN mAb treatments did not modulate proinflammatory cytokine protein levels in the culture supernatants of PRRSV-infected MoDCs. The results indicate that blocking the PRRSV-pDC-SIGN interactions by recombinant hICAM-3-Fc did not significantly affect virus infectivity, DC maturation, and proinflammatory cytokine gene expression in infected MoDCs. However, blocking the PRRSV-pDC-SIGN interactions on MoDCs with an anti-pDC-SIGN mAb revealed differential regulatory effects on specific proinflammatory gene expressions in those cells.
Introduction
P
DCs possess a large repertoire of pathogen recognition receptors, including Toll-like receptors (TLRs) and C-type lectin receptors (CLRs), both of which recognize the molecular patterns expressed by pathogens such as PRRSV (34). DC-specific intercellular-adhesion-molecule-3 (ICAM-3)-grabbing nonintegrin (DC-SIGN, CD 209) is a type II transmembrane protein that mediates DC differentiation, migration, and pathogen internalization, and plays an important role in the immune regulation of DCs (9,38). Human DC-SIGN contains a carbohydrate-recognition domain (CRD), a neck region composed of seven and half repeats containing 23 amino acid residues, a transmembrane region, and a cytoplasmic tail (8). Signaling through the DC–SIGN receptor includes downstream mediators in the Raf-1 pathway that leads to the production of proinflammatory cytokines IL-12, IL-6, TNF-α, and immunosuppressive cytokine IL-10. Certain pathogens can modulate T helper type 1 (Th1) polarization, Th2 response, and/or the induction of regulatory T cells through DC-SIGN binding. It has been demonstrated that mannose-rich pathogens enhance the expression of IL-10, IL-12, and IL-6, while fucose-expressing pathogens enhance the expression of IL-10, but downregulate the expressions of IL-12 and IL-6 (32).
The role of PRRSV and DC interaction during PRRSV infection is largely unknown, and current research focuses on the production and regulation of IFNs and specific inflammatory cytokines in DCs, which may play a role in immune modulation after infection. Plasmacytoid DCs are considered to be key effectors that express high levels of type I IFNs in the early stages of antiviral innate immunity (11). In vitro experiments with bone marrow-derived immature DCs and mature DCs (mDCs) derived from porcine monocytes exposed to PRRSV demonstrated an upregulation of CD80/86 and an increased secretion of IL-10 (6,10), but no effect was observed on IFN-α, IL-6, IL-8, IL-12, IFN-γ, or TNF-α secretion (2). Also, in vitro cultured MoDCs infected with PRRSV showed an increase in IL-10, IL-12, and TNF-α production (25). Whether PRRSV and DC interactions occur through specific receptor(s) is unknown, however, our previous studies have shown that PRRSV interacts with pDC-SIGN, possibly through certain N-glycans in structural proteins, and enhances PRRSV transmission in vitro (18).
In this study, we evaluated the immunomodulation of MoDCs by PRRSV through the pDC-SIGN receptor. We employed approaches such as blocking the pDC-SIGN receptor either through recombinant hICAM-3-Fc or anti-pDC-SIGN monoclonal antibody (mAb) before PRRSV infection. We measured the proinflammatory gene expressions in PRRSV-infected MoDCs with disrupted PRRSV-pDC-SIGN interactions. We demonstrated that disrupting the PRRSV-pDC-SIGN interactions on MoDCs by competitive anti-pDC-SIGN mAbs reduced the mRNA expression of TNF-α and IL-1α, but enhanced the expression of IL-12 induced by PRRSV in those cells.
Materials and Methods
Preparation of porcine MoDCs
Fresh heparinized venous blood was obtained by jugular venipuncture from cross-bred Yorkshire-Landrace grower pigs housed at Virginia Tech Swine Research Center. Pigs were tested negative for the presence of PRRSV and PCV2 antibodies and PRRSV RNA and PCV2 DNA in serum samples. MoDCs were generated essentially as previously described (4,24). Briefly, peripheral blood mononuclear cells (PBMCs) were isolated from blood by density-gradient centrifugation at 400 g for 40 min at room temperature over Ficoll-paque PREMIUM (1.077 g/L) (GE Healthcare). The PBMCs were washed twice with PBS at 250 g for 10 min at room temperature. After washing, the cells were suspended in Iscove's Modified Dulbecco's Medium (IMDM) supplemented with 10% fetal bovine serum (FBS) and 1% antibiotic/antimycotic (10,000 units/mL of penicillin, 10,000 μg/mL of streptomycin, and 25 μg/mL of Gibco Amphotericin B) (Thermo Fisher Scientific, Inc.). Monocytes were harvested by plastic adhesion overnight. After overnight incubation, cells were washed vigorously with PBS and cultured in IMDM supplemented with 10% FBS, 20 ng/mL of recombinant porcine granulocyte-macrophage colony-stimulating factor (Gibco), and 20 ng/mL of recombinant porcine IL-4 (Invitrogen) for 5 days. On day 3, half of the cell culture supernatant was replaced with a fresh medium supplemented with the aforementioned cytokines. On day 5, MoDCs were harvested with a cell dissociation buffer (Invitrogen) and scraped off the plate gently. The cells were washed with IMDM once, counted after Trypan blue staining, and plated with fresh IMDM supplemented with 10% FBS.
Flow cytometry analysis of surface markers
The effect of PRRSV infection on the maturation status of MoDCs in the presence or absence of the pDC-SIGN receptor blockade due to competitive hICAM-3 interaction was analyzed by flow cytometry (FCM) analysis of the surface expressions of swine leukocyte antigen class I (SLA-I), SLA-II, and CD80/86. As a positive control, cells were treated with lipopolysaccharide (LPS) (1 μg/mL). MoDCs infected with one multiplicity of infection (MOI) of PRRSV strain VR2385 with and without hICAM-3 treatment, as well as sham-inoculated and LPS controls were incubated for 18 h at 37°C in 5% CO2. Cells were washed with the FCM buffer (calcium- and magnesium-free PBS pH 7.4, 2% FBS). Viable cells were counted after staining with Trypan blue (0.4%) and resuspended in the FCM buffer at a concentration of 1 × 106 cells/mL. Cells (100 μL) were incubated at 4°C with primary antibodies for pig anti-SLA-II or anti-SLA-I (Monoclonal antibody center, Washington State University) or human CD152-muIgG2aFc (Ancell corporation) was used to stain pig CD80/86 at a concentration of 15 μg/mL for 45 min. Cells were washed twice with 1 mL FCM buffer by centrifugation at 250 g for 5 min and incubated with goat anti-mouse IgG conjugated with Alexa fluor 488 (Invitrogen) at a dilution of 1:200 for 30 min in the dark. Subsequently, cells were washed twice with 1 mL FCM buffer by centrifugation. Finally, cells were resuspended in the FCM buffer, and the data were acquired with a FACScan flow cytometer (BD Biosciences) and analyzed using FlowJo software. Dead cells were excluded from the analysis by DAPI staining.
Blocking pDC-SIGN and PRRSV interactions in MoDCs through recombinant hICAM-3-Fc or anti-pDC-SIGN mAb
The hICAM-3-Fc binds with porcine DC-SIGN with high affinity comparable with human DC-SIGN, which is competitively inhibited by PRRSV virion interaction with porcine DC-SIGN (18), suggesting that the binding sites of hICAM-3 and PRRSV on porcine DC-SIGN are overlapped. Therefore, PRRSV-pDC-SIGN interaction can be competitively inhibited by pre-treatment with hICAM-3-Fc. A total of 2 × 105 MoDCs were suspended in 100 μL IMDM that was supplemented with 10% (v/v) FBS and incubated for 30 min at 4°C with 1 μg recombinant hICAM3-Fc (R&D systems). Cells were washed twice with 500 μL IMDM by centrifugation at 250 g for 10 min at 4°C. After washings, one group was resuspended in 100 μL IMDM (ICAM-3/M) and another group was infected with one MOI of PRRSV strain VR2385 for 2 h at 4°C (ICAM-3/V). The negative-control group was treated only with IMDM (M/M), and the positive-control group was infected with the same dose of PRRS without hICAM-3 treatment (V/M).
For the anti-pDC-SIGN-specific blockage assay, a total of 2 × 105 MoDCs were suspended in 100 μL IMDM supplemented with 10% (v/v) FBS and incubated for 30 min at 4°C with various concentrations of an anti-pDC-SIGN mAb (DC131, developed in our laboratory), which is specific to the carbohydrate-recognition domain (CRD) of pDC-SIGN. Cells were washed twice with 100 μL IMDM by centrifugation at 250 g for 10 min at 4°C. After washings, one group was resuspended in 100 μL IMDM (mAb/M) and a second group was infected with one MOI PRRSV strain VR2385 for 2 h at 4°C (mAb/V). A negative-control group was treated with IMDM (M/M), and a positive-control group was infected with the same dose and virus strain, but without the DC-SIGN-mAb treatment (M/V). After treatments, the cells were plated in a 24-well plate and incubated at 37°C in 5% CO2 and one well per treatment was collected every 6 h for a period of 24 hour postinfection (hpi). Cell culture supernatants were aliquoted and stored at −80°C for the quantification of virus and cytokine levels. Cells were scraped off the well and resuspended in 300 μL Trizol (Invitrogen) for further mRNA quantification by quantitative polymerase chain reaction (PCR). Total RNAs were also extracted from the harvested MoDCs using an RNeasy Micro Kit (Qiagen) according to the manufacturer's instructions. RNAs were reverse transcribed using the iScript cDNA synthesis kit (Bio-Rad) and random primers. The following cycling conditions were used to synthesize the cDNAs: one cycle at 25°C for 5 min, one cycle at 42°C for 30 min, and one cycle at 85°C for 5 min.
Quantitative real-time PCR and cytokine ELISA to quantify the mRNA expression and secretion levels of cytokines
The relative quantification of gene expressions of TNF-α, IL-12p35, IL-1α, IL-1β, and IL-6 was performed with a Bio Rad CFX96 detection system (Bio Rad). Reactions were performed in duplicate, with cyclophilin A (CYPA) being used as the internal control. All primers were custom synthesized (Integrated DNA Technologies, Inc.) to target amplicons with sizes ranging from 160 to 190 nt, based on the cDNA sequence of each gene of interest collected from the NCBI database (Table 1). The mRNA expression levels were quantified according to the ΔΔCt method (20). A 15 μL reaction mixture, containing 12.5 ng cDNA, 7.5 μL SYBR Green PCR Master Mix (Bioline's Sensimix SYBR & Fluorescein Kit), and 7.5 picomoles of each primer, was assembled. The following thermal cycle conditions were used: an initial activation step at 95°C for 10 min, followed by a 3-step PCR program of 95°C for 15 sec, 55°C–60°C for 60 sec, and 72°C for 60 sec for 40 cycles. A dissociation curve was obtained for each quantitative PCR run to assess the target specificity of the PCR run.
IL, interleukin; TNF, tumor necrosis factor; CYPA, cyclophilin A; PCR, polymerase chain reaction.
The cytokine protein secretion levels in the cell culture supernatants were measured using commercially available multiplex ELISA MILLIPLEX MAP Porcine Cytokine and Chemokine Magnetic Bead Panel—Immunology Multiplex Assay (PCYTMAG-23), which was customized for the detection of TNF-α, IL-12p35, IL-1α, IL-1β, and IL-6 (EMD, Millipore) with standard internal controls. The reporter fluorescence of the beads was determined by using a dual-laser Bio-Rad Bio-Plex 200 instrument with Bio-Plex Manager software Version 6.0 (Bio-Rad) and expressed as the median fluorescent intensity of ≥50 microspheres per set per well.
Virus titration in the supernatant was measured by immunofluorescence assay using anti-PRRSV N-specific antibody (SDOW17) (RTI, Brookings, SD), followed by fluorescein isothiocyanate-conjugated goat anti-mouse IgG (KPL, Kirkegaard & Perry Laboratories, Inc.), and detected with a fluorescence microscope within excitation and emission spectrum peak wavelengths of ∼495 and 519 nm, respectively. The tissue culture infectivity dose 50 (TCID50) was calculated using the Reed and Muench method (27).
Statistical analysis
The significance between treatment groups was determined by two-way ANOVA with a Tukey HSD post-hoc test using Graph pad Prism v5 and Microsoft Excel. A p-value of less than 0.05 was considered to be statistically significant.
Results
MoDC susceptibility for PRRSV, activation and maturation are not affected by pDC-SIGN blockage by hICAM-3
We first investigated whether the blockage of pDC-SIGN with recombinant hICAM-3 affects PRRSV infectivity or the activation and maturation of pDCs. It has been reported that pDC-SIGN is not an entry receptor for PRRSV into pDCs, but it does enhance PRRSV transmission in vitro (18). DC differentiation is characterized by morphological changes characterized by the presence of large cytoplasmic projections of irregular shape with pseudopodia and the expressions of SLA-I, SLA-II, and the CD80/86 costimulatory molecules (1,19). In this study, we did not observe morphological difference between hICAM-3-blocked cells and mock-treated cells at 24 hpi with PRRSV (Fig. 1A, B). Both groups, ICAM-3/V and M/V, infected with one MOI PRRSV showed marked cytopathic effects (CPE) at 24 hpi. Morphological changes observed in infected cells were characterized by cell rounding, cytoplasmic swelling, and the loss of the characteristic cytoplasmic dendritic projection compared to the mock-infected cells (Fig. 1B).
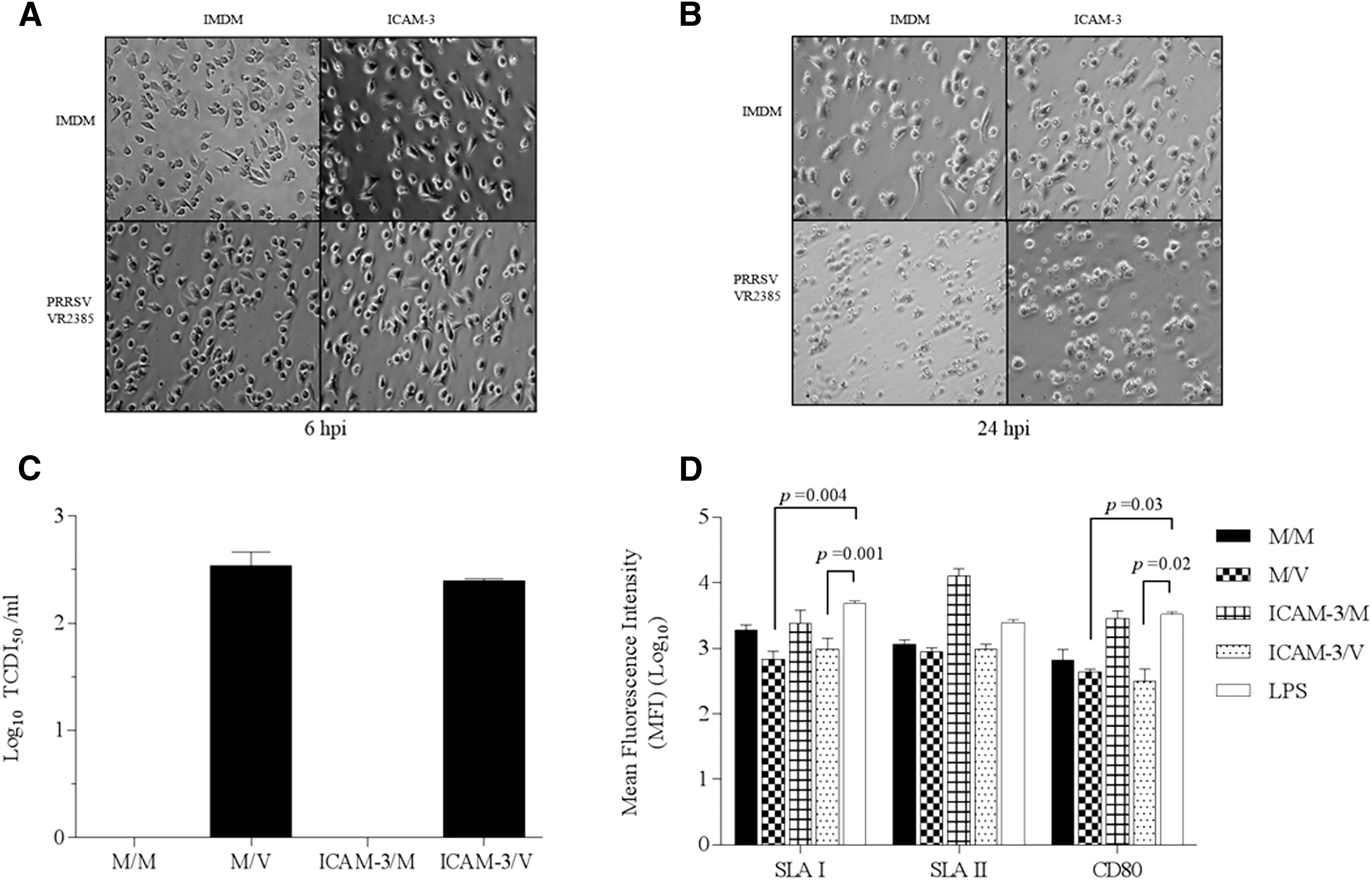
Effect of PRRSV infection after pDC-SIGN blockage with recombinant hICAM-3 on MoDC susceptibility, activation and maturation.
The infectivity of PRRSV was not affected, as there was no difference in TCID50 viral titers in the culture supernatants between M/V and ICAM-3/V groups (Fig. 1C). In this study, we also found that PRRSV infection significantly reduced the expression of SLA-I and costimulatory CD80 molecules (Fig. 1D), as previously reported (25). There was not a significant reduction of SLA-II in M/V and ICAM-3/V compared to the LPS control; however, PRRSV had a detrimental effect on SLA-II molecule expression. Therefore, the results indicate that the blockage of pDC-SIGN with recombinant hICAM-3 does not affect PRRSV CPE formation or infectivity, but downregulates the expression of SLA-I (but not class II) and CD 80/86 expression.
In vitro blockage of pDC-SIGN with hICAM-3 does not affect proinflammatory cytokine mRNA expression and protein secretion in MoDCs infected by PRRSV
Numerous studies have demonstrated that PRRSV has an effect on proinflammatory cytokine regulation in MoDC (2). However, the specific mechanisms underlying the modulation of proinflammatory cytokines in these cells are not well understood. To our knowledge, there is no information regarding the role of pDC-SIGN as a mediator in proinflammatory modulatory effects during PRRSV infection. A previous study in our laboratory demonstrated that pDC-SIGN enhances PRRSV transmission in trans and further showed that PRRSV interacts with porcine DC-SIGN, which could be blocked by treatment with recombinant hICAM-3 molecules (18). In this study, we measured the effect of pDC-SIGN blockage with hICAM-3 on proinflammatory cytokine gene expression and protein production in MoDCs infected with PRRSV.
No differences were observed in TNF-α, IL-12p35, IL-1α, IL-1β, or IL-6 mRNA expression levels at 6, 12, 18, and 24 hpi between the MEM-PRRSV (M/V) and hICAM-3-PRRSV (ICAM-3/V) treatments (Fig. 2). All genes under investigation in this study showed almost a two-log increase in the ICAM-3/V group at 24 hpi when compared to the mock-treated cells (M/M) (Fig. 2). All proinflammatory cytokines examined in this study, except for IL-1α, showed a marked mRNA upregulation in ICAM-3/V group compared to M/M group at 24 hpi. The upregulatory effect of PRRSV on the mRNA expression of TNF-α, IL-12p35, IL-1β, and IL-6 was not significantly enhanced in cells pretreated with hICAM-3 (Fig. 2A–E).
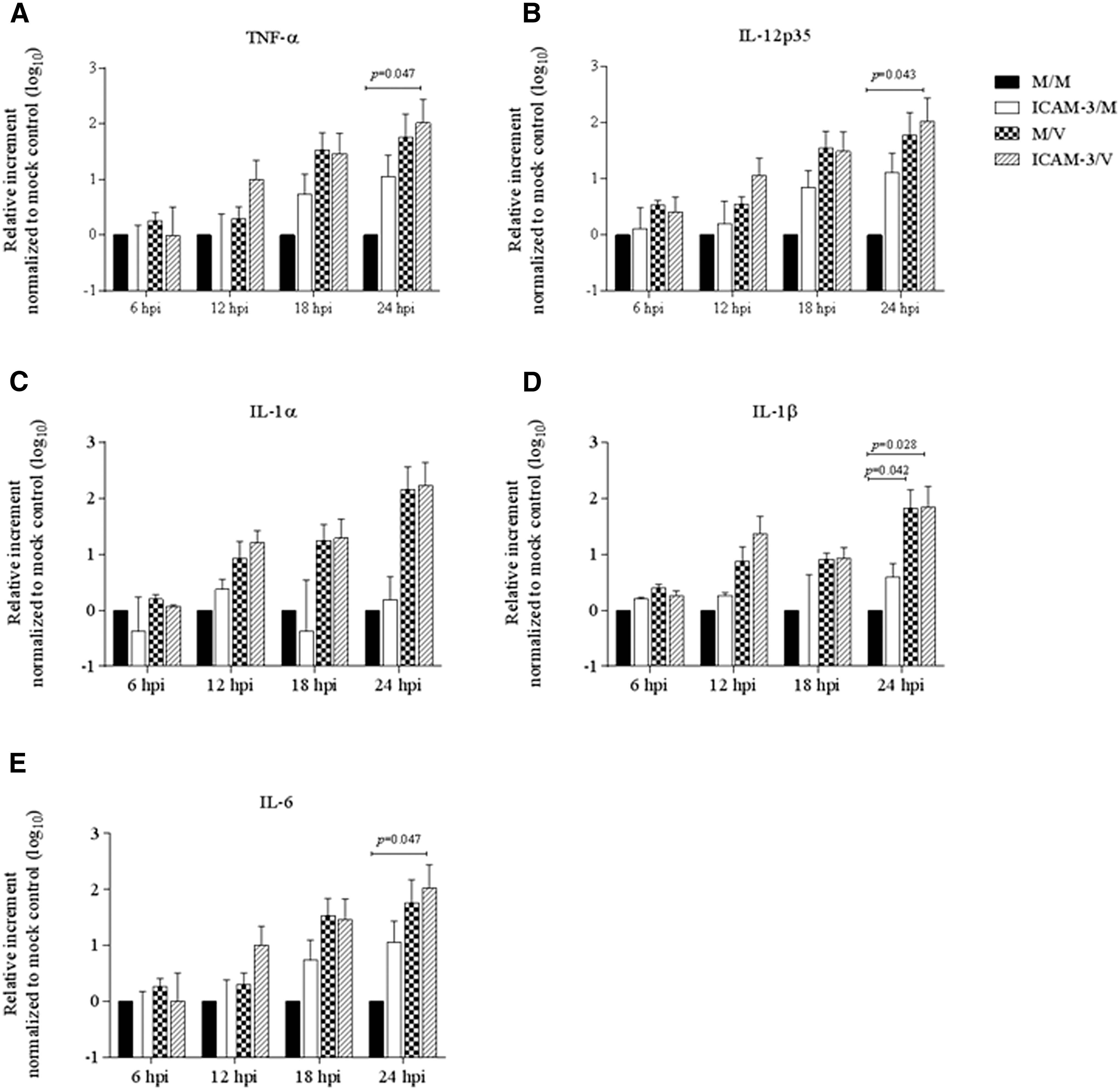
Regulatory effect of proinflammatory cytokine mRNA expression in MoDCs induced by PRRSV infection in the presence of pDC-SIGN blockage with hICAM-3.
The cytokine fluorescence intensities were evaluated in MoDC culture supernatants at 6, 12, 18, and 24 hpi. The results obtained for M/V, ICAM-3/M, ICAM-3/V, and mock-infected cells (M/M) were compared. There was no significant difference in TNF-α, IL-12p35, IL-1α, IL-1β, and IL-6 cytokine protein levels between treatment groups throughout the experiment. The intensities of TNF-α and IL-12p35 did not show a significant difference among the various treatments and also remained constant during the four time points evaluated in this experiment (Fig. 3A, B). Although there was not a significant difference among treatment groups at any time point, the IL-1α, IL-1β, and IL-6 proteins showed similar secretion patterns, which were characterized by a slight, constant, measurable increase in protein concentration through the four time points in all treatment groups (Fig. 3C–E). These results indicate that blocking pDC-SIGN-PRRSV interactions with hICAM-3 molecule does not affect the modulation of proinflammatory cytokines in MoDCs infected with PRRSV.
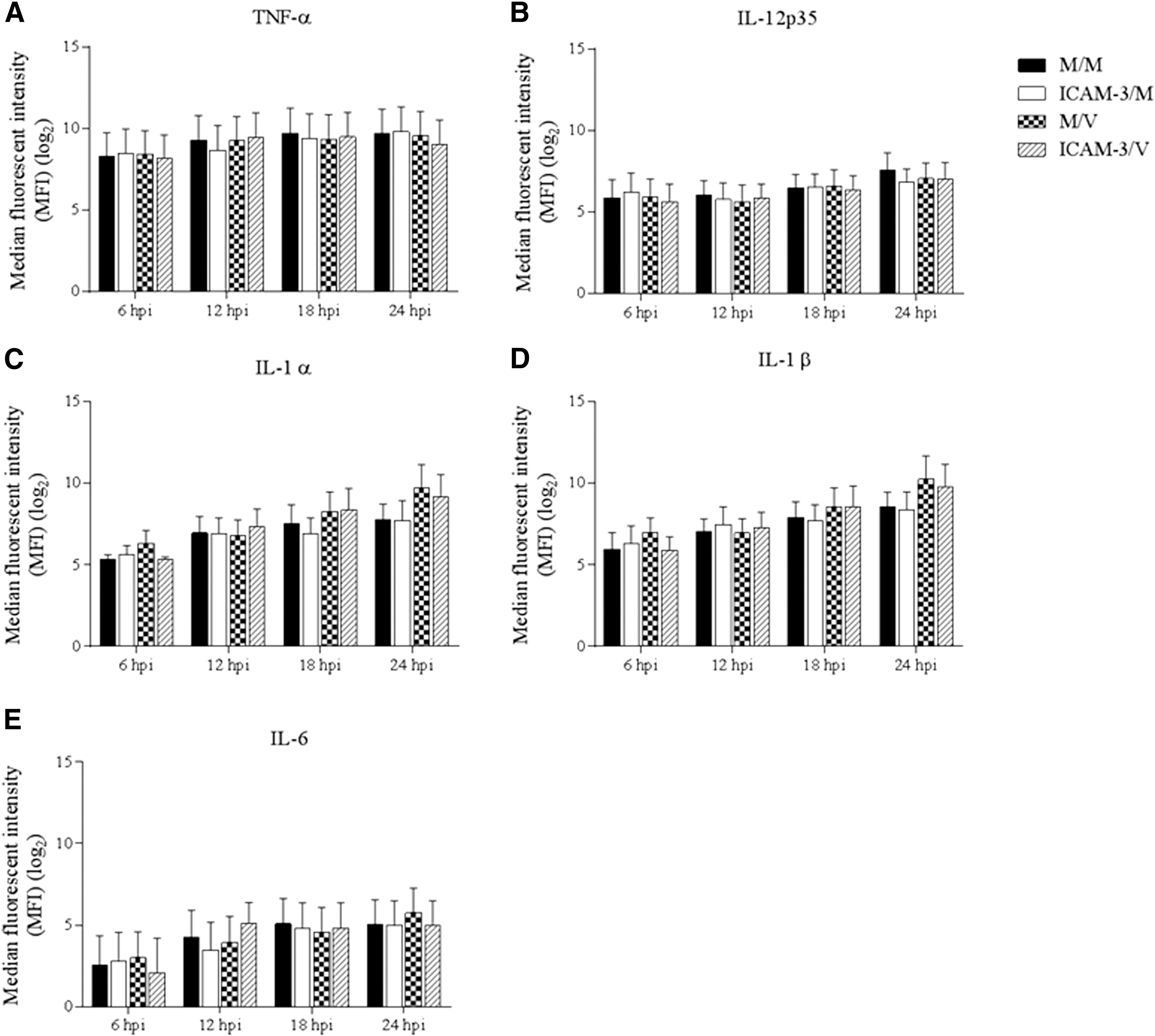
Cytokine protein production of MoDCs infected with PRRSV in the presence of pDC-SIGN blockage with hICAM-3.
In vitro blockage of pDC-SIGN with an anti-pDC-SIGN mAb differentially modulated proinflammatory cytokine mRNA expression, but does not affect the corresponding protein levels in culture supernatants of MoDCs infected with PRRSV
To evaluate the specific effect of PRRSV infection on proinflammatory cytokine gene regulation through the pDC-SIGN receptor, we utilized a different approach in which MoDCs were treated with an anti-pDC-SIGN mAb (DC131) before PRRSV infection (mAb/V). No statistical difference in proinflammatory cytokine mRNA modulation was observed between MoDCs infected with PRRSV (M/V) and MoDCs pretreated with anti-pDC-SIGN mAbs (mAb/M) at 6, 12, and 18 hpi. At 24 hpi, however, there was a significant reduction in TNF-α as well as IL-1α mRNA levels, but an increase in IL-12 mRNA levels in anti-pDC-SIGN mAb-pretreated PRRSV-infected MoDCs (mAb/V) when compared to PRRSV-infected MoDC control (M/V) (Fig. 4A–E).
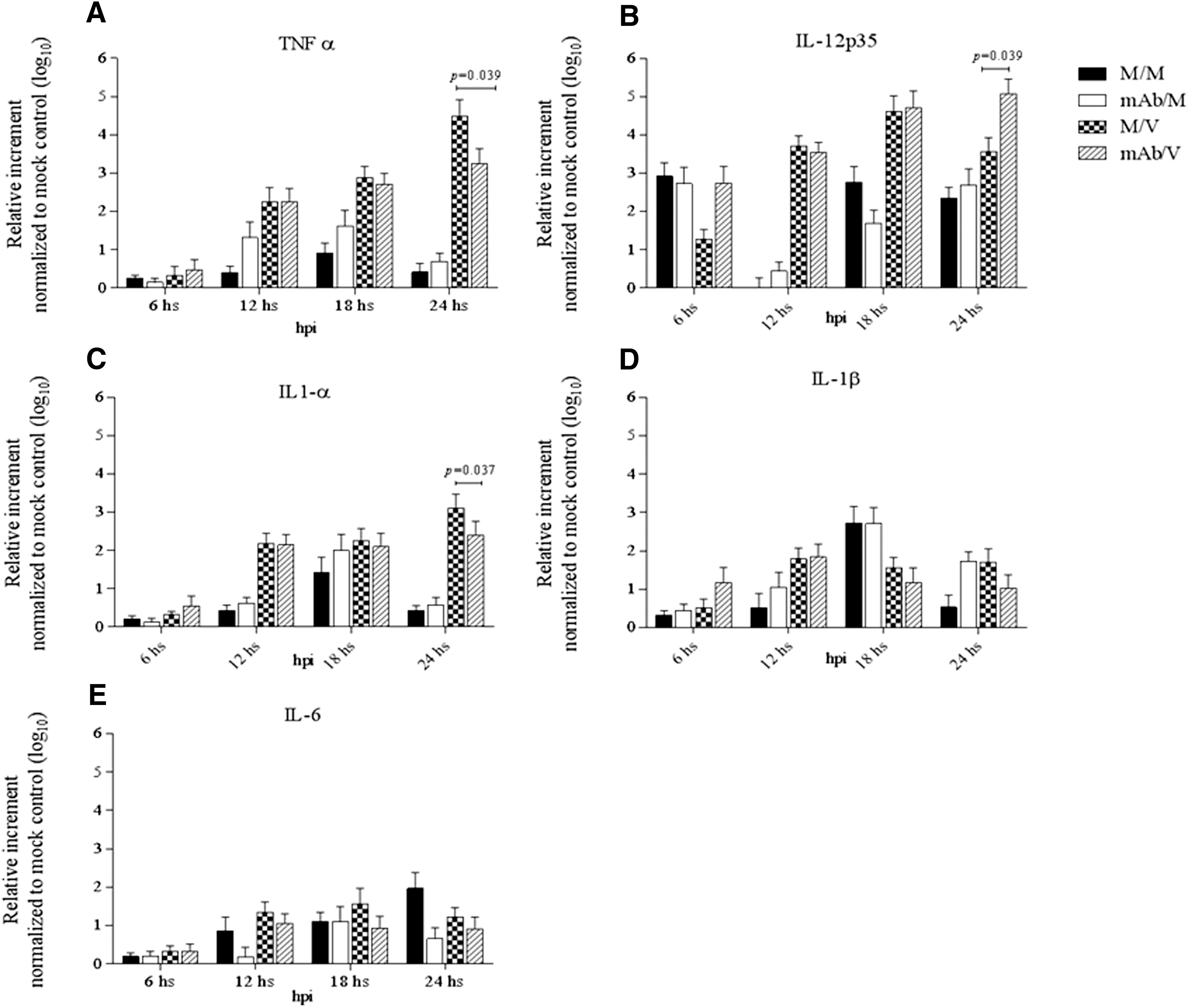
Regulatory effect of proinflammatory cytokine mRNA expressions in MoDCs induced by PRRSV infection in the presence of pDC-SIGN blockage with anti-pDC-SIGN mAb.
The effect of PRRSV infection on cytokine protein production after DC-SIGN blockage with anti-pDC-SIGN mAb was evaluated in MoDC culture supernatants at 6, 12, 18, and 24 hpi. No significant differences in TNF-α, IL-12p35, IL-1α, IL-1β, and IL-6 cytokine protein levels were observed between the M/V, mAb/V, and M/M groups throughout the experiment. Although the differences were not statistically significant, there was a trend of lower concentrations for all cytokines evaluated at 24 hpi in the mAb/V treatment group compared to the M/V positive-control group (Fig. 5A–E).
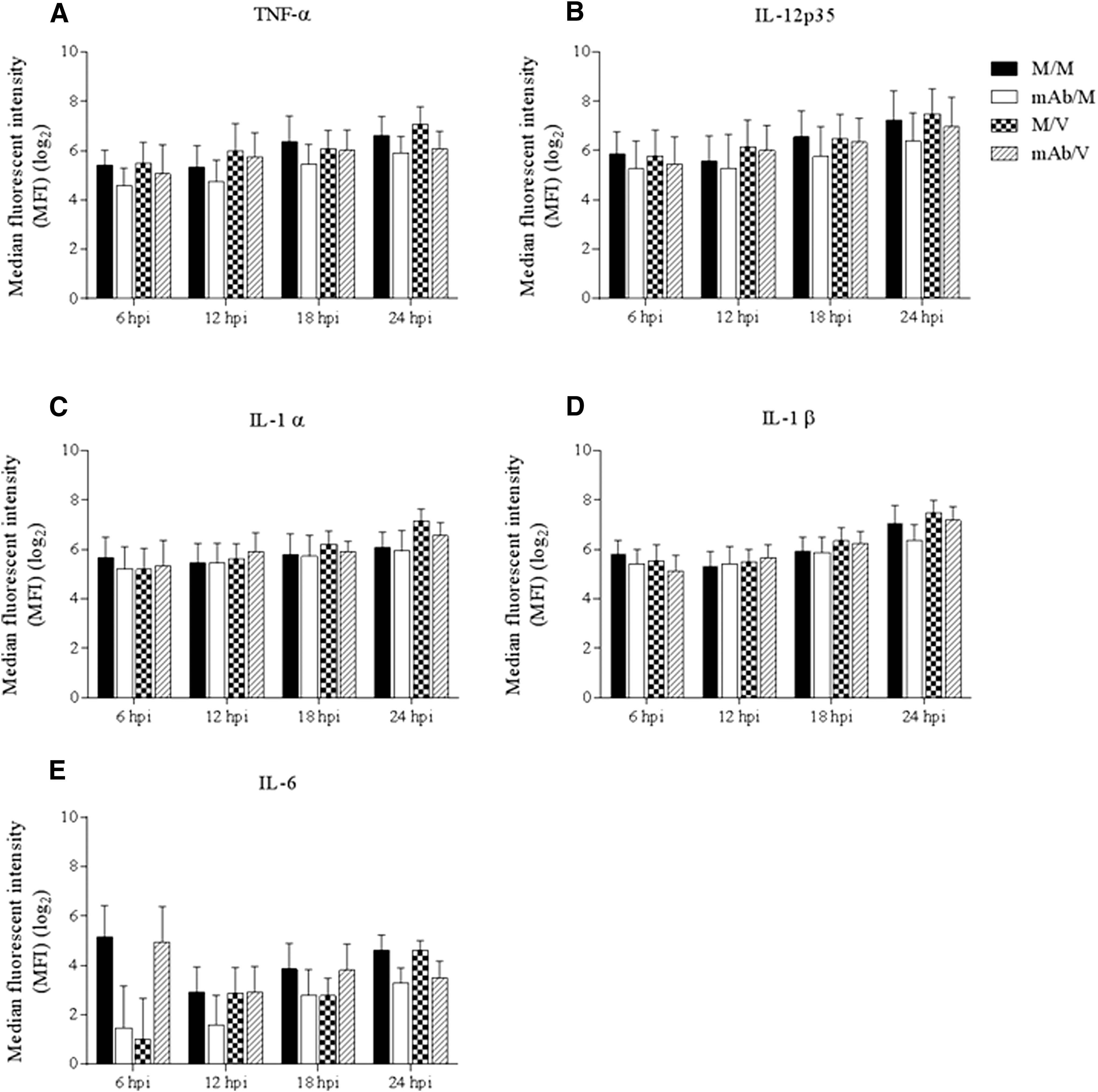
Cytokine production of MoDCs infected with PRRSV in the presence of pDC-SIGN blockage with anti-pDC-SIGN mAb.
Discussion
Evasion of host immune responses by immunosuppression and establishment of a persistent infection is one of the mechanisms of PRRSV pathogenesis. The key mechanism that PRRSV induces immunosuppression is probably through the modulation of numerous cytokines such as type I IFNs, TNF-α, IL-1, IL-6, and IL-12p35 in infected pigs (5,30,33). In this study, we evaluated the role of pDC-SIGN in cytokine gene expression and protein production in MoDCs experimentally infected with PRRSV in vitro. Our data suggest that pretreating MoDCs with an anti-pDC-SIGN mAb effectively reduced the ability of PRRSV to induce TNF-α and IL1-α gene transcription, but increased its ability to induce IL-12p35 gene transcription in the infected cells, even though the anti-pDC-SIGN mAb does not have an effect on the protein synthesis and secretion of the same cytokines.
Productive infection of porcine MoDCs by PRRSV has previously been reported (36). The CD163 and sialoadhesin molecules are responsible for PRRSV entry and infectivity. However, heparin sulfate, CD151, and vimentin have also been identified as potential PRRSV receptors due to their involvement in virus attachment, internalization, or uncoating (29). CLRs are expressed on the DC surfaces and are associated with capturing pathogen-derived glycosylated proteins and internalizing them for efficient antigen presentation (9). Although it has been postulated that pDC-SIGN is not an entry receptor for PRRSV in pDCs, it does enhance viral transmission in vitro (18). In this study, we demonstrated that blockage of the pDC-SIGN receptor by a recombinant hICAM-3 did not affect PRRSV intake or infectivity, which is consistent with the previous findings that pDC-SIGN does not play a role in virus entry or infectivity.
DC activation and maturation are characterized by morphological changes and the expression of SLA-I, SLA-II, and CD80/86 costimulatory molecules (25). It has previously been reported that PRRSV infection downregulates SLA-II and CD80/86 in mDCs (10). Other studies found that PRRSV (strain CNV-3) downregulates SLA-I and SLA-II expression on infected MoDCs (25). In addition to the SLA-I and SLA-II downregulation observed due to PRRSV infection in DCs, the CD14 and CD11b/c have also been downregulated by PRRSV infection (36). We found that pDC-SIGN blockage in MoDCs with hICAM-3 did not affect the regulatory effect of PRRSV on SLA-I, SLA-II, or costimulatory molecules CD80/86, suggesting that pDC-SIGN does not play an exclusive role in PRRSV activation and maturation activities of MoDCs.
DCs play a pivotal role in establishing the adaptive immune responses and are crucial in determining the type of adaptive immune response. In addition to adhesive and antigen recognition properties, the engagement of DC-SIGN with cognate ligands or antibodies in DCs resulted in the activation of signal transduction pathways that can cause an extensive modulation of immune responses (14).
Infection with PRRSV induces a weak TNF-α production both in vivo and in vitro (21,35). Moreover, it has been demonstrated that, during the early stages of PRRSV infection, TNF-α mRNA expression levels do not differ between PRRSV-infected and mock-infected MoDCs and PRRSV may induce a late, observable upregulation between 12 and 24 hpi (31). Moreover, in vitro studies demonstrated that PRRSV induces IL-1 and IL-6 in bone marrow-derived DCs (26). However, in vivo studies have shown that IL1-β and IL-6 were not elevated by 10 dpi, which is coincident with the presence of acute clinical signs and pulmonary lesions in PRRSV cases, but these interleukins were elevated in the persistent stage at 28 dpi (33). We also observed a moderate increase in gene expression levels of TNF-α and other proinflammatory cytokines such as IL-1α, IL-1β, IL-12p35, and IL-6, particularly in late time points (18–24 h) of our experiments. Our results indicate that the anti-pDC-SIGN mAb (DC131) effectively blocked the effects of PRRSV on proinflammatory cytokine gene expression in MoDCs. Thus, pDC-SIGN mAb treatment abrogated PRRSV-mediated delayed upregulatory effect on TNF-α and IL-1α mRNA expression. Interestingly, it produced an opposite effect on IL-12p35 gene expression in MoDCs. The molecular mechanisms underlying these proinflammatory immunomodulations mediated by PRRSV need to be further investigated.
It has been postulated that PRRSV harbors posttranscriptional mechanisms to suppress TNF-α production at the translation or secretion levels (2,3,31). No differences were observed in the protein production of TNF-α and other proinflammatory cytokines evaluated in this study both in between treatments and in between time points. Our data are consistent with previous studies in which PRRSV failed to induce protein secretion of TNF-α, and our data further indicate that there is a strong blockage of protein production and/or secretion of other proinflammatory cytokines such as IL-1α, IL-1β, IL-12, and IL-6 by PRRSV in MoDCs. The absence of differences in the proinflammatory cytokine production among the different treatments and time points evaluated with this experimental design is in agreement with previous reports (13). Through pDC-SIGN, PRRSV modulates the proinflammatory cytokines at the gene expression level rather than at protein synthesis and secretion level. Additional viral mechanisms independent of pDC-SIGN interactions may exist to completely suppress the protein production of proinflammatory cytokines investigated in this study.
In this study, the differential modulation observed among various cytokines due to pDC-SIGN blockage through hICAM-3 or anti-pDC-SIGN mAb may be the result of the differential or incomplete modulation of pDC-SIGN functions by hICAM-3. Previously, it has been demonstrated that hICAM-3 causes an incomplete inhibition of DC-SIGN, resulting in a failure to inhibit human immunodeficiency virus transmission through pDC-SIGN (37). We believe that there is a similar mechanism of inefficient blockage of pDC-SIGN by hICAM-3, which failed to modulate the effects of PRRSV mediated through pDC-SIGN in our experiments.
In summary, we demonstrated that the interactions of PRRSV with pDC-SIGN modulated proinflammatory cytokine gene expression at transcription levels, particularly for TNF-α, IL-1α, and IL-12p35. A specific anti-pDC-SIGN mAb abrogates PRRSV's modulatory effect on the gene expression of above-said proinflammatory cytokines. Although the innate cytokine responses against PRRSV infection are regulated by a complex balance of negative and positive signals mediated through diverse TLRs and CLRs, in this study, we demonstrated a role for pDC-SIGN in the immunomodulation caused by PRRSV in porcine DCs.
Footnotes
Acknowledgments
This work is supported by internal funds from Virginia-Maryland College of Veterinary Medicine at Virginia Tech. We thank the FCM specialist Ms. Melissa Makris for her expert assistance. We also thank the technical support from Iowa State University College of Veterinary Medicine animal care staff.
Author Disclosure Statement
No competing financial interests exist.