
Abstract
Marek's disease virus (MDV), a highly cell-associated lymphotropic α-herpesvirus, is the causative agent of Marek's disease (MD) in domestic chickens. MDV replicates in chicken cells and establishes a latent infection within CD4+ T cells. Although MD vaccines have been in use for several decades, the exact mechanism of vaccine-induced protection is unclear. It is believed that the innate immune system plays a role in vaccine-induced immunity against pathogenic strains of MDV. To shed light on the possible function of the innate immunity in vaccine-mediated protection, we investigated the effect of vaccination, Rispens/CVI988, on the activation of cellular components of the innate immune system by analyzing the expression pattern of select immune-related genes in the cecal tonsils (CT) and duodenum of two MD-susceptible and MD-resistant chicken lines at 3, 5, and 10 days postvaccination (dpv). The differential expression patterns of the tested genes within the CT and duodenum of vaccinated birds revealed the activation of the innate immune system in both the susceptible and resistant lines. Stronger innate immune response was induced within the CT of the vaccinated birds of the susceptible line at 5 dpv. Upregulation of some of the tested genes at 10 dpv was likely due to the activation and response of the adaptive immune system to vaccination. Immunohistochemical analysis showed no increase in the number of CD3+ T cells in the CT and duodenum of the vaccinated birds of either line at 5 dpv. There was, however, an increase in the macrophage populations within the duodenum of the vaccinated birds of both the susceptible and resistant lines at 5 dpv. The vaccine strain antigen was detected in the CT and duodenum of the susceptible line, but not the resistant line at 5 dpv.
Introduction
M
Macrophages, natural killer (NK) cells, cytokines, chemokines, and cytotoxic T lymphocytes play crucial roles in MDV pathogenesis and resistance or susceptibility to MD (1). Studies have shown that depletion of macrophages before MDV infection or continuous depletion during the course of infection results in severity of MD-related complications and increases incidence of tumors (21,22). It has also been shown that activated macrophages from MDV-infected chickens produce NO that inhibits viral replication (16,39).
NK cells also have been shown to play a critical role in controlling herpesvirus replication and infection (14,30). NK cells are a subset of cytotoxic lymphocytes that mediate non-MHC-restricted cytotoxicity against tumor and virally infected cells (10,33). The killing of infected and tumor cells is mediated by cytotoxicity and cytokine production (5,9,51). The potential role of these essential cellular components of the innate immune system in MDV pathogenesis has been confirmed. Sharma and Coulson (42) have reported NK cell cytotoxic activities against MDCC-MSB1 cells. In addition, it has been shown that MD-resistant chickens have a higher NK cell activity than the susceptible birds. The reduced biological activities of NK cells were more pronounced in the susceptible line developing MDV-induced tumors (26,41). Moreover, studies have shown that the level of NK cell activity increases shortly after MD vaccination (25,43).
MDV strains are categorized into three pathotypes based on genomic differences and biological features. Oncogenic MDV strains, including their attenuated forms, are classified as serotype 1. Serotypes 2 and 3 are nononcogenic viruses isolated from chickens and turkeys, respectively (28,39). Nononcogenic strains of serotypes 2 (SB-1) and 3 (HVT) and the attenuated serotype 1 (CVI988/Rispens) have been used as vaccines to control MD since 1968 (34,37,40,49). CVI988/Rispens, a naturally attenuated serotype 1, is the most effective vaccine used against highly pathogenic strains of MDV (37). Live MDV vaccination prevents lymphoma formation and a significant reduction in virus replication in the FFE but not host infection, viral replication, and shedding (3,8,23,31,50).
The exact mechanism of vaccine-induced protection is unknown. It is speculated that the innate immune system plays a role in vaccine-induced immunity against pathogenic strains of MDV. It is believed that shortly after vaccination, activated NK cells produce IFN-γ and destroy infected B cells. IFN-γ inhibits viral replication and activates macrophages leading to production of NO that has direct inhibitory effect on viral replication and infection (39). The antigenic similarity between the vaccine and pathogenic strains leads to stimulation of immune responses against the virulent viruses and consequently, prevention of virus replication and reduction in viremia (39).
Materials and Methods
Experimental chickens
The specific pathogen-free chickens in this study were from two highly inbred lines of 63 (MD-resistant) and 72 (MD-susceptible) birds (2). These birds were from unvaccinated breeder hens and carried no maternal antibodies to MDV or herpesvirus of turkeys and were not vaccinated posthatch. The chicks were hatched at Avian Disease and Oncology Laboratory (ADOL) poultry facility and housed in modified Horsfall-Bauer isolation units for the duration of the experiment. All animal experiments were approved and carried out in accordance to the guidelines set forth by the ADOL Institutional Animal Care and Use Committee and the Guidelines for Care and Use of Laboratory Animals published by Institute for Laboratory Animal Research (ILAR Guide) in 1996 (
Virus
CVI988/Rispens, a commercially available MD vaccine, was used in this study.
Immunohistochemistry
Samples previously flash frozen in an embedding medium at an optimal cutting temperature (OCT) (Sakura Finetek, Torrance, CA) were sectioned on a cryotome at 5 microns and placed on slides coated with 2% 3-Aminopropyltriethoxysilane and air dried at 25°C overnight. Subsequently, microtome sections were fixed in formal acetate fixative for 10 min at room temperature followed by three changes of Tris-buffered saline (TBS)—5 min each. The endogenous peroxidase activity was blocked with 0.3% hydrogen peroxide in TBS for 20 min followed by tap and distilled water rinses. Following pretreatment standard, Avidin–Biotin complex staining steps were performed at room temperature on the DAKO Autostainer (Agilent Technologies, Carpentaria, CA). All staining steps were followed by rinses in TBS + Tween 20 (Scytek Laboratories, West Logan, UT). After blocking for nonspecific protein with normal horse serum (1/30 dilution in phosphate-buffered saline; Vector Labs, Burlingame, CA) for 30 min, sections were incubated with Avidin/Biotin blocking system for 15 min each (Avidin D–Vector Labs; d-Biotin–Sigma) (Sigma, St. Louis, MO). Samples were then incubated with mouse anti-chicken macrophage or CD3+ T cells monoclonal antibodies (KUL01, CT-3, respectively; SouthernBiotech, Birmingham, AL) for 1 h in Normal Antibody Diluent (NAD) (Scytek) followed by rinsing and incubation with biotinylated horse anti-mouse IgG (H + L) prepared at 11.5 μg/mL in NAD incubated for 60 min. For detection of CVI988/Rispens antigen, mouse anti gB monoclonal antibody (28) was used. Samples then were incubated with R.T.U. Vector Elite Peroxidase Reagent (Vector) for 30 min. The reaction development utilized Vector Nova Red peroxidase chromogen incubation of 15 min followed by counterstain in Gill Hematoxylin (Thermo Fisher, Kalamazoo, MI) for 15 sec, differentiation, and dehydration, clearing and mounting with a synthetic mounting medium. The working dilution for the monoclonal antibody specific for Rispens gB antigen was 1:1,000.
RNA isolation
Total RNA was isolated from the homogenized cecal tonsils (CT) or duodenum tissues of three birds of each group (see Experimental Design) at 3, 5, and 10 days postinoculation (dpi; three biological replications) using Tri Reagent RT (Molecular Research Center, Inc., Cincinnati, OH) according to the manufacturer's instructions.
Real-time reverse transcriptase-polymerase chain reaction
Real-time polymerase chain reaction (PCR) analysis of the relative quantification of chicken gene expression transcripts was carried out at the Research Technology and Support Facility of Michigan State University in East Lansing, Michigan. Briefly, 2.5 μL of a 1:20 dilution of the oligo dT-based RT product from 4 μg of total RNA isolated from the tissues was used for each reaction. Three hundred nanomolar of each specific sense and antisense primers was used in the presence of 5 μL SYBR Green PCR master mix (Applied Biosystems, Foster City, CA). The amplification program was as follows: 50°C for 2 min, 95°C for 10 min, 40 cycles at 95°C for 15 sec, followed by 57°C for 1 min. All the reactions were run in triplicates in a 7900HT Sequence Detection System (Applied Biosystems). The primers for chicken genes were designed using MacVector software (Accelrys, San Diego, CA). All the primers were synthesized by Operon Biotechnologies, Inc. (Huntsville, AL). With the exception of CCL-5 (RANTES), all the primer sequences have been published previously (24). The CCL-5 primers are as follows: forward 5′-TTGGGGCTGATACAACCGTG-3′ and reverse 5′-TTCCTGGTGATGAACACAACTGC-3′. Relative quantification of the chicken genes was determined using 2−ΔΔCT method (32). The levels of gene expression in the CT or duodenum tissues of age-matched control birds were used as reference or baseline for calculation of fold changes in gene expression in the CVI988/Rispens-vaccinated chickens of each line. The expression of each gene was normalized to the expression level of the housekeeping gene, β-actin.
Experimental design
One-day-old chicks from each line were randomly distributed into two groups of 25 birds each in separate isolators. Birds from one group of each line were inoculated intraperitoneally with 2,000 pfu of CVI988/Rispens at 12 days posthatch. The second group of each line served as uninoculated negative control. At 3, 5, and 10 days postvaccination (dpv) three birds from each group were euthanized by CO2 inhalation and necropsied for tissue collection. One of the CT of each bird from each group was pooled for RNA isolation and immediately stored in RNAlater (Ambion, Austin, TX) to prevent RNA degradation. The other CT was pooled into an OCT medium, snap-frozen in liquid nitrogen, and stored in −80°C until used for immunohistochemical analysis. Sections of each duodenum tissue were also stored in RNAlater for RNA isolation and snap-frozen in liquid nitrogen for immunohistochemistry.
Results
Immunohistochemistry
An immunohistochemical analysis of CD3+ T cell populations in the duodenum at 5 dpv revealed no visible increase within the tested tissues of the vaccinated birds of either line (Fig. 1B and D, lines 63 and 72, respectively) when compared to the control birds (Fig. 1A and C, lines 63 and 72, respectively). Although no specific software was used in determining the accurate cell populations within the tested tissues, there appeared to be an increase in the number of macrophages migrating into the duodenum of the vaccinated birds of both lines (Fig. 1F and H, lines 63 and 72, respectively), in comparison to the control tissues (Fig. 1E and G, lines 63 and 72, respectively). Staining for the gB glycoprotein of CVI988/Rispens in the tissues of the vaccinated birds detected no virus-specific antigen within the CT or duodenum of the vaccinated birds of the resistant line 63 at 5 dpv (Fig. 2B and D, CT and duodenum, respectively). Figure 2A and C represent the corresponding control tissues for line 63. There were, however, a significant number of virus particles detected within the CT and duodenum of the vaccinated birds of the susceptive line 72 (Fig. 2F and H, CT and duodenum, respectively). Figure 2E and G are the corresponding control tissues for line 72.
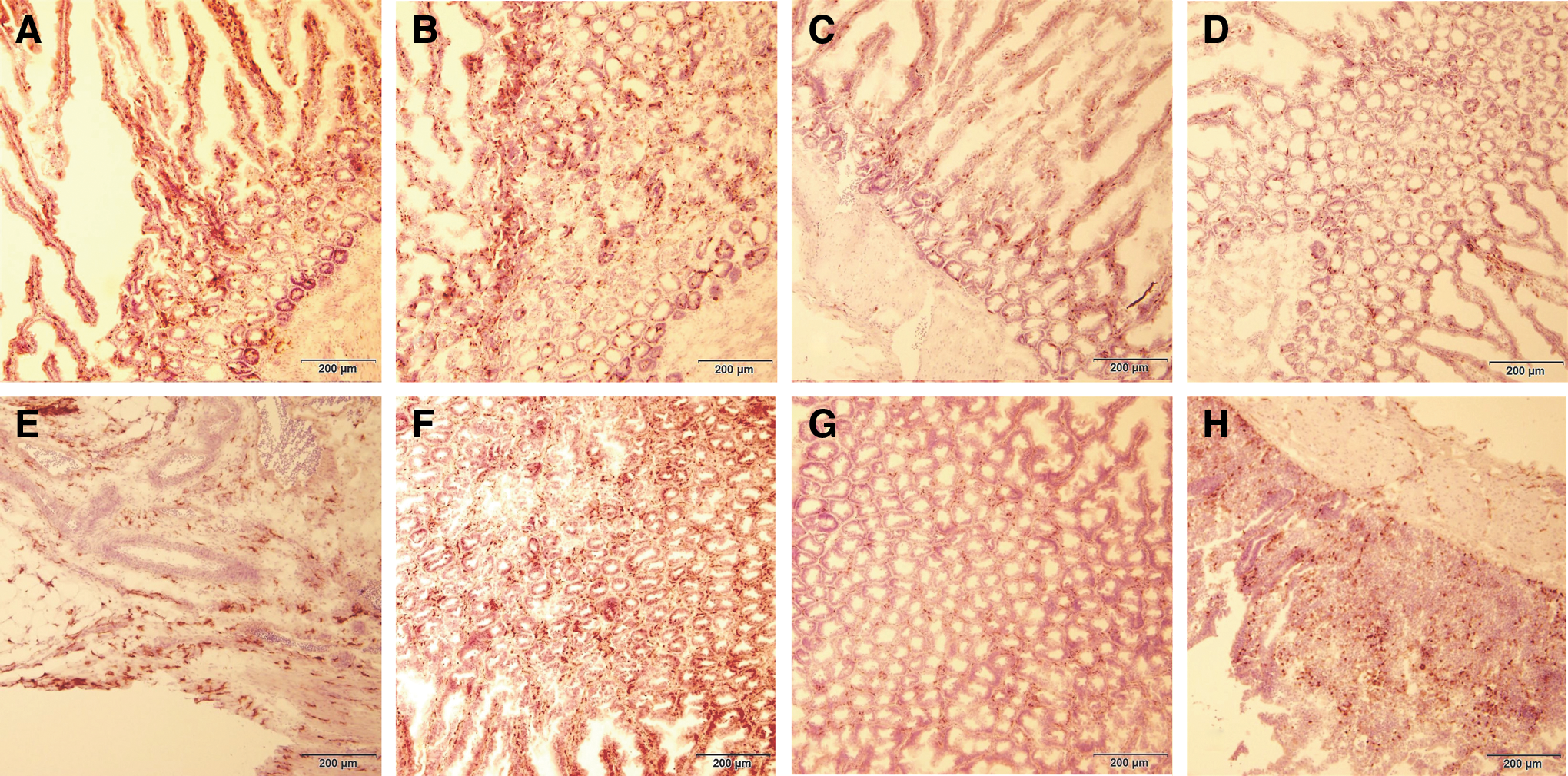
The figure depicts the immunohistochemical analysis of CD3+ T cells and macrophage populations within the duodenum of the control and vaccinated birds of the resistant and susceptible lines at 5 dpv.

The figure represents the immunohistochemical analysis of CVI988/Rispens antigen, gB, within the CT and duodenum of the vaccinated birds of the resistant and susceptible lines at 5 dpv.
Gene expression analysis
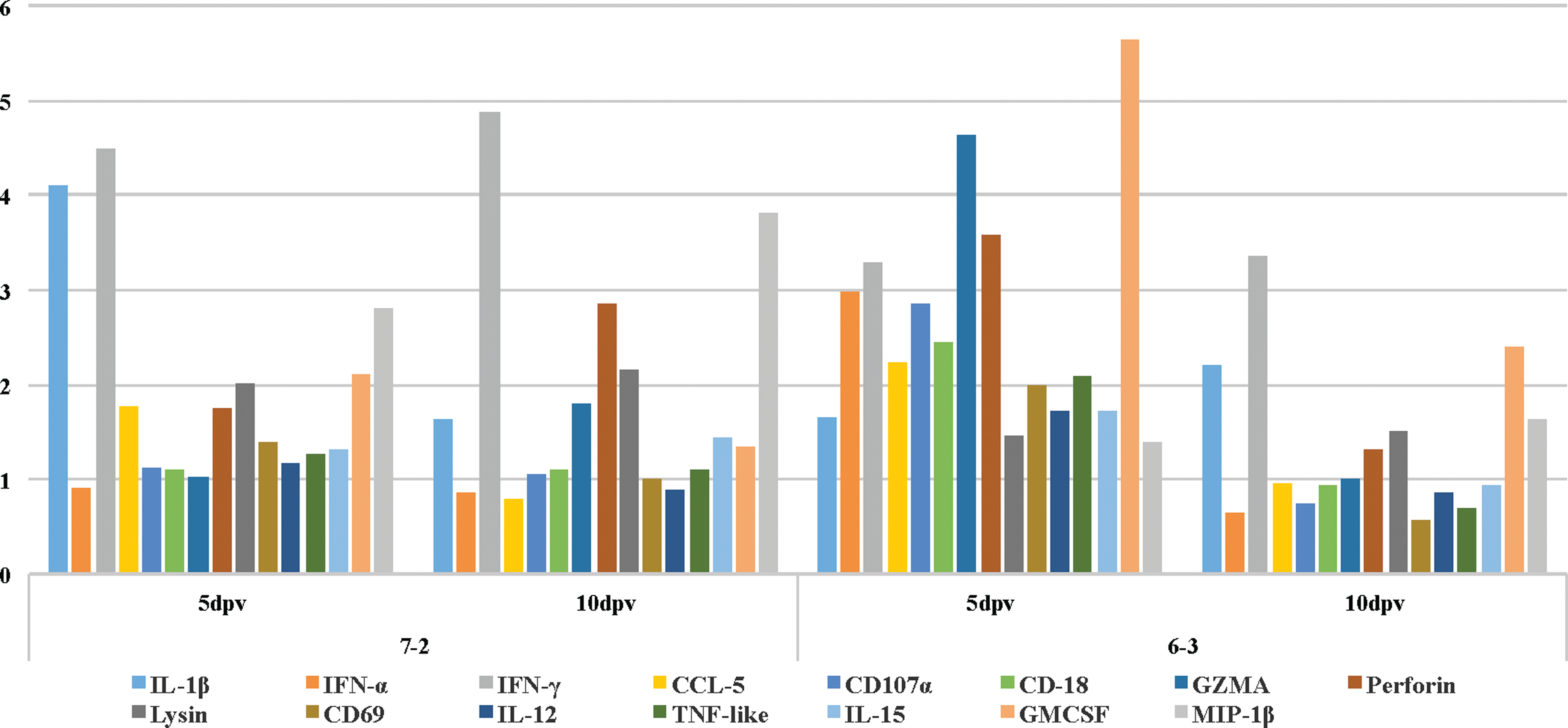
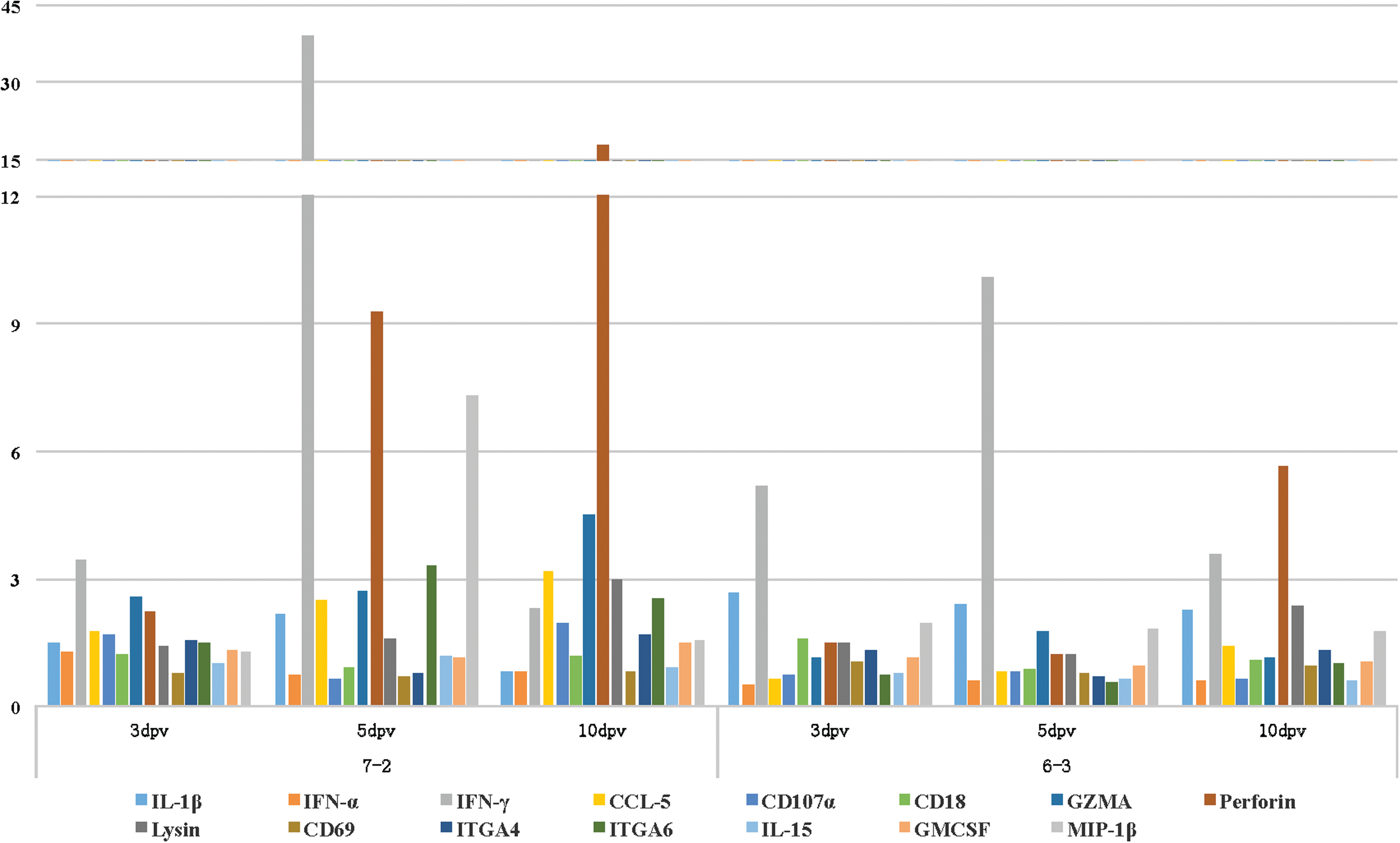
Real-time PCR-based gene expression analysis of a select number of immune-related genes within the duodenum of the vaccinated birds of either line showed no noticeable changes in the expression levels of the tested genes at 3 dpv when compared to the age-matched control birds (data not shown). In comparison to the control birds, the expression levels of IL-1β, IFNγ, and MIP-1β were all upregulated within the duodenum of the vaccinated birds of the susceptible line 72 at 5 dpi (Table 1). In the duodenum of the vaccinated birds of the resistant line 63, there was an increase in the expression levels of IFN-α, IFNγ, CD107a, granzyme-A (GZMA), perforin, and GM-CSF (Table 2). In addition to perforin, the expression levels of IFNγ and MIP-1β were also increased within the duodenum of the vaccinated birds of the susceptible line 72 at 10 dpv (Table 3). IFNγ and GM-CSF were also upregulated within the duodenum of the vaccinated birds of line 63 at 10 dpv (Table 4). The bar graph in Figure 3 depicts the expression pattern of all the tested genes at 5 and 10 dpv within the duodenum of the vaccinated birds of both the resistant and the susceptible lines. The transcriptional activities of IFNγ and GZMA were increased within the CT of the vaccinated birds of the susceptible line 72 at 3 dpv (Table 5). IL-1β and IFNγ were the only two cytokines that were upregulated within the CT of the resistant line 63 at 3 dpv (Tables 6). There was an increase in the expression level of IFNγ, CCL5, GZMA, perforin, ITGA6, and MIP-1α within the CT of the vaccinated birds of the susceptible line at 5 dpv (Table 7). Again, IL-1β and IFNγ were the only two cytokines that were upregulated within the CT of the resistant line 63 at 5 dpv (Table 8). At 10 dpv, IFNγ, CCL5, GZMA, perforin, NK-lysin, and ITGA6 were all upregulated within the CT of the susceptible line 72 (Table 9). In addition to IL-1β and IFNγ, there was an increase in the expression level of perforin and NK-lyisn at 10 dpv within the CT of the vaccinated birds of the resistant line 63 (Table 10). Figure 4 helps with the visualization of the expression pattern of all the tested genes within the CT of the vaccinated birds at all three time points.
dpv, days postvaccination; SD, standard deviation.
CT, cecal tonsils.
Discussion
MD is a common lymphoproliferative and neuropathic disease of domestic chickens that has caused significant economical losses to the poultry industry. MD is the first neoplastic disease that has been successfully controlled by vaccination (12). Vaccination could prevent tumor formation, transient paralysis, and lymphoid organ atrophy in the susceptible chickens, but not MDV infection and transmission (15,18). Evolutionary trend of MDV toward greater virulence with repeated outbreaks, however, has been a major concern for the poultry industry (17,48). Deciphering the molecular mechanism of vaccine-induced protection is essential for developing new and more efficacious vaccines that would induce sterile immunity and prevent virus dissemination, evolution, and future outbreaks.
Studies have shown that unlike most human and animal vaccines that require activation of the adaptive immune system for protection, MD vaccines induce partial immunity within 24–48 h after vaccination. The protection efficacy of the vaccines reaches 97–100% by 5 dpv (29). In contrast to the adaptive immune system that takes days for activation, the innate immune responses are rapidly induced, which play a critical role in controlling the initial events in viral infection (16). NK cells are the major cellular components of the innate immune system that are likely involved in vaccine-mediated protection (16,41). NK cell-induced apoptotic death of target cells is mediated by exocytosis of perforin and granzymes and production of IFN-γ that has direct affect on the outcome of adaptive immune response that is critical for an effective defense against virally infected and tumor cells (5,9,47,51).
The focus of this study was to investigate the effect of MD vaccination on the activation of NK cells within the gut-associated lymphoid tissues (GALT) by analyzing the expression profiling of a panel of immune-related genes in the CT and duodenum of two MD-susceptible and MD-resistant chicken lines at three different time points postvaccination. GALT are heavily laden with NK cells, macrophages, and other cell types that participate in immune responses against bacterial and viral pathogens (20,27). In contrast to mammals, the intestinal intraepithelial leukocytes of chickens comprise a large population of NK cells, a rich source for isolation and functional analysis (20).
Gene expression analysis revealed that there were no significant changes in the expression levels of the examined genes at 3 dpv within the duodenum of the vaccinated birds of either line when compared to the age-matched unvaccinated control birds (data not shown). At 5 dpv, however, there was an increase in the expression level of IL-1β, a proinflammatory cytokine, IFN-γ, a cytokine mainly produced by activated NK cells in response to viral infection (36), and MIP-1β within the duodenum of the vaccinated birds of the susceptible line 72 (Table 1). MIP-1β is a chemotactic cytokine produced by activated macrophages and NK cells (35,45). In the duodenum of the vaccinated birds of the resistant line 63, in addition to an increase in the expression level of IFN-α, there was an upregulation in the expression levels of IFN-γ, CD107a, GZMA, perforin, and GM-CSF, all expressed by activated NK cells (13,19,35,47) (Table 2). At 10 dpv, which coincides with the activation of the adaptive immune system, there was an upregulation in the expression of IFN-γ, perforin, and MIP-1β within the duodenum of the vaccinated birds of the susceptible line 72 (Table 3). The activation of the NK cells within the duodenum of the vaccinated birds of the resistant line was extended to 10 dpv, as was evident by the higher expression levels of IFN-γ and GM-CSF in comparison to the age-matched control birds (Table 4).
Unlike the duodenum, there were higher transcriptional activities for some of the tested genes within the CT of the vaccinated birds at 3 dpv. IFN-γ and GZMA were upregulated within the CT of the vaccinated birds of the susceptible line 72 (Table 5). In addition to IFN-γ, the expression of IL-1β was increased within the CT of the vaccinated birds of the resistant line 63 (Table 6). At 5 dpv, however, several genes, including IFN-γ (39-fold increase), GZMA (2.7-fold increase), perforin (9.3-fold increase), and MIP-1β (7.3-fold increase), all directly associated with the activation of NK cells, were upregulated in the CT of the vaccinated susceptible line 72 (Table 7). In addition, ITGA6, a member of integrins α6 adhesion molecules that mediate cell adhesion to laminin in an activation-dependent manner (44,46), showed higher expression level in the same vaccinated birds. Unlike the susceptible line, there were only two genes IL-1β and IFN-γ that exhibited higher transcriptional activities within the CT of the vaccinated chickens of the resistant line 63 (Table 8). At 10 dpv, the expression levels of GZMA, perforin, NK-lysin, and ITGA6 were still higher in the CT of the vaccinated birds of the susceptible line 72 (Table 9). It should be noted that it is likely that at 10 dpv, the adaptive immune system is activated and the cytotoxic T cells are partially contributing to the higher expression levels of GZMA, perforin, and other genes. Like the susceptible line, higher expression levels for IFN-γ, perforin, and NK-lysin were detected within the CT of the vaccinated resistant line (Table 10).
The expression patterns of all the tested genes within the CT and the duodenum of the vaccinated birds are depicted as bar graphs for easier visualization in Figures 3 and 4, respectively.
Immunohistochemical analysis of CD3+ T cells and macrophages in the tissues of the control and vaccinated birds revealed no discernible increase in the population of T cells within the duodenum of the vaccinated birds of either line at 5 dpv, suggestive of lack of activation of adaptive immune system (Fig. 1A–D). There was, however, an increase in the population of macrophages in the duodenum of the vaccinated birds of both susceptible and resistant lines (Fig. 1E–H). There were not enough CT tissues to test for population of CD3+ T cells or macrophages at either 3 or 5 dpv. In addition, immunohistochemical analysis detected no CVI988/Respins antigen within the CT or duodenum of the vaccinated birds of the resistant line at 5 dpv. A significant number of virus particles, however, were detected within the CT and duodenum of the susceptible line (Fig. 2A–H). Considering the marked increase in the expression levels of some of the tested genes associated with activation of macrophages and detection of a substantial number of virus particles at 5 dpv in the duodenum of the vaccinated susceptible line, it is safe to speculate that the inhibitory effect of MDV on macrophage function has resulted in the lack of activation of NK cells and elimination of virus particles. The absence of any detectable CVI988/Rispens antigen within the tested tissues of the resistant line, on the other hand, is probably an indication that the virus was unable to break the macrophage barrier that resulted in activation of NK cells and ultimately, clearance of viral infection.
In summary, MD vaccination activates NK cells of the innate immune system, which shapes the emerging adaptive immune system that is so critical for an effective defense against viral infection.
Footnotes
Acknowledgment
The authors thank Jennifer Pierluissi for excellent technical assistance.
Author Disclosure Statement
No competing financial interests exist.