
Abstract
While upregulation of 2,3-dioxygenase (IDO) accompanied by degradation of tryptophan along the kynurenine pathway have been reported to exert antimicrobial effects against a wide range of infectious agents, its role in the replication of influenza A virus remains uncertain. We performed experiments using influenza A/WSN/33 virus infection of mouse fibroblast cell-line (NIH-3T3) to study the effects of IDO on viral replication. Influenza infection resulted in prominent elevations of transcripts encoding IDO, interferon (IFN)-β, and segment 8 of the virus in NIH-3T3 cells. Introduction of siRNA targeted against IDO followed by infection resulted in further increased levels of viral RNA without altering IFN-β expression. Inhibition of IDO during the infection also resulted in reduction of virus-driven upregulation of 3-hydroxyanthranilate 3,4-dioxygenase (HAAO), but not kynurenine 3-monooxygenase (KMO), which are enzymes downstream in the kynurenine pathway. Thus, induction of IDO appears to contribute to limiting replication of the WSN/33 strain of influenza A virus in murine NIH-3T3 cells.
I
Interestingly, in vivo studies indicated that tryptophan deprivation and the accumulation of tryptophan metabolites along the kynurenine pathway had important immune modulatory effects, including attenuation of T cell responses, reviewed in (10). Yoshida et al. reported that an experimental influenza A virus infection resulted in an 120-fold increase in IDO activity in lungs of infected mice (16). Similar observations were made more recently by Huang et al. who reported greatly increased levels of tryptophan metabolites in lung tissue of Balb/c mice following influenza A/PR/8/34 virus infection (9). Interestingly, ablation of Indo in such mice did not increase viral loads but conferred reduced mortality, enhanced CD8 responses, and earlier recovery of infected animals (9). More recently, Fox et al. reported that pharmacological inhibition of IDO by 1-methyl-D, L-tryptophan (1MT) during influenza A virus infection resulted in enhanced expression of proinflammatory genes, including IFN-β during the early stages of infection both in vivo and in vitro (6). The effect of IDO inhibition on virus replication was not reported. This group of researchers also reported that infection of lung respiratory epithelial cells with the influenza A/WSN/33 virus strain caused an upregulation of Indo transcripts that appeared to be driven by induction of both type I and III IFNs. Interestingly, pharmacological inhibition of IDO, again by 1MT, during this in vitro infection resulted in decreased viral titers (5). Thus, while IDO appears to have antireplicative activity against a range of infectious agents its activity against the single-stranded RNA virus influenza A remains obscure.
IDO could be induced by the mouse-adapted neurotropic influenza A/WSN/33 virus strain not only in respiratory epithelium but also in murine neurons and astroglia (7). To determine whether IDO is induced also in fibroblasts in response to influenza A/WSN/33 virus infection, we initially examined the levels of transcripts encoding mouse IDO in the mouse embryo fibroblast cell-line (NIH-3T3) following infection with influenza A/WSN/33 virus. Twenty-four hours after inoculating cells with 0.5 multiples of infection of virus, as previously described (3), we observed marked elevations of IFN-β transcripts and viral RNA as determined by real-time polymerase chain reaction (ABI 7000; Applied Biosystems, Palo Alto, CA) using random-primed cDNA (Invitrogen, Carlsbad, CA) generated from total cellular RNA (RNeasy mini kits; Qiagen, Hilden, Germany), suggesting an efficient virus infection and replication along with induction of an antiviral response. Normalized to the levels of GAPDH transcripts, significantly higher levels of Indo transcripts (24.7- to 64.1-fold) were observed in infected cells as compared to uninfected cells (Fig. 1).
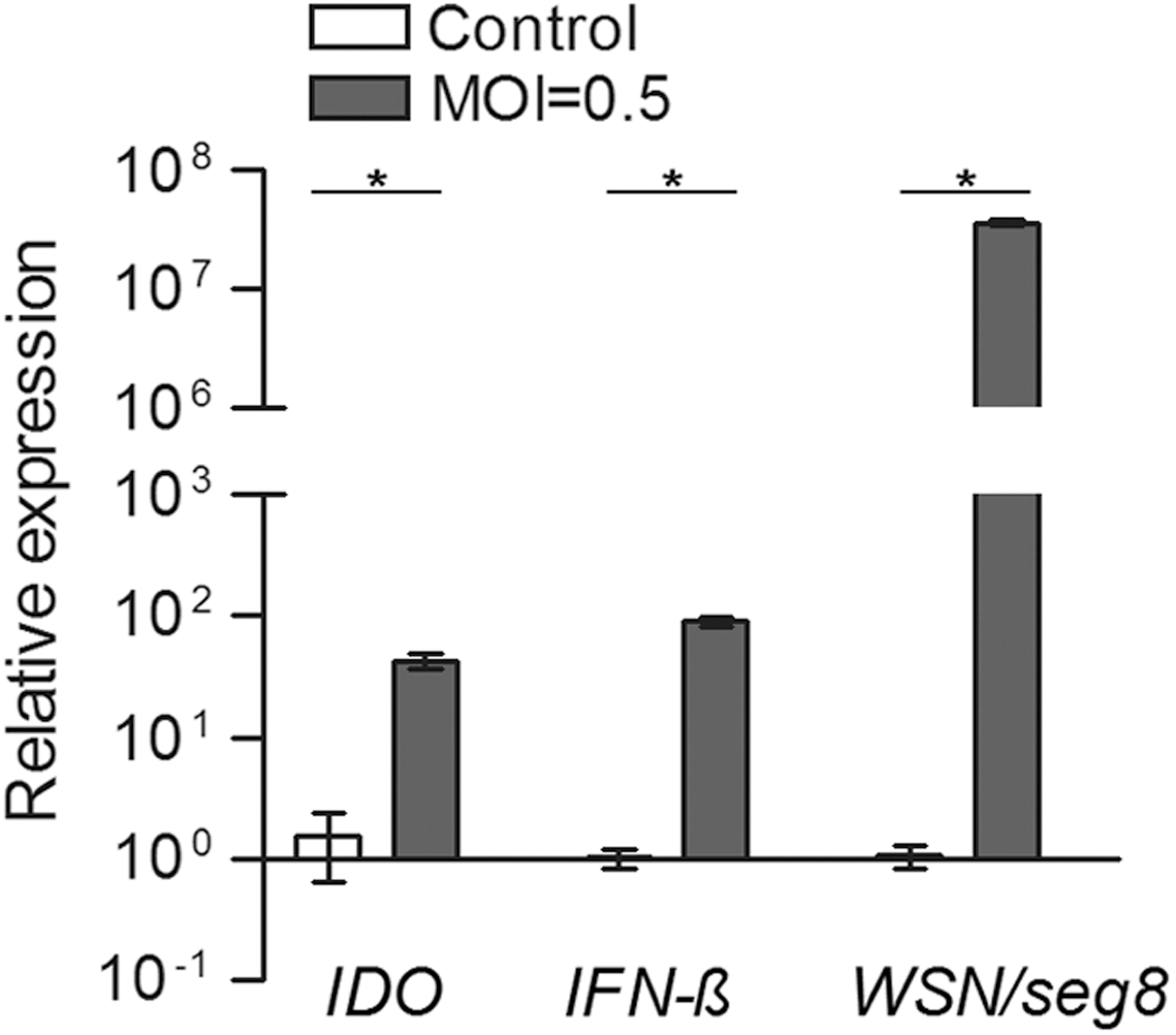
Relative expression levels of transcripts encoding IDO, IFN-β, and segment 8 of the influenza A/WSN/33 virus in NIH-3T3 cells in response to influenza A/WSN/33 virus infection. Comparison between groups analyzed by Mann–Whitney U test. *p < 0.05. IDO, 2,3-dioxygenase; IFN, interferon; MOI, multiplicity of infection.
To investigate whether inhibition of IDO during virus infection influences viral replication, a synthesized siRNA pool against Indo transcripts was applied in the subsequent experiments. NIH-3T3 cells transfected with 300 ng siRNA pool targeted against IDO (Dharmacon, Diegem, Belgium) for 24 h followed by another 24 h influenza A/WSN/33 infection exhibited significantly lower levels of transcripts encoding IDO as compared to infected cells transfected with scrambled siRNA (Fig. 2A). Such cells harbored significantly increased levels of segment 8 of the influenza A virus genome but similar levels of transcripts encoding IFN-β compared to infected cells transfected with scrambled siRNA, Fig. 2B.

Relative expression of transcripts encoding
We previously reported that transcripts encoding not only IDO but also other downstream enzymes in the kynurenine pathway, such as kynurenine 3-monooxygenase (KMO, catabolizes kynurenine to 3-hydroxykynurenine) and 3-hydroxyanthranilate 3,4-dioxygenase (HAAO, catabolizes 3′hydroxyanthranilic acid to quinlinic acid), were increased several fold during influenza A/WSN/33 infection. We therefore took this opportunity to investigate the degree to which transcription of the genes encoding KMO and HAAO depend on IDO expression levels. NIH-3T3 cells transfected with siRNA targeted against IDO followed by influenza A/WSN/33 infection exhibited significantly lower levels of transcripts encoding HAAO compared to infected cells treated with scrambled oligonucleotides. The levels of transcripts encoding KMO did not differ between the two groups (Fig. 2A).
Here, we report induction of transcripts encoding IDO in a mouse fibroblast cell-line in response to influenza A virus infection. siRNA-mediated attenuation of this induction resulted in larger numbers of viral RNA molecules, suggesting that IDO activity potentially contributes to limiting replication of influenza A virus in vitro. As described above, this observation is in line with previous studies on CMV, HSV, measles virus, hepatitis B virus, and HIV (1,4,11,12,15). In study on antiviral activity of IDO, Suh et al. also used siRNA directed at Indo transcripts following stimulation of human astrocyte cultures with poly-I:C and reported an ∼50% reduction of inhibition of gag protein expression in cultures infected with pseudo-typed HIV (15). The effects of IDO on influenza virus replication observed at RNA levels here appear to be consistent with their investigations. Our results, however, contrast those of Fox et al. who used 1MT to competitively inhibit IDO activity during influenza A/WSN/33 virus infection of MLE-15 cells rather than knockdown of transcripts encoding the enzyme. Moreover, these investigators assessed the effects on the virus by quantitating titers in the supernatants rather than viral RNA, which was a more direct measure of viral replication and was not affected by interference with assembly of infectious viral particles (5). It, however, remains to be determined whether these methodological differences explain the contrasting results. In vitro pharmacological inhibition of IDO in fibroblasts is needed to be performed for further validation. Based on their experimental studies in mice, Fox et al. indicated that IDO inhibition during influenza A virus infection was beneficial for the host immune response (6). Our current results suggest that the effect of such inhibition should be carefully evaluated with regard to viral infection, replication, and invasion before such regimens are introduced into humans.
We and others previously observed that proinflammatory molecules had impact on the expression of the kynurenine pathway enzymes in various cell types. IFN-γ treatment produced abundant elevation of transcripts encoding human IDO1 and increased kynurenic acid synthesis in fibroblast (2). Sellgren et al. recently reported similar results in astrocytes (14). Little is known regarding the mechanisms regulating transcription of downstream enzymes in this pathway following induction of IDO. Our previous in vitro studies reported consistently elevated levels of transcripts encoding IDO, KMO, and HAAO in influenza A/WSN/33 virus-infected neurons and glia cells (7). Using siRNA against IDO during virus infection, we detected corresponding reductions of IDO and HAAO, but not of KMO, suggesting that IDO by causing an accumulation of the tryptophan metabolite 3-hydroxyanthranilic acid potentially contribute to the transcriptional regulation of HAAO. Kynurenine can also be converted to anthranilic acid via the catalytic effects of kynureninase (Kynu). 3-hydroxykynurenine and anthranilic acid produce 3-hydroxyanthranilic acid by Kynu and an unknown enzyme, respectively. Kynu and the other unknown enzyme might play roles in the downregulation of HAAO following knockdown of IDO in virus infection investigated in our studies.
It has been reported that IDO mediated the antiviral effect of IFN-γ on suppression of hepatitis B virus replication by mainly inhibiting viral protein translation (11). In line with this, our performance of knockdown of IDO followed by influenza A/WSN/33 infection resulted in elevated virus RNA, indicating that IDO possibly interferes with viral replication. In contrast, inhibition of IDO in mouse studies detected increased type I IFNs and hence repressed murine leukemia virus replication (8). In addition, during an in vitro influenza virus infection, a decrease in viral load was observed following inhibition of IFN-λ-driven IDO upregulation (5). Our experimental data of unchanged levels of transcripts encoding IFN-β suggested that IDO is likely implicated in the process of host cell defense against influenza A virus, but not via IFN-β. Further studies are needed to determine whether IFN-α or IFN-λ or both are involved in the upregulation of IDO during influenza A/WSN/33 virus infection of fibroblast cells.
Footnotes
Acknowledgments
This work was supported by the Key Discipline Construction Program of Hunan Province, the Foundation of key programs of Hunan Educational Committee (No. 16A026), and the Stanley Medical Research Institute (No. NV-002).
Author Disclosure Statement
No competing financial interests exist.