
Abstract
Programmed death-1 (PD-1) expression has been revealed to be upregulated on T cells and contributes to T cell exhaustion in patients with hepatitis B virus (HBV) infection. In this study, we investigated the dynamic expression of programmed death ligand-1 (PD-L1), the ligand of PD-1, on circulating CD14+ monocytes and CD19+ B cells of HBV-infected patients at the stages of chronic HBV (CHB) infection, liver cirrhosis (LC), and hepatocellular carcinoma (HCC), respectively. The results showed that compared with healthy controls, the levels of PD-L1 expression on CD14+ and CD19+ populations were both upregulated in CHB, LC, and HCC groups. Although there was no significant difference of PD-L1 expression on CD14+ population among three disease groups, further analysis demonstrated that the frequency of CD14+PD-L1+ population was negatively correlated with HBV DNA load, the levels of alanine aminotransaminase (ALT), and the levels of aspartate aminotransferase (AST), respectively, at CHB stage, while it did not present significant correlation with such parameters at LC stage and was only positively correlated with HBV DNA load at HCC stage. Similarly, the levels of PD-L1 expression on CD19+ population also did not present much difference among three disease groups. Intriguingly, the frequencies of CD19+PD-L1+ population at CHB and LCC stages were both positively correlated with the levels of ALT and AST, but they were not significantly correlated with HBV DNA load. Thereby, the current study elucidated the dynamics of PD-L1 expression on monocytes and B cells, along with the dynamic regulation of PD-1 on T cells, which had a close relationship during the progression of HBV infection. Collectively, our findings demonstrated that in the course of HBV infection development, PD-L1 expression on CD14+ monocytes and CD19+ B cells varied and significantly correlated with clinical parameters, which could be utilized as a potential clinical indicator.
Introduction
O
It has been well known that the progression of HBV infection is closely correlated to the function of immune system among which HBV-specific T lymphocytes play an important role for viral clearance (14). To eradicate HBV, it is necessary for T cells to be activated and become effector T cells (23). The two-signal model of T cell recognition assumes that T cell activation is controlled by two kinds of signals from antigen-presenting cells. The first signal is derived from the interaction between T cell receptors and major histocompatibility complex upon antigen presentation. The second signal triggered by cosignal molecules can either be positive or negative to modulate immunocyte proliferation, activation, and differentiation. HBV-specific T cells are the main effector cells for antiviral response (20).
However, their function is usually suppressed and blocked in CHB infection, which is generally characterized by impaired cytotoxic T lymphocyte activity, insufficient antiviral cytokines, and increased inhibitory molecules or regulatory T cells (Tregs) suppressing the proliferation of effector T cells (22). Hence, HBV cannot be effectively eliminated with an increased risk of CHB, LC, and HCC. One of the remarkable findings that may explain the mechanism underlying T cell malfunction in CHB infection is coinhibitory receptors expressed on HBV-specific T lymphocytes with similarities to that in tumorigenesis (5,9). The group of coinhibitory receptors is now defined as the immune checkpoint molecules, including cytotoxic T-lymphocyte associated antigen-4, programmed death-1 (PD-1), B and T lymphocyte attenuator, Lymphocyte-activation gene-3, T-cell immunoglobulin mucin-3, and so on, which play a negative role in inhibition of activation and proliferation of T cells and antiviral cytokine production (27).
Our previous study proved that the levels of PD-1 expression on peripheral CD4+ and CD8+ T cells in CHB were positively correlated with viral DNA load and liver function injury (31). In addition, we found that the expression of programmed death ligand-1 (PD-L1), the ligand of PD-1, on circulating Tregs of CHB patients was obviously elevated and changed dynamically, associated with liver damage and Treg expansion (12), indicating that by engaging with PD-L1 expressed on Tregs or other subsets, PD-1 on T cells would be involved in T cell dysfunction in CHB. Moreover, it was reported that blocking PD-1/PD-L1 engagement in the total CD8+ T cells during the immune clearance stage of CHB infection significantly enhanced the expansion of T cells in the immune clearance stage of CHB infection (13).
Based on the findings of the characteristics of PD-L1 expression on Tregs of CHB patients and its clinical significance, in this study, we aim to further explore the patterns of PD-L1 expression on CD14+ monocytes and CD19+ B cells, the two important subsets in the peripheral blood, since their correlations with the progression of HBV infection have not been well described. For this reason, we determined the PD-L1 expression on circulating monocytes and B cells from CHB, LC, and HCC patients and analyzed the correlations of dynamics of PD-L1 expression with clinical parameters, including HBV DNA and liver injury.
Materials and Methods
Subjects
Thirty-four patients with CHB infection, enrolled in this study, were positive for HBsAg with persistent elevated serum alanine aminotransaminase (ALT) level for 6 months. The patients were identified with detectable HBV DNA level. Thirteen LC patients were serum HBsAg positive carrying HBV DNA. The morphology of liver was examined by computed tomography, which showed evidence of cirrhosis. Eleven HCC patients with positive serum HBsAg and elevated levels of alpha-fetoprotein had multiple arterial vascular nodules by computed tomography imaging examination. All the patients were negative for acute hepatitis B, antibodies (Abs) to Hepatitis C virus (HCV), Hepatitis D virus, human immunodeficiency virus, combined tumor, and other symptoms of liver damage such as autoimmune disease, alcoholic liver disease, and drug-induced hepatitis. None of the patients received antiviral therapy and/or immune therapy before they were enrolled (clinical baseline is listed in Table 1). 21 healthy blood donors served as normal controls. Healthy controls (HC) were serum HBV antigen and antibody negative and did not show infection of viral hepatitis and other autoimmune diseases. All the patients and HC were Chinese. Our study was approved by the local ethics committee, and all patients provided their written informed consent.
Data are presented as number or mean ± standard error of the mean.
BCLC, Barcelona Clinic Liver Cancer; CHB, chronic hepatitis B virus; HC, healthy controls; LC, liver cirrhosis; HCC, hepatocellular carcinoma; HBeAg, hepatitis B virus e antigen; HBV-DNA, hepatitis B virus-DNA; ALT, alanine aminotransaminase; AST, aspartate aminotransferase; ALB, albumin; TBIL, total bilirubin; NA, not applicable.
Virology assessment
HBsAg, HBeAg, HBsAb, HBcAb, and HBeAb were detected by enzyme-linked immunosorbent assay with commercially available kits (Abbott Ireland, Sligo, Ireland). Serum HBV-DNA level was measured by fluorescent quantitative PCR with commercially available kits (Shanghai Shenyou Technology Co. Ltd., Shanghai, China). Positive samples had a viral load >5 × 102 copies/mL. Biochemical analysis was applied to measure ALT, aspartate aminotransferase (AST), and other liver function.
Flow cytometry
Heparin-anticoagulated blood was collected from healthy donors and HBV patients and used directly for fluorescence-activated cell sorting (FACS). Cells were stained with fluorescein isothiocyanate (FITC)-, phycoerythrin (PE)-, PE-Cy5, Peridinin chlorophyll (PerCP), and allophycocyanin-labeled monoclonal Abs, respectively, according to their manufacturers' instructions. One hundred microliter sample for each test was incubated with anti-human CD45-PE-Cy5, CD14-FITC, and PD-L1-PE or CD45-PE-Cy5, CD19-PerCP, and PD-L1-PE for 15 min. Lysed blood was prepared by RBC lysis buffer and washed twice in phosphate-buffered saline and analyzed immediately. FACS data were analyzed using FlowJo software (San Carlos, CA). The CD45-PE-Cy5, CD14-FITC, CD19-PerCP, and PE-labeled anti-PD-L1 monoclonal Abs were purchased from BD Pharmingen (BD Biosciences, San Jose, CA).
Statistical analyses
Statistical analyses were performed using GraphPad Prism 6.0 software showing mean ± standard error of the mean. Mann–Whitney U tests of SSPS 12.0 were used to assess the difference between different groups. The Spearman correlation coefficient was calculated to assess PD-L1 expression and HBV viral load and ALT or AST level. A two-tailed p < 0.05 was considered statistically significant.
Results
Investigation of the frequency of CD14+ monocytes and PD-L1 expression on CD14+ monocytes of CHB, LC, and HCC patients
The baseline characteristics of healthy donors and HBV patients are provided in Table 1, including age, gender, Child-Pugh score, Barcelona Clinic Liver Cancer stage, HBeAg sera-status, serum HBV DNA concentrations, serum ALT and AST levels, albumin, and total bilirubin. The case groups included patients with CHB (n = 34), LC (n = 13), and HCC (n = 11). We initially analyzed CD14+ monocytes among CD45+ population of CHB, LC, and HCC patients (Fig. 1A). There was a significant increase of the frequency of circulating CD14+ monocytes in HCC group than the HC group (p < 0.05) (Fig. 1B), while there was no significant difference between CHB and HC groups, as well as between LC and HC groups (Fig. 1B). Meanwhile, the absolute monocyte count was evaluated. LC and HCC patients had a decreased absolute count of CD14 monocytes in comparison with CHB group (both p < 0.05). Interestingly, the levels of PD-L1 expression were all higher on the surface of monocytes in the CHB, LC, and HCC groups than HC (all p < 0.05), while no significant difference of PD-L1 expression was found among three types of patients (Fig. 1D).
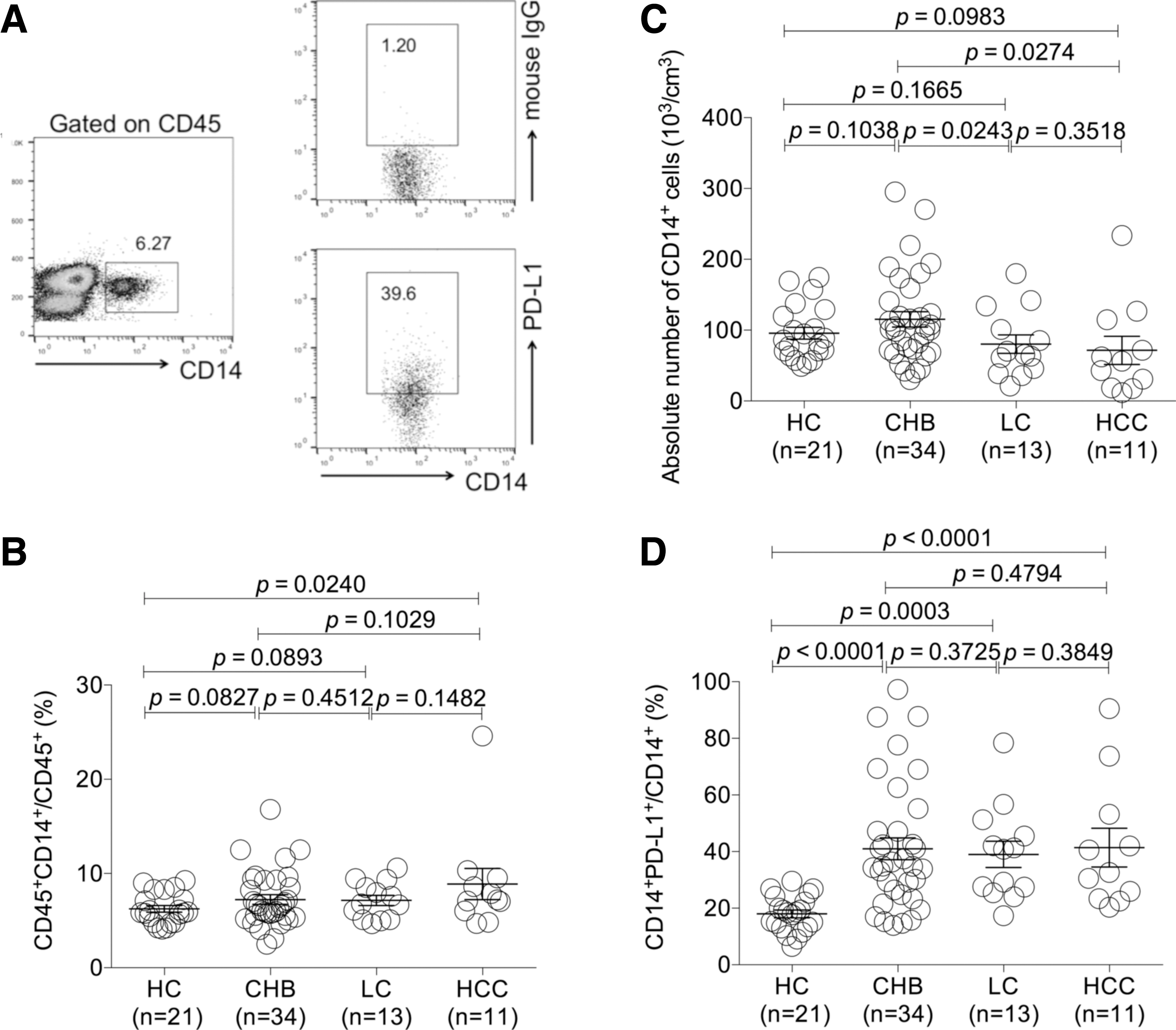
The expression profiles of PD-L1 on peripheral CD14+ monocytes of CHB, LC, and HCC patients.
Analysis of the correlation of the levels of PD-L1 expression on CD14+ monocytes with HBV DNA load and ALT and AST levels
Our previous study described that PD-1 expression on CD4+ and CD8+ T cells changed dynamically in the HBV infection progression with a significant increase in CHB, LC, and HCC groups (31). In addition, we also proved its ligand PD-L1 on CD4+CD25+Foxp3+ Tregs positively correlated to the ALT and AST levels (12). These findings indicated that the upregulation of PD-L1 on Tregs or probably on other subsets, synergistically with the increase of PD-1 expression on T cells, was involved in the progression of HBV infection. Considering PD-L1 expressed on monocytes may also contribute to delivering a coinhibitory signal to T cells by engaging with PD-1 and, subsequently, associate with the progression of HBV infection; we further analyzed the correlation of PD-L1 expression on CD14+ monocytes of CHB, LC, and HCC patients with clinical parameters. Intriguingly, the results showed that the frequency of CD14+PD-L1+population in CHB was negatively correlated with HBV DNA load and ATL and AST levels, respectively (all p < 0.05) (Fig. 2A–C). However, no significant correlation between PD-L1+CD14+ population and such clinical parameters was observed in LC group (Fig. 2D–F). In HCC group, except that the frequency of PD-L1+CD14+ monocytes was positively correlated with HBV DNA load (p < 0.05) (Fig. 2G), there was no additional significant correlation presented (Fig. 2H, I).
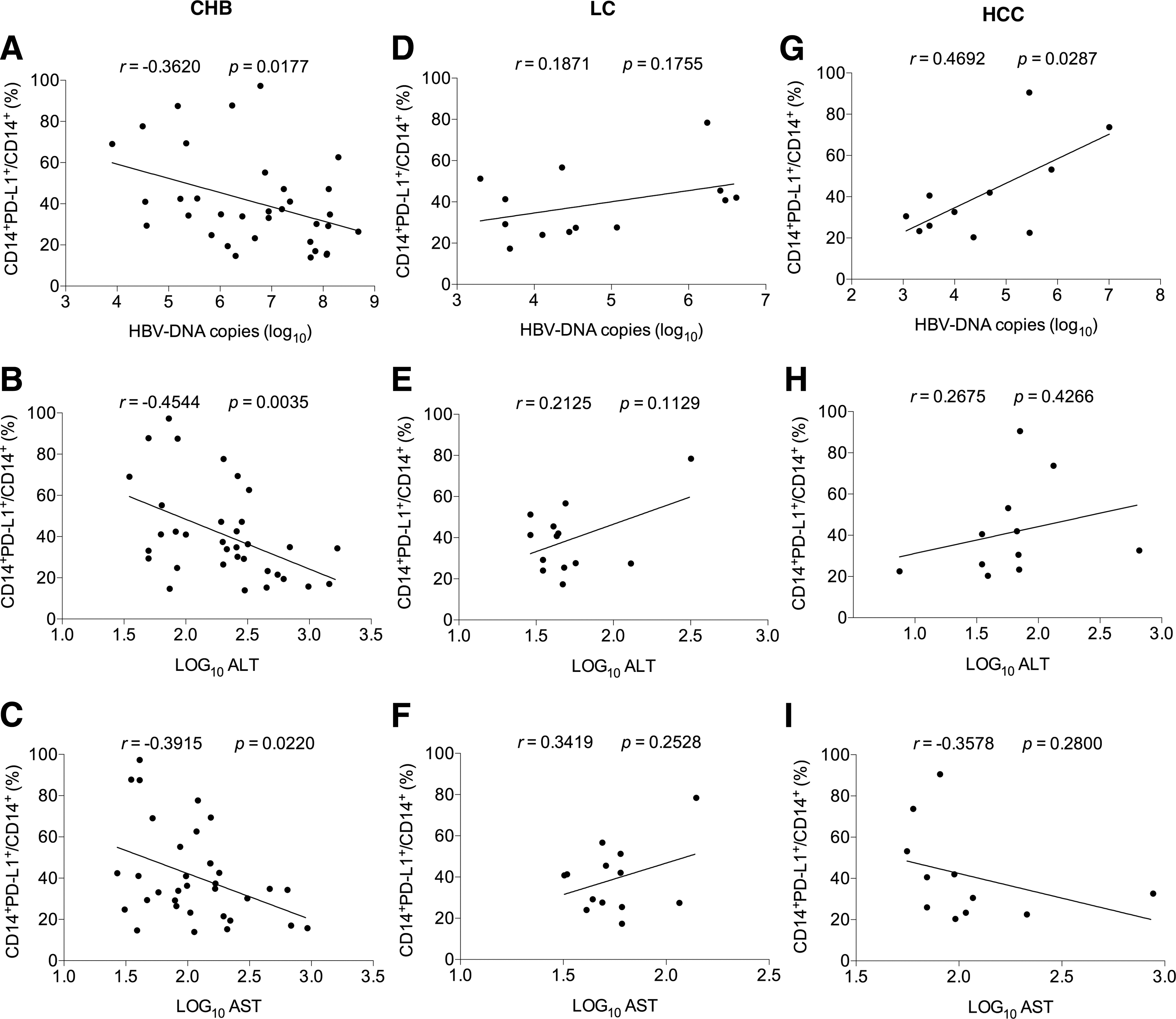
The correlation of the levels of PD-L1 expression on CD14+ monocytes with clinical parameters of CHB, LC, and HCC patients.
Investigation of the frequency of CD19+ B cells and PD-L1 expression on CD19+ B cells of CHB, LC, and HCC patients
B lymphocytes, known to be involved in humoral immune responses, play an important role in defense against viral infections. In the context of HBV infection, B cell responses are a T cell-dependent process and lead to an efficient antibody production in patients. At the same time, B lymphocytes can exert a function on T cells through cosignal molecules during cross talking. Therefore, PD-L1 on B cells may deliver a coinhibitory signal into T cells by engaging with PD-1 expressed on T cells and, subsequently, contribute to T cell exhaustion. For this reason, we investigated PD-L1 expression on CD19+ B cells from patients with HBV infection in the current study. Gating strategies for determining CD19+ B cells and PD-L1 on CD19+ B cell population were shown in Figure 3A. The results showed that the frequency of circulating CD19+B cells in HCC group was higher than that in HC group (p < 0.05) (Fig. 3B). At the same time, the absolute number of CD19+cells in CHB group was significantly higher than that in HC and LC groups (both p < 0.05) (Fig. 3C). In addition, it was found that the frequency of PD-L1+CD19+ population in CHB, LC, and HCC groups was all significantly elevated compared with that in HC group (all p < 0.001) (Fig. 3D). The frequency of PD-L1+CD19+cells among CHB, LC, and HCC groups had no significant difference (Fig. 3D).
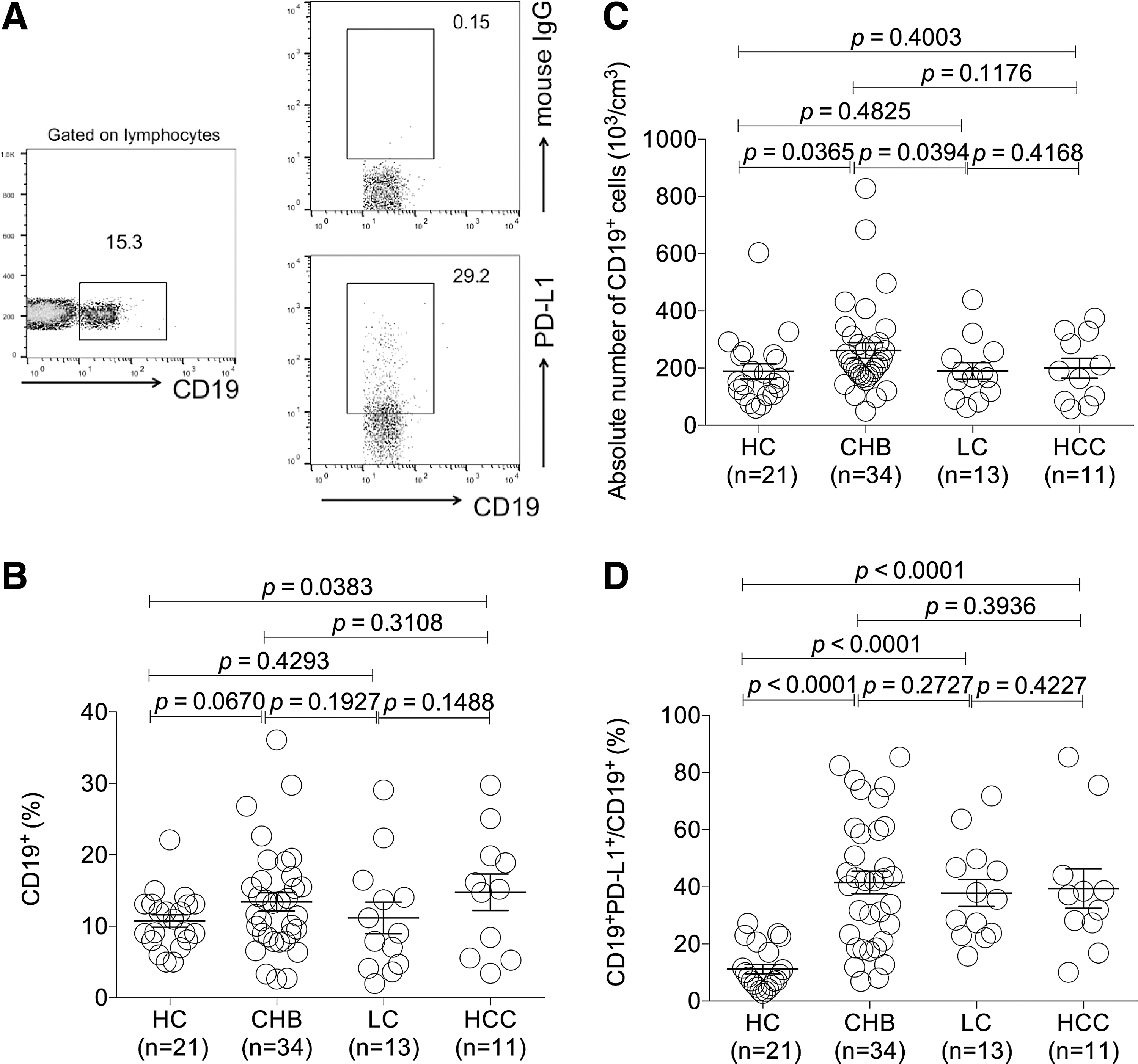
The expression profiles of PD-L1 on peripheral CD19+ B cells of CHB, LC, and HCC patients.
Analysis of the correlation of the levels of PD-L1 expression on CD19+ B cells with HBV DNA load and ALT and AST levels
T cell exhaustion owing to PD-1 on T cells engaging with PD-L1 on CD19+ B cells may be involved in the progression of HBV infection. Therefore, we further analyzed the correlation of the levels of PD-L1 expression on CD19+ B cells with clinical parameters. The results showed in CHB group that there was no significant correlation between the levels of PD-L1 expression on CD19+ B cells and HBV DNA load except that a positive correlation with AST levels (p < 0.05) and a tendency of positive correlation with ALT were observed (Fig. 4A–C). In LC group, although the frequency of CD19+PD-L1+ subsets did not present a correlation with HBV DNA load (Fig. 4D), it was correlated with ALT and AST levels, respectively, with a positive significance (both p < 0.05) (Fig. 4E, F). However, in HCC group, any correlation between the frequency of CD19+PD-L1+ and HBV DNA load and ALT and AST levels could not be found (Fig. 4G–I).
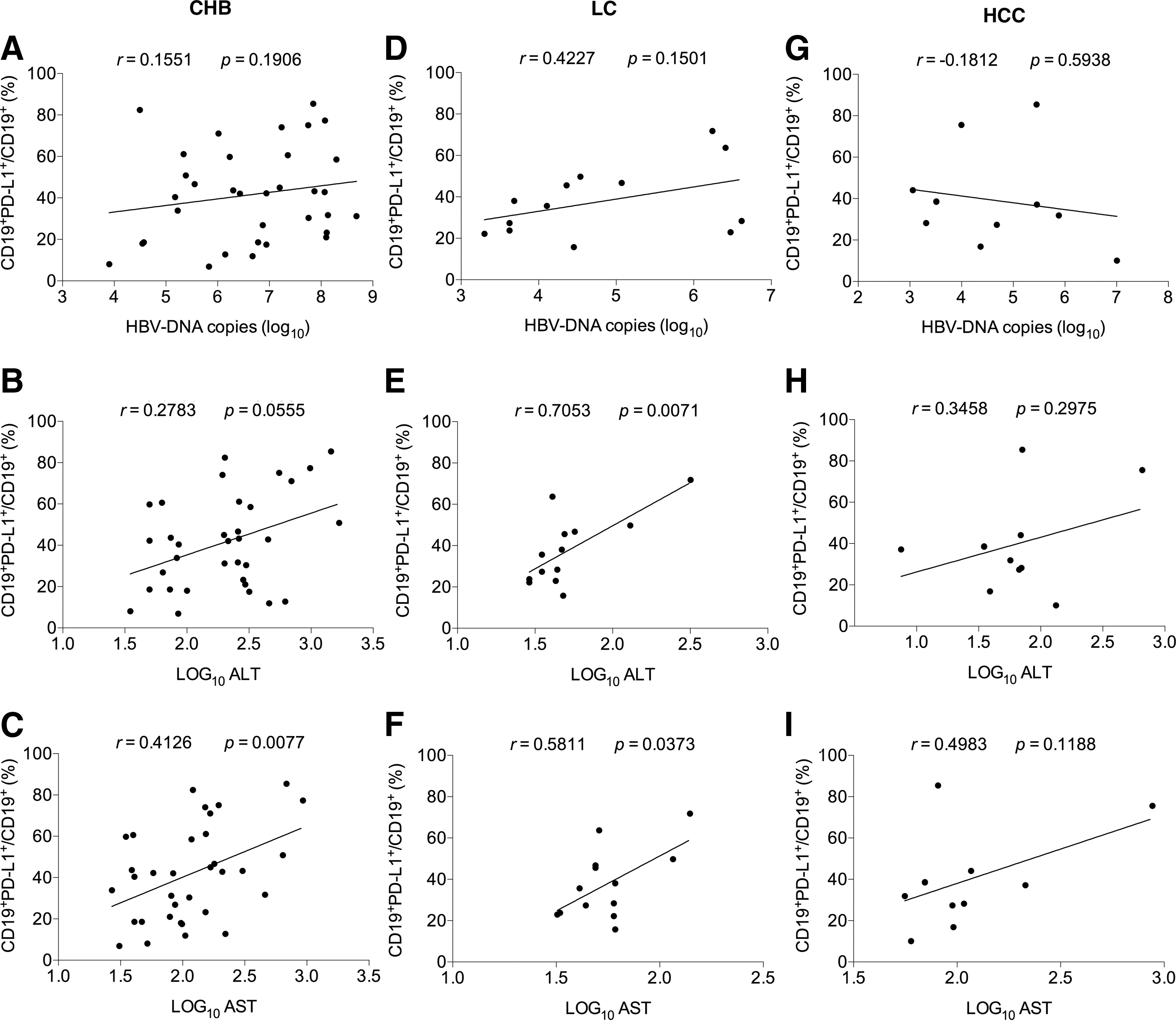
The correlation of the levels of PD-L1 expression on CD19+ B cells with clinical parameters of CHB, LC, and HCC patients.
Discussion
PD-1, dependent on ligation with its ligand PD-L1 or PD-L2, can transduce an inhibitory signal, resulting in immunocyte arrest (32). It has been well understood that the PD-1/PD-L1 pathway greatly contributes to inhibition of immune responses, and blockade of the interaction between them could remarkably restore the capacity of immune subsets against pathogen infection and tumor progression (5,21,26).
In HBV and HCV infection, direct activation of Kupffer cells driven by viruses and recruitment of other leukocytes and the subsequent immune responses are the main culprits for the pathogenesis of liver damage (16). Therefore, the activity and characteristics of these immunocytes are closely associated with the disease's progression. Among them, the activity of T cells during antiviral immune responses is greatly modulated by PD-1/PD-L1 axis. Our pervious study, as well as other reports, has clearly described that an increase of PD-1 expression on CD4+ and CD8+ T cells of CHB patients and on CD8+ T cells of HCC patients was positively related to HBV load and liver function (7,18,29). Moreover, upregulation of PD-L1 on Tregs, as well as monocytes, has also been reported during CHB and HCV infection (11,21,30). Thereby, blocking PD-1/PD-L1 interaction in the course of HBV infection could restore HBV-specific T cell function and the antiviral effect (28).
For a better understanding of the influence of PD-1/PD-L1 on the progression of HBV infection, a systematic investigation of PD-L1 expression pattern remains to be further elucidated during HBV infection from CHB to LC and then to HCC. Based on such consideration, we investigated the levels of PD-L1 expression on circulating CD14+ monocytes and CD19+ B cells of CHB, LC, and HCC patients.
First, our results demonstrated that expression of PD-L1 was upregulated on CD14+ monocytes of CHB patients. Such finding is consistent with previous reports (27,30), but different from other's discovery (21). The reasons might lie in subjects with a different range of viral load to be recruited in different studies since virus-derived antigen can induce PD-L1 expression (4), probably attributed to different samples with peripheral blood or liver tissues, and (or) was due to different detection methods with flow cytometry or immunohistochemistry to be used (21). At the same time, PD-L1 was also identified to increase on CD14+ monocytes of LC and HCC patients. Intriguingly, we found that the frequency of CD14+PD-L1+ population was negatively correlated with HBV DNA load and ALT and AST levels in CHB patients, while was not related to such clinical parameters except for being positively correlated with HBV DNA load at HCC stage. It is understandable that the increased inhibition of immune responses, which was mediated by the upregulation of PD-L1 combining with PD-1, relieved immunocyte-caused injury of hepatocytes at the stage of CHB (1,10). Thereby, we could find that along with PD-L1 upregulation, hepatocyte damage was alleviated, characterized by less ALT and AST levels at this stage. As for the inversed correlation between viral DNA load and the levels of PD-L1 expression on monocytes, one reason might be due to much amplified viruses driving more inflammation responses and then the inflammation cytokines such as macrophage colony-stimulating factor inducing a portion of new production of monocytes from bone marrow into blood, which expressed low levels of PD-L1 and had not been able to receive effective stimulation of HBeAg to upregulate PD-L1 expression (17) since almost half of patients were negative with HBeAg in their blood (Table 1). However, the mechanism underlying it remains to be explored. When HBV infection developed to LC and HCC, after persistent stimulation with HBV antigens, immune responses were exhausted due to upregulation of immune checkpoints such as PD-1/PD-L1, Treg-mediated inhibition, and so on. During these advanced stages, the proliferation and regeneration of immunocytes were also suppressed. Therefore we could find that at the stages of LC and HCC, the absolute number of CD14+ monocytes decreased compared with CHB stage. However, the frequency of monocytes in CHB patients was higher than that in HC, which probably attributed to the number of other immune subsets that decreased much more than monocytes (11). In addition, a positive correlation between the frequency of CD14+PD-L1+ population and HBV DNA load occurred at HCC stage since persistent stimulation with HBeAg antigen, which was positively detectable in most HCC patients (Table 1), could upregulate the expression of PD-L1 on monocytes.
Actually, CD14+ monocyte-expressed PD-L1 ligation with its receptor PD-1 on T cells has been reported to stimulate interleukin-10 production, suggesting that PD-L1 abnormal expression was involved in the progression of HBV infection by manipulating the immune responses toward Th2 dominance (3,6,10). In recent investigations of HBV and HCV-related patients, correlation was also found between PD-L1 expression on monocytes and liver inflammation. It was reported that PD-L1/CD86 ratio in the CD14++CD16+ monocyte correlated positively with the HCV load and HCV core antigen in the chronic HCV infection (33). And upregulation of PD-L1 on Kupffer cells and infiltrating monocytes/macrophages was closely associated with liver inflammation and antiviral immunity (4,24).These results further proved that the PD-1/PD-L1 pathway might contribute to the profile of immunosuppression observed in CHB, as well as LC and HCC, patients.
In addition, we verified that compared with HC group, PD-L1 was also upregulated on peripheral CD19+ B cells from patients of CHB, LC, and HCC. Further analysis revealed that PD-L1 expression was positively correlated with AST in both CHB and LC groups and with ALT in LC group, but was not related to HBV DNA load. In this study, we speculate that the underlying mechanism of PD-L1 upregulation on CD19+ B cells may be more complicated than that on CD14+ monocytes. As a kind of important adaptive subsets, B cells are also involved in antiviral immune responses, undergoing activation, differentiation, and remodeling. High expression of PD-L1 has been defined as an important mediator on regulatory B cells (Bregs), a subpopulation of B cells suppressing immune responses with similarities to Tregs. It has been reported that PD-L1 can be upregulated on Bregs by interferon (IFN)-γ (19). During HBV infection, increased levels of IFN-γ might dominantly promote PD-L1 expression on B cells other than HBeAg antigens, resulting in a negative correlation of it with viral DNA load. At the same time, the increase in levels of IFN-γ is synchronized to the activation and elevation of immune responses, which generally cause damage to liver. Hence, we could find a positive correlation between PD-L1 expression on B cells and the levels of ALT and (or) AST in CHB and LC patients, which is not indirect according to our above hypothesis. The significance of the increase of PD-L1-overexpressed B cells, including substantial numbers of Bregs, contributes to forming a negative feedback mechanism to protect liver from damage from excessive immune responses.
It was also reported that B cells could induce the suppression of HBV-specific CD8+ T cell responses, which could further constrain antiviral effect along with coinhibitory signals (5,8,25). Higher CD19+ PD-L1+ B cells showed the possibility that PD-L1 expression might be responsible for the progression of HBV infection through a direct suppression of HBV-specific CD8+T cells.
In conclusion, an increase in the frequency of circulating CD14+PD-L1+monocytes, as well as CD19+PD-L1+ B cells, was observed in CHB, LCC, and HCC patients and was correlated with some clinical parameters during different stages. Since antiviral chemicals against CHB infection can suppress HBV replication, but not eradicate HBV infection in the majority of cases, further investigation still needs to be done to better understand the immune-mediated pathogenesis of HBV infection for adopting the promising immunotherapy. Our findings of dynamic variations of PD-L1 on monocytes and B cells and the correlations with clinical parameters during the progression of HBV infection from CHB to LC and then to HCC not only make PD-1/PD-L1 itself another valuable clinical parameter but also provide valuable information to prevent the risk of CHB patients from developing LC and liver cancer and develop potential therapeutic strategies of PD-1/PD-L1 blockade against HBV infection.
Footnotes
Acknowledgments
This research was supported by the National Natural Science Foundation of China (31570889, 31320103918), a project Funded by the Priority Academic Program Development of Jiangsu Higher Education Institutions, 973 Program (2013CB530501), the Project of Scientific and Technical Development of Suzhou (SYS201101, SS201664, SZS201412, SYS201433, SYS201155, and SYS210424), and Jiangsu Province's Key Laboratory of Medicine (XK201135).
Author Disclosure Statement
No competing financial interests exist.