
Abstract
African swine fever (ASF) is a fatal disease for domestic pigs, leading to serious economic losses in countries where ASF is endemic. Despite extensive research, efficient vaccines against ASF are lacking. Since peripheral blood cells are important mediators for vaccines, we study the impact of ASF on blood parameters in pigs with different ages and infected with different doses of ASF virus. Four different groups were studied: (1) 12 weeks of age/low virus dose; (2) 12 weeks of age/high virus dose; (3) 18 weeks of age/low virus dose; and (4) 18 weeks of age/high virus dose. By varying in age and/or ASFV inoculation dose, we monitor blood parameters during different degrees of disease. Thirty percent of the pigs survived the infection with a moderately virulent strain of African swine fever virus (ASFV). Animals that did survive infection were generally older, independent from the inoculation dose used. A firm reduction in many different cell types at 3–5 days postinfection (DPI) was accompanied by an increase in body temperature, followed by clinical signs and mortality from day 6 PI. While blood parameters generally normalized in survivors, γδ T cells and IL-10 levels could be related to mortality. These conclusions should be considered in new approaches for protection against ASF.
Introduction
A
The disease is often fatal for domestic pigs, characterized by lymphoid depletion and hemorrhages in lymphoid organs (11). ASF outbreaks are generally associated with large economical losses (21). Over the past years, research has increased the knowledge about ASF tremendously, but effective vaccines are still lacking. All proposed vaccine strategies are based on stimulation of (systemic) immune cells, which are, however, most affected by the disease itself (3). Knowledge of the effect of ASF on immunological defense mechanisms is therefore essential and many studies were performed trying to establish the effect of ASF on immunological parameters (29).
The effect of ASF is generally studied in younger pigs (8–12 weeks), often with one inoculation dose of the virus only. While age is of proven importance for Porcine Reproductive and Respiratory Syndrome and Classical Swine Fever (14), there are indications that the process of ASF disease could be age dependant as well (2). Also, the inoculation dose of the virus seems important (12). In this study, we investigate in a comprehensive manner, a number of different hematological and immunological blood parameters that might be relevant for protection against ASF in an age-dependent and inoculation dose-dependent model for the development of ASF.
Under our experimental conditions, older animals better survive an ASF virus infection, independent of the inoculation dose used. During 3–5 days postinfection (DPI), a firm reduction in many different cell types in blood was accompanied by changes in serum cytokine levels and an increase in body temperature, followed from day 6 PI by clinical signs and mortality. Among those changes, we have observed that γδ T cells and serum IL-10 levels could be related to survival against ASF, paving the way for further vaccination strategies against ASF.
Materials and Methods
Animals and housing
Ten pigs of 12 weeks old and 10 pigs of 18 weeks old (±25 and 60 kg, respectively) were used. The pigs were inoculated with a low (3.5 log10 TCID50/mL) or high (5.5 log10/TCID50/mL) virus dose resulting in four experimental groups of five pigs each: (1) 12 LVD (12 weeks of age, low virus dose); (2) 12 HVD (12 weeks of age, high virus dose); (3) 18 LVD, and (4) 18 HVD. The groups were housed in an isolation unit, each pen separated by solid walls with at least an 80-cm space between pens to prevent transmission between the groups. The experiment was approved by the Ethics Committee for Animal Experiments of the Animal Sciences Group of Wageningen UR.
Virus and inoculation
The pigs were inoculated intranasally with 2 mL of the moderately virulent ASFV isolate Netherlands’86 (31). The stock virus was passaged once in primary lung macrophages and the supernatant was then stored as 105.5 50% tissue culture infectious dose (TCID50) aliquots at −70°C. Virus dose of 3.5 log10 TCID50/mL was chosen as the dose that infected ∼50% of the animals (5). Virus dose of 5.5 log10 TCID50/mL was chosen as the maximal yield.
Sampling and observations
The pigs were monitored daily for body temperature, development of clinical signs (5), and mortality. EDTA blood samples were taken at 3, 5, 7, 10, 13, 17, 20, 24, and 27 DPI. Serum samples were taken at 7, 10, 17, and 27 DPI. Samples taken before the experiment (0 DPI) served as controls.
Part of the EDTA blood samples was stored (−80°C) for further analysis. These stored blood samples were used for measuring viral titers by PCR (5). The fresh EDTA blood samples were analyzed within 2 h for total white blood cells (WBCs), lymphocytes, granulocytes, mid-range cells (predominately monocytes), platelets (PLT), and red blood cells (RBCs) using the Medonic® CA 620 (Medicon, Newry, Ireland). After analysis, WBCs were isolated from these samples using Leucosep (Greiner Bio-one, Kremsmunster, Austria) and counted (Countess; Thermo Fisher, Waltham, MA). Fresh isolated cells were used for FACS analysis and the remaining cells were stored in liquid nitrogen for LPT- and NK-assays. Serum samples were stored at −20°C until tested.
To further investigate features of the different blood parameters during the development of the disease, for the analysis, pigs were reassigned to new groups on the basis of mortality: during the experimental period nine pigs died before day 13 PI. Another five animals died between day 13 and 15 PI, while the rest of the pigs (6) survived the treatment. As a consequence of these observations, three groups were made: (1) animals that died before day 13 PI, (2) animals that died on/after day 13 PI, and (3) animals that survived the treatment (Table 1).
DPI, days postinfection; HVD, high viral dose; LVD, low viral dose.
Flow cytometry
Cells (0.5 × 106) were incubated on ice with primary monoclonal antibodies (mAbs; Table 2) for 30 min and washed thrice with the FACS buffer (PBS +1%BSA +0.1%NaAz). For nonconjugated primary mAbs, cells were additionally incubated with labeled secondary mAbs. Subsequently, cells were suspended in 4% paraformaldehyde. Suspensions were measured using a FACSCalibur flow cytometer (BD). The numbers of each cell population per μL blood were calculated as described by Weesendorp et al. (33).
mAbs, monoclonal antibodies.
The combination of anti-CD3, CD4, CD8, CD21, and γδ resulted roughly in the identification of Cytotoxic T Lymphocytes (CD3+CD4−CD8β+), T helper cells (CD3+CD4+CD8α +), memory T cells (CD3+CD4+CD8α −), NK cells (CD3−CD4−CD8α +), γδ T cells (CD3+ γδ +), and B cells (CD3−CD21+) (10, 34). The combination of anti-CD4, CD8, and CD25 resulted in the identification of natural (CD4+CD25+) and activated/memory-like regulatory T cells (Tregs) (CD8α +CD25+) (13). The combination of anti-CD3 and CD107 resulted in the identification of activated CTL (CD3+CD107+) and NK cells (CD3−CD107+) (32). The combination of anti-CD14 with CD163 resulted in the identification of CD14hiCD163− and CD14loCD163+ monocytes (19).
Lymphocyte proliferation test
Thawed PBMC (2.5 × 105 per well) were incubated for 72 h at 37°C with 5 μg/mL ConA in a culture medium (CM; RPMI supplemented with FCS and pen/strep). For the last 18 h of incubation, 1% Alamar Blue (Thermo Fisher) was added. Cells were pelleted (3 min, 175 g) and 100 μL supernatant from each well was harvested and measured for absorbance (570 and 600 nm) on a SPECTROstar (BMG Tabtech, Offenburg, Germany). OD values are converted to cell counts by a cell standard curve and the stimulation was expressed as the fold change compared to the medium control. Since the different cell types may be not equally hampered by the treatment and differences in ASF sensitivity between groups exist, the proportion of ConA reactive cells may vary between groups and sampling points, therefore possibly troubling the functional measurements.
NK assay
Fresh K562 cells (106/mL) were incubated with 0.25 μM Carboxy Fluorescein Succinimidyl Ester (CFSE) in PBS at 37°C for 10 min, then washed with PBS/FCS and suspended in CM (5 × 105/mL). Thawed PBMC (107/mL CM) supplemented with 104 IU/mL recombinant porcine IL-2 and labeled K562 cells were mixed with effector cells in a 60:1 effector–target ratio (E:T) and incubated overnight at 37°C. Suspensions were washed in PBS, and propidium iodide (PI; Sigma, St. Louis, MO) was added at a final concentration of 5 μg/mL. K562 killing (PI+) was measured on a GUAVA easyCyte (Meck Millipore, Darmstadt, Germany). Controls consisted of K562 cells, incubated with IL-2 only. Specific lysis was calculated as ([sample PI+−control PI+]/[100%−control PI+]).
Cytokine Elisa
Porcine IL-1, IL-6, IL-8, TNFα, TGFβ (R&D, Minneapolis, MN), IL-4, IL-10 (Invitrogen, Camarillo), IL-12, IL-17 (Thermo scientific, Waltham, MA), IFNα, IFNγ (Genorise, Glen Mills, PA), and IL-18 (Affimetrix, Santa Clara, CA) Elisa kits were used to determine serum cytokine levels. Based on these cytokines, a rough indication of proinflammatory Th1, Th2, Th3, and Th17 responses can be evaluated (1,9). ELISA was performed according to manufacturer's instructions and the levels of cytokines were measured using a colorimetric reader SPECTROstar (BMG Labtech) based on the cytokine standard curve supplied with the kit.
Statistical analysis
Data are expressed as the arithmetic mean ± SD. Generally, differences between groups were analyzed by one-way ANOVA. The Fisher's exact test was used to measure differences in mortality between groups. A p-value <0.05 was considered statistically significant.
After 10 DPI, the number of pigs became too small for reliable statistical comparison between groups. Therefore, only data till day 10 PI were used for statistical analysis. When groups were compared on the basis of mortality, statistical comparison could be performed until 13 DPI.
Results
Observations
A rapid increase in blood virus titers in all inoculated pigs was observed, reaching a maximum at 5 DPI (Fig. 1A), while fever developed in all animals (Fig. 1B) followed by the development of clinical signs (Fig. 1C) and mortality (Fig. 1D). Although not significant, at 3 DPI, two interesting trends were seen in blood virus titers (Fig. 1A): first, the high inoculation dose caused higher blood virus titers in both younger and older pigs compared to the lower inoculation dose. Second, when comparing equal inoculation doses, blood virus titers were higher in the younger animals. After 5 DPI, the blood virus titers seemed to reach a maximum without any differences between the groups.
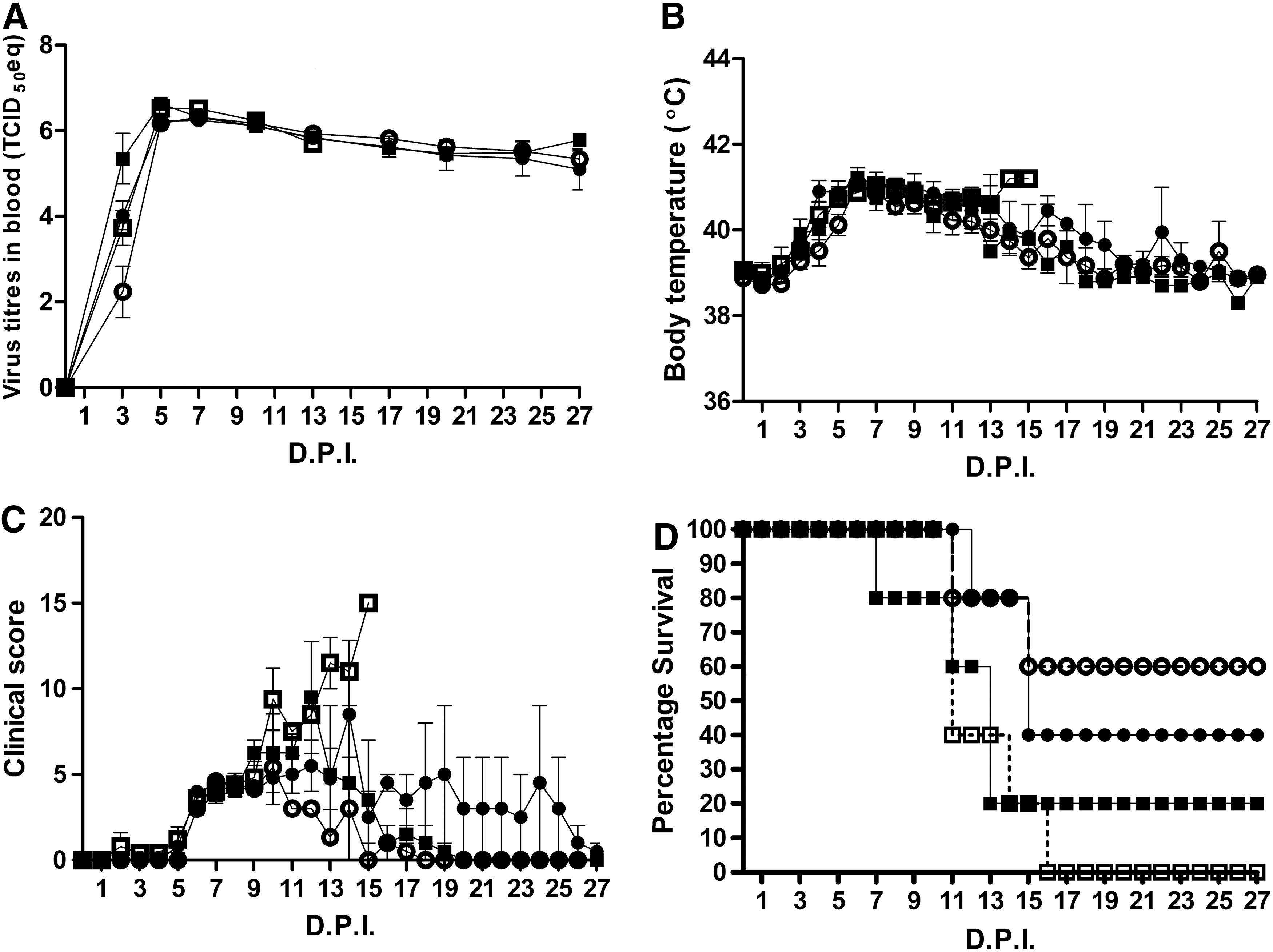
Observations after ASFV inoculation:
Body temperature reached a maximum between 5 and 7 DPI, after which it slowly normalized (Fig. 1B). The clinical score (Fig. 1C) seemed related to cases of mortality (Fig. 1D). After 15 DPI, the remaining pigs survive and the clinical signs disappear. Group 12 LVD seemed to diverge at 10 DPI from the other groups in body temperature and clinical score at the moment that the last animals of this group are dying.
ASF was lethal for 70% of the pigs from our experiment within 15 DPI. Although not statistically significant, younger pigs seemed to be more sensitive to the infection, compared to the older pigs. Notably, more of the younger pigs died because of the infection (12 weeks: 90% mortality vs. 18 weeks: 50% mortality). Moreover, at 10 DPI, when the first animal died from the group of older pigs, already 50% of the younger pigs had died.
Blood cell counts
Similar for all groups, a strong reduction in WBC count was detected between 3 and 5 DPI (Fig. 2A), which was mainly caused by a drop in the number of lymphocytes (Fig. 2B), whereas granulocytes were only slightly affected (Fig. 2C). Mid-range cell counts also dropped by day 5 (Fig. 2D), but these cells only constitute a relatively small percentage of the total WBC.
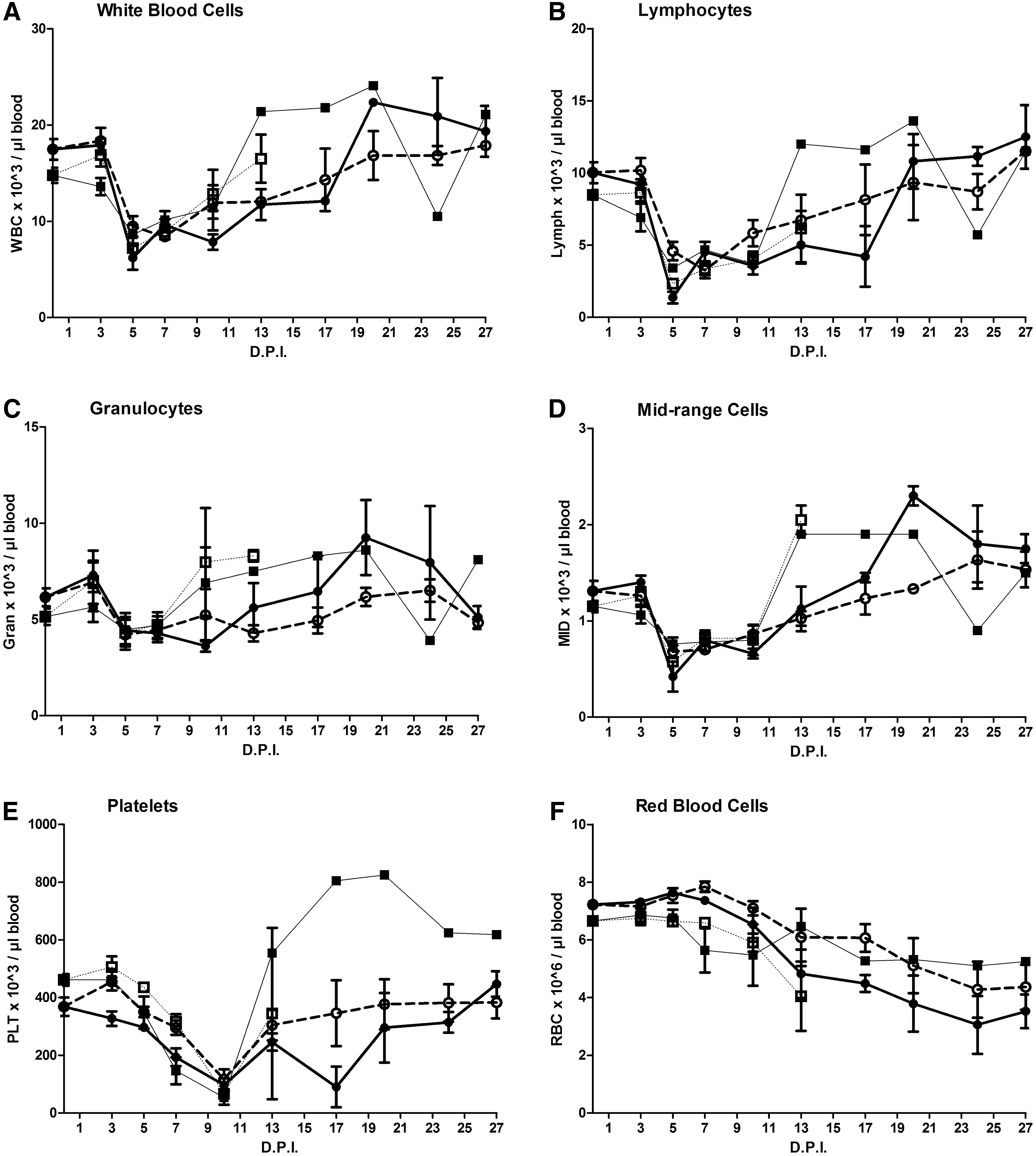
Frequency of
A strong reduction in platelet counts was also observed (Fig. 2E). However, in comparison to the WBC, the decrease in platelet count was delayed and reached its minimum around day 10 PI. Only a slight gradual decrease toward the end of the experimental period was observed for RBC counts (Fig. 2F).
When the pigs were reassigned to new groups on the basis of mortality, significant differences were found for RBC and PLT only (Fig. 3). Compared to the surviving animals, a significant decrease in RBC counts was seen on day 10 and 13 PI in animals that died between day 10–13 and 13–15 PI, respectively. A decrease in PLT count was seen on day 10 PI in animals that died between day 10 and 13 PI.
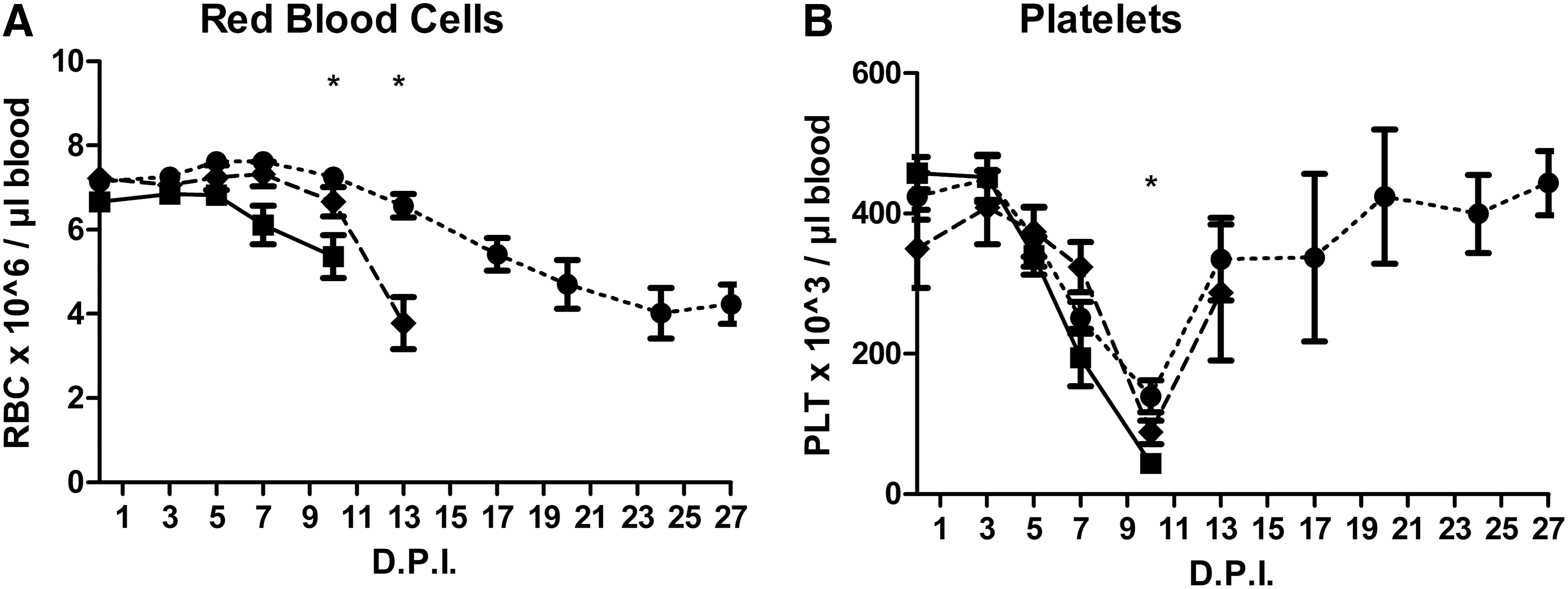
Frequency of
Flow cytometry
The patterns of the immune cells were largely comparable for the different groups (Fig. 4). No differences were found at 0 DPI (only differences in age). Besides some exceptions (Fig. 4A–C, E, H, J), especially on day 5 PI, no significant differences between the groups were observed. Roughly, three patterns were recognized.
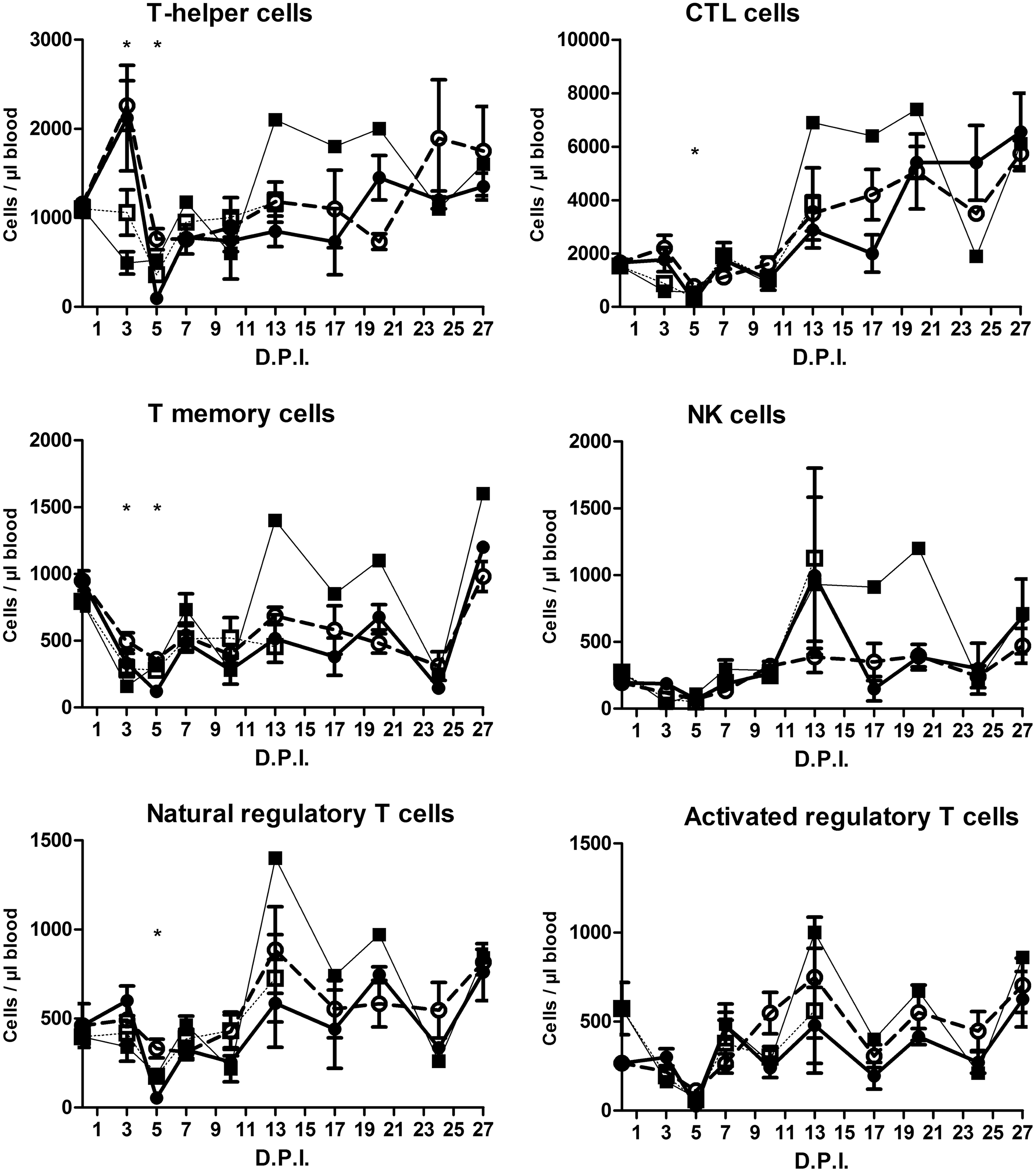
Frequency of peripheral immune cells (see header of each individual graph). Pigs of 12 or 18 weeks of age were inoculated with a low (LVD; 3.5 log10/mL TCID50) or high (HVD; 5.5 log10/mL TCID50) virus dose of the moderately virulent ASFV isolate Netherlands’86 resulting in four experimental groups: (□) 12 LVD; (■) 12 HVD; (○) 18 LVD; and (●) 18 HVD. Pigs were sampled on the indicated days. Asterisks in the figure indicate significant differences between groups (p < 0.05).

First, a pattern was seen comparable to that of WBC counts: a firm drop in counts around 5 DPI after which, generally, a recovery was seen. T helper cells (Fig. 4A), CTL cells (Fig. 4B), natural Tregs (Fig. 4E), activated Tregs (Fig. 4F), B cells (Fig. 4G), γδ T cells (Fig. 4H), activated NK cells (Fig. 4J), and CD14hiCD163− (Fig. 4K) cells belong to this group. The T helper-CTL and γδ T cells differ somewhat from the WBC pattern in the way that reduced counts were already seen for the younger animals at 3 DPI.
The second group consisted of memory T- (Fig. 4C), NK- (Fig. 4D), and activated CTL cells (Fig. 4I), where the maximal decrease in counts was already reached after 3 DPI.
The third group consisted of CD14loCD163+ (Fig. 4L). The pattern of these cells differs remarkably from the cells of the first two groups. Only minor changes were seen during the first 5 days of infection after which a firm increase was seen at 10 DPI.
When the groups were redefined on the basis of mortality, significant differences were found for CTL and γδ T cells (Fig. 5).
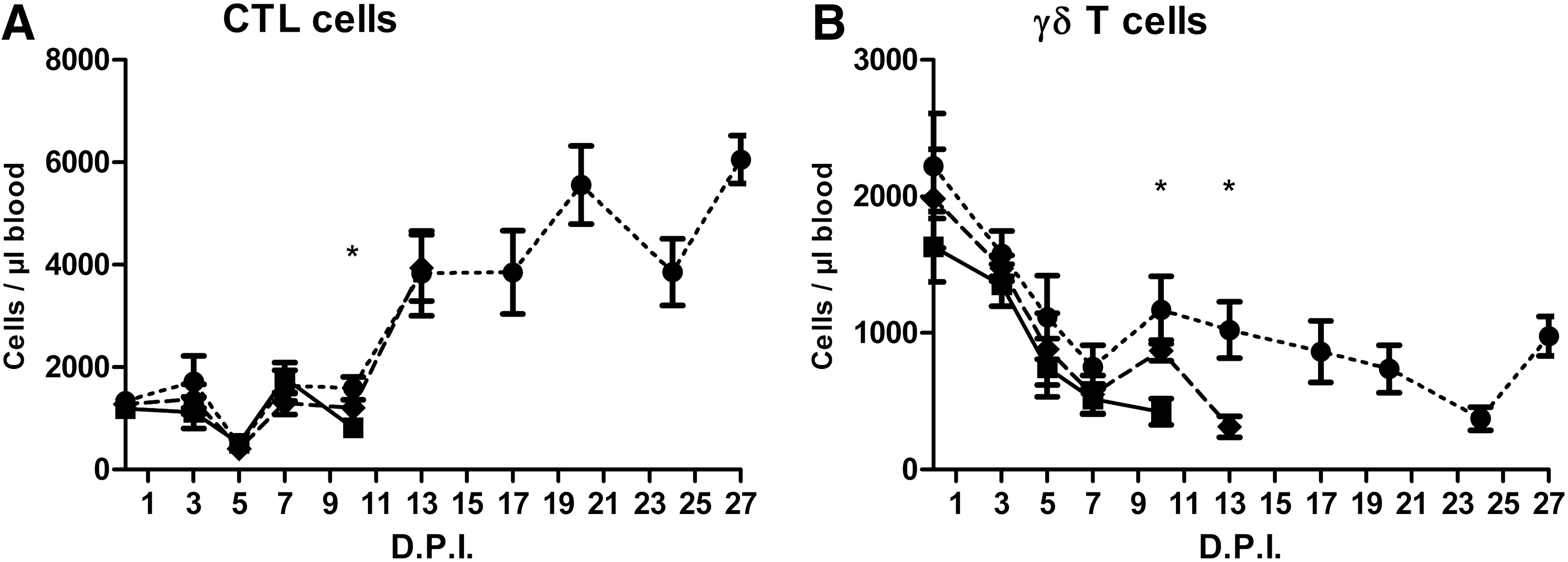
Frequency of
Lymphocyte proliferation assay
The response on the mitogen ConA decreases slowly until 5 DPI, with a significant difference for only group 2 between day 0 and 5 PI (Fig. 6A), after which the responsiveness was restored. No significant differences were found when the groups were compared mutually.

Functional assays. Liquid nitrogen-stored cells were tested for the response against
NK assay
The specific lysis tended to decrease toward 5 DPI, but no significant differences were found between groups or DPI (Fig. 6B).
Serum cytokines
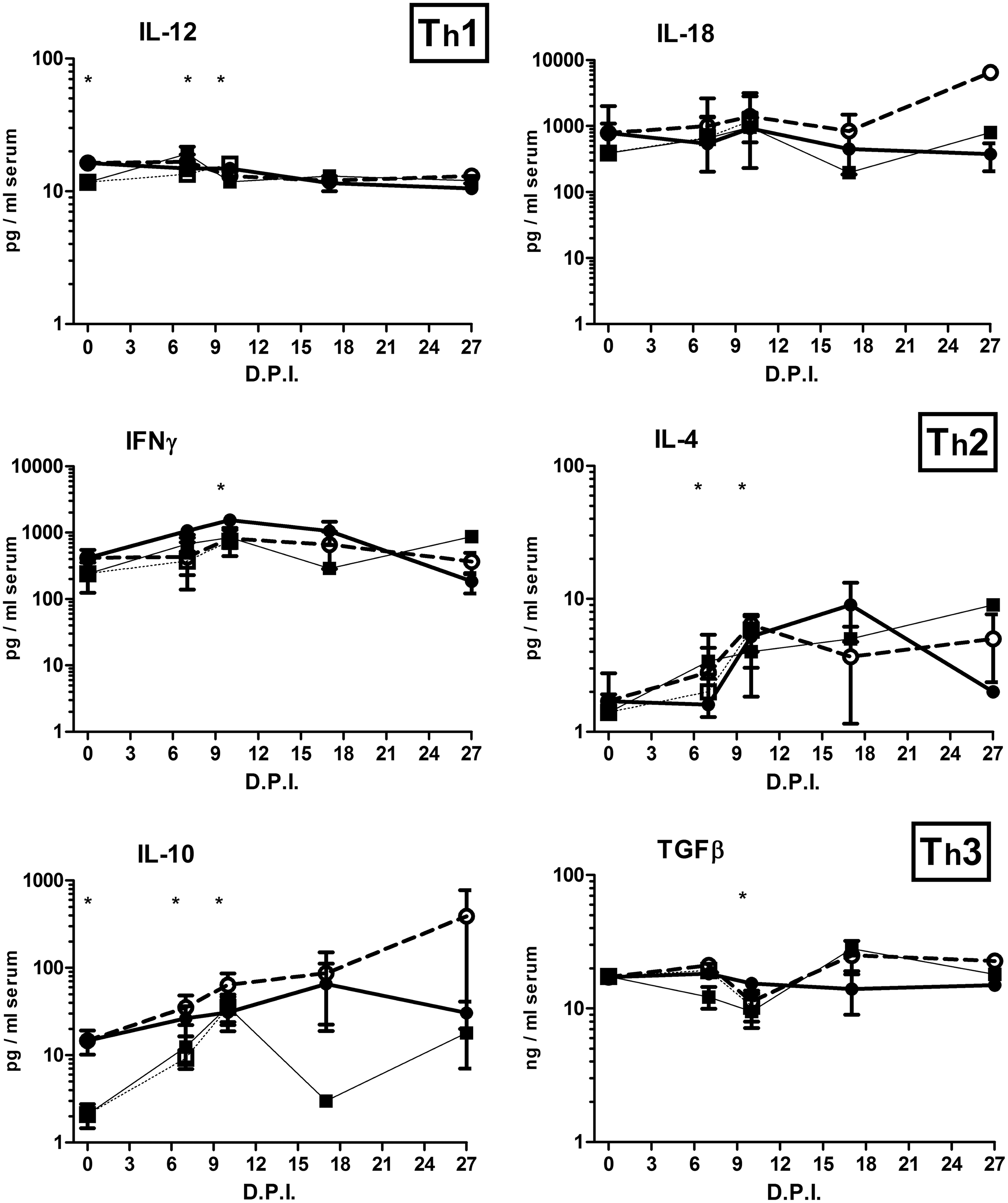
Apart from a few exceptions, cytokine levels remained stable over the experimental period (Fig. 7). An age effect (at 0 DPI) was found for TNFα, IL-12, and IL-10: older pigs started the experiment with significantly higher TNFα, IL-12, and IL-10, the latter one being the most dramatic. Significant increases compared to 0 DPI were especially found for younger pigs at 7 or 10 DPI for TNFα, IL-4, IL-10, and IL-12. Nonstructural significant differences between groups were found for IL-6, IL-8, IL-17, IFN-γ, and TGFβ, possibly indicating differences in the development of responses. Only the serum concentration of the anti-inflammatory cytokine TGFβ was significantly decreased at 10 DPI. The peak values of IL-6 and IL-8 on day 10 PI resulted from one (the same) divergent pig only. When the groups were redefined on the basis of mortality, differences were only found for IL-10 (Fig. 8): survivors started at 0 DPI with a significantly higher IL-10 level.

Serum cytokines (see header of each individual graph). Pigs of 12 or 18 weeks of age were inoculated with a low (LVD; 3.5 log10/mL TCID50) or high (HVD; 5.5 log10/mL TCID50) virus dose of the moderately virulent ASFV isolate Netherlands’86 resulting in four experimental groups: (□) 12 LVD; (■) 12 HVD; (○) 18 LVD; (●) and 18 HVD. Pigs were sampled on the indicated days. Asterisks in the figure indicate significant differences compared to 0 DPI (p < 0.05). Asterisks at 0 DPI indicate significant differences due to age (12 weeks vs. 18 weeks; p < 0.05).
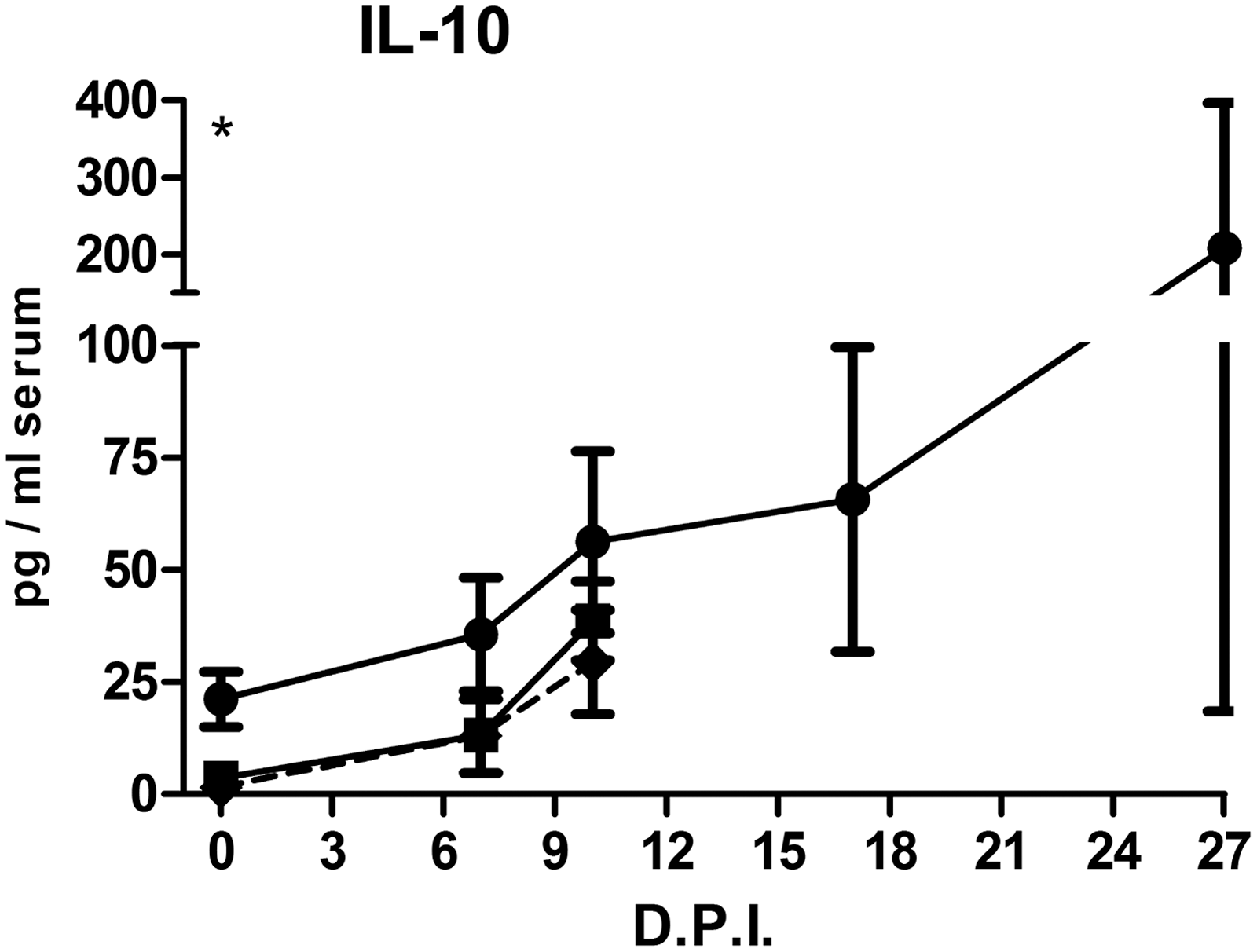
Serum IL-10 in relationship to the time of death. Regardless of age or virus dose, pigs were classified into three groups by the moment of death: (■) animals that died before 13 DPI, (♦) animals that died on/after 13 DPI, and (●) animals that survived ASF infection. Pigs were sampled on the indicated days. Asterisks at 0 DPI indicate significant differences due to age (12 weeks vs. 18 weeks; p < 0.05). ASF, African swine fever.
Discussion
The objective of this study was to increase knowledge of the impact of ASF on different blood parameters that might be involved in the immune response against ASFV in animals of two different ages, infected with two different doses of virus. In summary, age seems of more importance than virus dose for the development of ASF disease, possibly through blood γδ T cells and serum IL-10 concentrations.
There is a tendency for a relationship between age and mortality as five out of six surviving animals were of older age. An additional check was carried out for a possible correlation between fatality rate and genetic background of the pigs (litter of origin), but none was found (data not shown). Differences in blood viral titers were found only at 3 DPI. Differences in inoculation dose might explain the dose-related difference in blood virus titers, while age-related differences in virus titer might be provoked by a simple dilution or hematological differences. Our results differ from the findings of de Carvalho Ferreira et al.,5 who inoculated 12-week-old pigs with 1 mL of the low dose (3.5 log10 TCID50/mL) of the same ASFV strain and found no mortality. It remains ambiguous whether the slight difference in dose volume counts for the considerable difference in disease development between our studies.
Two immune cell subtypes that correlated to the time of death were CTL and γδ T cells. Particularly, for γδ T cells, there was a strong correlation with survival as we found on day 10 and 13 PI, significantly lower γδ T cell counts in pigs that did not survive the next few days. However, it remains to be explored whether γδ T cells have a direct function in protection against ASF or if they are merely indicative. In contrast to man and mice, porcine peripheral blood contains a large proportion of γδ T cells, which probably contribute to the “innate” phase of immune responses to viruses (28,34). Porcine γδ T cells are related to protection against Foot and Mouth Disease after vaccination by the production of a variety of cytokines and chemokines and antigen presentation (28). Hampered antigen presentation was also observed for ASF and it was suggested that γδ T cells may overcome the compromised antigen presentation by cells of the mononuclear phagocytic system (29). A plausible hypothesis could be that a reduced amount of γδ T cells may adversely affect the immune response to ASFV. The important role of γδ T cells in the course of ASF was further supported when groups were compared on the basis of age only (12 weeks vs. 18 weeks), independent of inoculation dose. Just before the critical phase of the disease (7 DPI) when clinical signs became obvious and the first (younger) pig died, a significant higher amount of γδ T cells was found in the blood of the older pigs (data not shown). The effect of γδ T cells may be mediated by IL-6 and IL-8, which were, in our study, significantly increased at 7 DPI in the older (more resistant) pigs (Fig. 7B, C) and are known mediators for γδ T cell activation (28). No significant differences on the basis of age were found on other DPI.
The relationship between reduced amounts of CTL cells and time of death was less clear. All animals seemed to recover after the drop 5 DPI, after which some animals again showed reduced CTL counts and died. However, at 13 DPI, no reduction of CTL counts was found in animals that died shortly afterward. Also, no significant differences on the basis of age (independent of inoculation dose) were found for CTL cells. Finally, activated CTLs (CD107+) recovered quickly from the initial decrease and seemed unaffected at the critical moment of disease. CTL cells contribute in a challenge model to the clearing of ASFV in outbred pigs (23). However, the same study also demonstrated the ambivalent importance of CD8β CTLs for inbred pigs, confirming the controversial relationship between systemic CTL cells and ASF survival.
Before inoculation (0 DPI), no significant age-related differences (12 weeks vs. 18 weeks) were found for WBC counts. Small, but significant differences were, however, found for RBC and platelet counts at this time point. Age-related differences in WBC counts were found in earlier studies, however, only shortly after birth (30). Independent from age or inoculation dose, a clear effect of the ASFV infection on total WBC, lymphocyte, MID cell, and platelet counts was found. As lymphocytes are the predominant cell type in blood, the drop in WBC counts was primarily provoked by lymphopenia (3). Since apoptosis of systemic monocytes is not convincingly established (3) the decrease in MID cells may also be the result of migration of monocytes and macrophages (m-Mφ) toward lymphoid tissues during ASFV infection (24). The decrease in cell counts became visible simultaneously with a raise in body temperature (3 DPI), but well before the occurrence of clinical signs preceding death (from 6 DPI). In our study, the effect of ASF on granulocyte and RBC counts was limited.
For the younger pigs, our cell counts fit well between the range of earlier findings (18,26). To our knowledge, no recent data are available for 18-week-old pigs.
A firm drop in the majority of measured cells 3–5 DPI coincides with the drop in WBC, but, comparable with Leitão et al.,16 without clear differences of responsiveness to the T cell mitogen ConA. As noticed for WBC, the decrease does not coincide with clinic signs and mortality caused by ASF, as the counts of most cell types have already normalized at the moment that first animals died. On days 3 and 5 PI, some significant differences were found between groups, but without a clear direction. However, when cell counts in blood were compared on the basis of age (12 weeks vs. 18 weeks; independent from dose), a significant higher amount of different types of αβ T cells and NK cells were found for the older pigs at 3 DPI (data not shown). These results are not in line with the findings of Lacasta et al., who found no differences in CD4+ (and SWC+, CD8+, CD21+) cells between 7-week-old pigs that did survive or did not survive a challenge (15). No differences were found when cell amounts in blood were compared on the basis of inoculation dose (independent of age).
Activated NK cell counts (CD3-CD107+) did show significant differences between groups during the period of clinical signs and mortality caused by ASF (7 and 10 DPI). NK cells are certainly of importance in defending against ASF (16), but neither did we find a relationship between activated NK cells and clinical signs or time point of mortality (data not shown) nor did we find differences of functionality in the NK assay for this virus strain (Fig. 6).
We found a decrease in the percentage of B- and Tregs-positive cells in blood within 5 DPI. A decreased CD21 and CD25 expression may result from general T and B cell apoptosis (22) in combination with the fact that products from ASFV inhibit the expression of CD21 (27) and activation markers like CD25 (4).
In accordance with the drop in monocytes, we found reduced numbers of CD14hiCD163− cells in blood 5 DPI, while the counts of CD14loCD163+ cells hardly changed except for a peak on day 10 PI. Although the nomenclature of these cell populations varies, it is well understood that increased CD163 expression is related to the maturation of monocytes (6) and to the susceptibility for ASFV infection (17). The low baseline values of the more mature “inflammatory” CD14loCD163+ cells and peak counts 10 DPI may therefore reveal maturation of monocytes under influence of ASFV, as was seen for Salmonella and Actinobacillus pleuropneumoniae infections (19,20). The reason for the rapid disappearance of CD14loCD163+ cells from blood afterward was not clear, but may be related to migration to infected organs (19). Despite the close relationship of these cell types with ASFV entry and immunological defense in general, we could not relate counts of these cell types to the course of disease.
The levels of most cytokines were comparable between the groups and were slightly elevated during the initial experimental period, after which the levels generally normalized. Only TNFα, IL-4, IL-10, and IL-12 deviate from this pattern: (1) significant age-related differences were found before the infection (TNFα, IL-10, and IL-12), (2) the initial significant lower levels of younger pigs increased significantly toward levels similar to older pigs (TNFα, IL-10, and IL-12), and (3) IL-4/IL-10 levels remain high compared to the initial levels. When pigs were grouped by age only (12 weeks vs. 18 weeks; independent of inoculation dose), significantly higher IL-10 levels were found in the serum of older pigs, not only on 0 DPI (Fig. 7) but also on 7 DPI (data not shown). The important role of IL-10 in ASF disease was further supported by the finding that survivors had initially significantly higher serum IL-10 levels. Differences in IL-10-related disease development might also be the consequence of polymorphisms, as for the porcine IL-10 gene, 21 polymorphisms were detected (8).
IL-10 and IL-4 belong to the Th2 cytokines, which are powerful inhibitors of proinflammatory responses, thereby influencing the proposed inflammation-related development of lesions in organs (11).
IL-12 is a known mediator of Th1 responses, but we found no relationship with the Th1 cytokines (IL-18 and IFN-γ) and speculate that development of a firm Th1 response is blocked by the increase of the Th2 cytokines (Il-4/IL-10), which increased simultaneously with IL-12 in younger pigs.
Although γδ T cells are potent producers of IL-4, IL-10, and IL-12 (among others), no correlation between these cytokines and γδ T cells was found (data not shown).
The inoculation dose of strain Netherlands’86 appears of little importance for the course of the disease. Independent of inoculation dose, age does seem to have an influence on the development of the disease. Compared to younger pigs, the initial higher serum levels of TNFα, IL-10, and IL-12 cytokines in older pigs were followed by lower blood virus titers and higher amounts of blood αβ T cells at 3 DPI and prolonged higher serum levels of IL-10 and higher amounts of γδ T and activated NK cells at 7 and/or 10 DPI. Only IL-10 and γδ T cells could be related to mortality. How all these features are connected is not clear and needs further exploration. IL-10, which seems to play a crucial role in our study, is considered an inhibitory cytokine. However, prolonged higher IL-10 serum levels and the finding of higher amounts of αβ T cells at 3 DPI and higher amounts of γδ T and activated NK cells at 7 and/or 10 DPI in surviving pigs may demonstrate the pleiotropic effects of IL-10 (7).
Taken all the findings together, a possible explanation could be that cytokines like IL-10 may prevent uncontrolled (proinflammatory) cytokine production and tissue damage allowing αβ T cells to facilitate an effective immune response in consultation with γδ T cells.
Footnotes
Acknowledgment
This research was funded by the Dutch Ministry of Economic Affairs (WOT-01-003-010-01).
Author Disclosure Statement
No competing financial interests exist.