
Abstract
Human enterovirus 71 (EV-A71) is one of the main etiological agents of hand, foot, and mouth disease (HFMD), which has been prevalent mainly in the Asia-Pacific region in the past several decades. The nonstructural proteins of EV-A71 will be expressed significantly during viral replication in host cells after EV-A71 infection. For the determination of the antibodies response against nonstructural proteins of EV-A71, in this study, the complete 2ABC, 3ABC, and 3D proteins were expressed in Escherichia coli and were then studied for their immunoreactivity by immunoblot assay and indirect enzyme-linked immunosorbent assay (ELISA), respectively. Three His-tagged fusion proteins were expressed effectively in E. coli, which were in agreement with the expected molecular mass. The results from immunoblot assay and indirect ELISA showed that all three purified fusion proteins can react with IgG antibodies from EV-A71-infected patients, but can hardly be recognized by IgG antibodies derived from mice or rabbits immunized by inactivated EV-A71 virus particles. The IgG antibody response against nonstructural proteins of EV-A71 is associated with viral infection or replication, which indicate that these nonstructural proteins could be used as candidate antigen for early diagnosis of EV-A71 infection, or to distinguish the EV-A71-specific antibodies after viral infection from inactivated vaccine immunization.
Introduction
E
EV-A71 is a small, single-stranded, positive-sense RNA virus belonging to the genus Enterovirus within the family Picornaviridae. Similar to other picornaviruses, genomic RNA of EV-A71 encodes a polyprotein precursor, which can be promptly cleaved by the viral proteases into at least 11 mature proteins, including four structural proteins (VP1–VP4) and seven nonstructural proteins (2A, 2B, 2C, 3A, 3B or VPg, 3C, and 3D) (23). The studies on the identification of antigenic epitopes mainly focus on structural proteins of EV-A71, especially VP1 (9,10,24). It was widely acknowledged that VP1 contains major specific or neutralizing epitopes of EV-A71. For nonstructural proteins, the majority of research currently focuses on its functions in viral replicate mechanisms or roles in viral-induced pathogenesis. However, the host immune responses against nonstructural proteins during EV-A71 infection, or identification epitopes of nonstructural proteins have not been investigated as yet. Therefore, this study utilizes recombinant nonstructural proteins expressed in Escherichia coli as antigens to investigate its immunoreactivity with antibodies against EV-A71 by immunoblot assay and enzyme-linked immunosorbent assay (ELISA), respectively. In addition, it was also investigated if differences existed in the antibodies response against nonstructural proteins between natural EV-A71 infection and inactivated vaccine immunization.
Materials and Methods
EV-A71 genes and antisera
A full-length cDNA clone of EV-A71 was constructed on the basis of the parental EV-A71 strain HeN09 (GenBank accession No. JN256064) (21). The serum specimens were prepared according to the methods described previously (28). Briefly, anti-EV-A71 animal sera were prepared by immunizing New Zealand white rabbits and BALB/c mice, respectively, with the heat-inactivated EV-A71 viron particles. All animals were housed in specific pathogen-free conditions and the studies were conducted according to the regulations of animal care and administration formulated by the State Science and Technology Commission of China. The human sera against EV71 were collected from hospitalized children (<3 years of age) of HFMD in Pediatrics, Shaoxing People's Hospital. Etiological diagnosis was made by RT-PCR assay using EV-A71 specific primers. Negative human sera were collected from healthy children (<3 years of age), who were confirmed to have not been infected with EV-A71. Human sera collection was approved by the ethics committee of the Shaoxing People's Hospital, and written informed consent was obtained from each subject.
Construction of the expression plasmids
The cDNA clone of EV-A71 was used as a template to amplify the coding regions of complete VP1, 2ABC, 3ABC, and 3D proteins using the Expand High Fidelity PCR system (Roche). All forward primers were designed with the BamH I restriction site, and the reverse primers were designed with the Not I restriction site and a stop codon at the 5′ end (Table 1). After being digested with BamH I and Not I (NEB), the cloned fragments were separately inserted into the multiple cloning sites of pET28a vector (Novagen) to generate recombinant plasmids. At the N-terminus of the target peptides, a 6His-tag was added to form a fusion protein. These expression plasmids were individually verified by DNA sequencing analysis (Invitrogen).
All primers were designed based on the genome sequence of EV71 M188-1181F/HeN/CHN/2009 (GenBank accession No. JN256064). The number in the name of primer denotes the location of amino acid from N- or C- terminus of target protein.
The recognition sequences of restriction enzymes are marked in italics, and stop codons are marked with bold letters.
Expression and purification of recombinant proteins
Expression and purification of recombinant proteins was performed according to the procedures described previously (27). In short, E. coli BL21 (DE3) competent cells were transformed with the recombinant plasmids, and were then cultured in Luria Broth (LB) medium with 40 μg/mL kanamycin at 37°C. The fusion protein expression was induced with isopropyl β-D-1-thiogalactopyranoside (IPTG, 0.4 mM; 2 h) at 37°C when the OD600 of the culture reached around 0.6. The bacterial cells were collected by centrifugation at 8,000 g for 10 min at 4°C, and the targeted proteins were detected by sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE). Fusion proteins were purified using Ni-NTA agarose (Qiagen) under denaturing conditions.
SDS-PAGE, immunoblot analysis, and indirect ELISA
SDS-PAGE, immunoblot analysis, and indirect ELISA were performed according to the methods described previously (27,28). As the primary antibodies, appropriately diluted antisera from EV-A71-infected patients and mice/rabbits immunized by inactivated EV-A71 viron particles were used. Meanwhile, as the negative control, preimmune animal sera or healthy children sera were also included in each assay. Horseradish peroxidase-conjugated goat anti-human, anti-mouse, and anti-rabbit IgG (KPL) were used as secondary antibodies.
Results
Expression and purification of VP1 and three nonstructural proteins of EV-A71
Four PCR fragments comprising VP1, 2ABC, 3ABC, and 3D coding sequences of EV-A71 were separately amplified using PCR. The size of PCR products were ∼891, 1734, 873, and 1389 bp, respectively, which was in agreement with the expected size (Fig. 1). To construct recombinant prokaryotic expression vectors, these gene fragments were separately inserted into multiple cloning sites of pET28a. The DNA sequencing results showed that four protein-coding sequences inserted into plasmids were fully consistent with its original sequences. Four recombinant plasmids, including pET28a-VP1, pET28a-2ABC, pET28a-3ABC, and pET28a-3D, were transformed into E. coli BL21(DE3). These cloned transformants were subsequently induced with IPTG. The results of SDS-PAGE showed that four recombinant proteins can be effectively expressed and its sizes were ∼32, 64, 32, and 52 kDa, which were consistent with the expected molecular weight (Fig. 2). In addition, all four His-tag fusion proteins were fully expressed as inclusion bodies. After being solubilized in 8 M urea, these fusion proteins were purified under denaturing conditions using Ni-NTA (Fig. 3a).
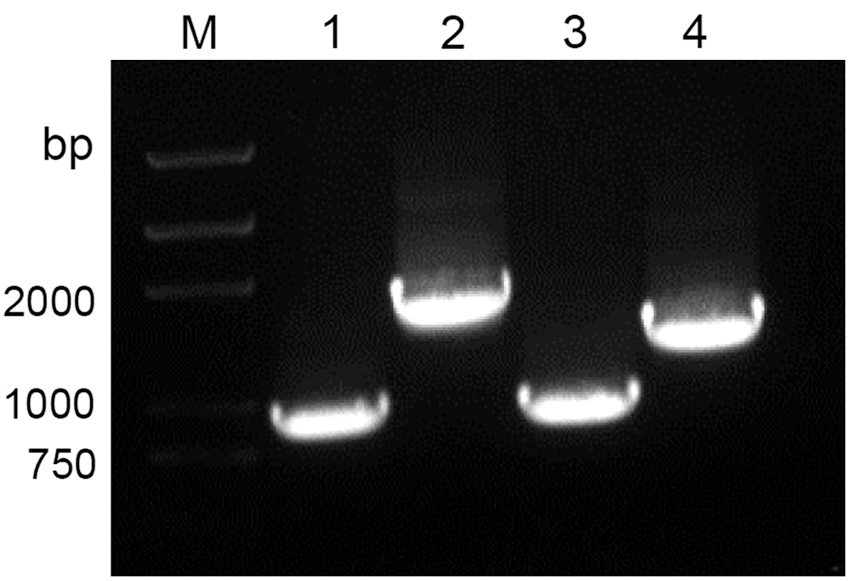
Polymerase chain reaction amplification of VP1, 2ABC, 3ABC, and 3D genes of EV-A71 using the primer pairs contained with restriction sites for BamH I and Not I, respectively. Lane M: DNA marker; lane 1, VP1; lane 2, 2ABC; lane 3, 3ABC; lane 4, 3D.

EV-A71-6His fusion proteins expressed in Escherichia coli and separated on a 15% SDS-PAGE gel. Lane M: protein molecular weight marker; lanes 1, 3, 5, 7: preinduced cells; lane 2, VP1; lane 4, 2ABC; lane 6, 3ABC; lane 8, 3D. Small black triangle symbols (▼) denote the target protein bands. SDS-PAGE, sodium dodecyl sulfate–polyacrylamide gel electrophoresis.
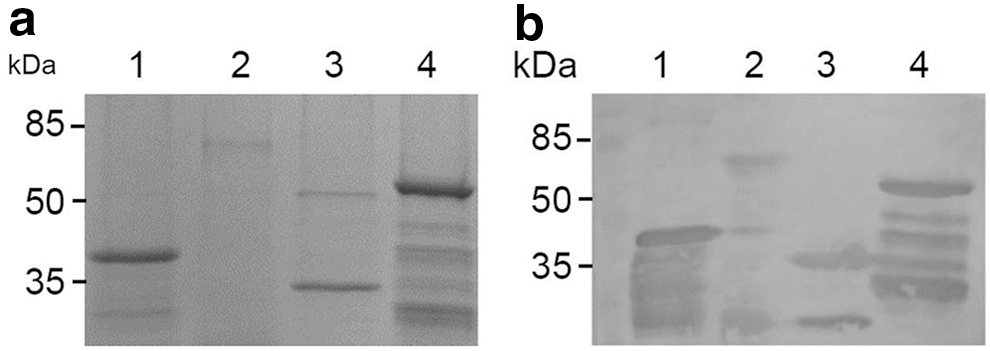
Immunoreactivity of the purified fusion proteins with EV-A71-infected patient sera using immunoblot assay. Purified proteins were separated on a 15% SDS-PAGE gel
Immunoreactivity analysis of the nonstructural proteins of EV-A71
The immunoreactivity of three recombinant nonstructural proteins of EV-A71 was tested separately by immunoblot assay and indirect ELISA with IgG antibodies against EV-A71. As a positive control, VP1 was also included in the immunoreactivity analysis. The results of immunoblot assay show that VP1 can be recognized by IgG antibodies from both patients infected with EV-A71 and mice/rabbits immunized by inactivated EV-A71 virus particles. However, all three nonstructural proteins, including 2ABC, 3ABC, and 3D, can only be recognized by anti-EV-A71 human sera (Fig. 3b), but not by animal immune sera (data not shown). The result of indirect ELISA was similar to that of immunoblot assay, except that these nonstructural proteins showed weak at best immunoreactivity with animal immune sera (Table 2). These results indicate that, VP1, a major structural protein of EV-A71, can induce an IgG antibody response after whether viral infection or immunization with inactivated vaccine. Different from VP1, however, nonstructural proteins can only induce an IgG antibody response through viral infection, but hardly induced after inactivated EV-A71 immunization.
OD450 nm values are expressed as the average of three separate experiments. Standard deviation of OD450 nm values is in parenthesis.
ELISA was carried out using pooled healthy or preimmune sera (n = 3) and animal antisera at a dilution of 1:100.
ELISA, enzyme-linked immunosorbent assay.
Discussion
Immune response, especially antibody response, induced by natural infection of productive virus, has been a valuable research area. The identification of immunodominant B or T cell epitopes will be helpful for the development of serological diagnosis reagents and epitope-based vaccine design. It has long been known that antibodies response against proteins comprising the EV-A71 virion particle can be induced whether by viral natural infection or by inactivated vaccine immunization. These antibodies may react with some linear or conformational epitopes on the capsid surface of EV-A71 virion particle. The detection of these antibodies in the sera forms the basis for serological diagnosis of EV-A71 infection. However, these tests cannot distinguish between an infected person and one who was immunized with the inactivated whole-virus vaccine or viral subunit vaccine.
In the late 1980s, it was observed that animals infected with foot-and-mouth disease virus (FMDV) can induce an antibody response against the viral nonstructural proteins (25). Since then, these nonstructural proteins or its truncated fragments have been used to study the serological detection of FMDV infection (3,6,7,11,12,16,18,22). Hepatitis A virus (HAV) and poliovirus (PV) are important members within the family Picornaviridae. Similar to FMDV, persons naturally infected with HAV or PV can develop significant antibody titers against the viral nonstructural proteins (8,14,19). EV-A71 and PV are both members of the genus Enterovirus within the family Picornaviridae. Presumably, individuals infected naturally with EV-A71 should be able to stimulate an antibody response against the viral nonstructural proteins. However, the spectrum of viral antigens of EV-A71, especially antigenicity of nonstructural proteins, has not been determined so far, even though the virus has been studied intensively for the last several years.
In this study, the 2ABC, 3ABC, and 3D genes of EV-A71, together with VP1 gene, were cloned into the pET28a expression vectors, respectively, and its recombinant proteins fused with 6His-tag were expressed in E. coli. Immunoreactivity analysis showed that, like VP1, each of the nonstructural proteins can react highly with human IgG antibodies against EV-A71, but not with mouse and rabbit anti-EV-A71 IgG antibodies. The difference of origin between these specific anti-EV-A71 antibodies is that, human anti-EV-A71 antibodies were induced by EV-A71 natural infection, and animal anti-EV-A71 antibodies were induced by inactivated vaccine immunization. So, these results indicate that natural infection of EV-A71 can develop an antibody response against both structural proteins and nonstructural proteins. As for the latter, the mechanism of immune response might be involved in lysing of host cells, releasing viral nonstructural proteins for antigen processing. In contrast, vaccinated animals develop antibodies only to the viral structural proteins, which are the major components of inactivated material. In other words, without viral replication, there would be no antibody response against nonstructural proteins.
In conclusion, the results of this study suggest that the antibody response against nonstructural proteins can be developed significantly with natural infection of EV-A71, which is only associated with viral replication. These findings will contribute to a further understanding of the spectrum of viral antigens that induce a humoral immune response during EV-A71 infection, and the development of new serological diagnostic reagents for EV-A71 infection.
Footnotes
Acknowledgments
The authors are grateful to Dr. Jihong Meng (Southeast University, People's Republic of China) for his valuable suggestions and help to Dr. Bo Zhang (Chinese Academy of Science, People's Republic of China) for the gift of infectious clones of EV-A71. This work was supported by the grants from the Natural Science Foundation of Zhejiang Province (grant no. LY16H190005) and the Undergraduate Scientific and Technological Innovation Project of Zhejiang Province (grant no. 2014R426033).
Author Disclosure Statement
No competing financial interests exist.