
Abstract
Foamy viruses (FV) are retroviruses that infect several species without pathological signs, but induce substantial antibody responses in the infected host. In the case of feline FV (FFV), antibodies against Gag, Bet, and Env have been used to indicate infection; however, it is unclear whether the response to specific epitopes correlates with immunity. Here, we investigated the epitope specificity of antibodies targeting the Env protein using peptide microarrays. Sera from naturally and experimentally FFV-infected cats and pumas and from rats immunized with FFV Env expression plasmids were analyzed. An immunodominant epitope was identified in the Env leader protein (Elp), and a strong reactivity to two epitope clusters in the transmembrane (TM) subunit of Env was observed. Moreover, a short stretch of residues in the C-terminal part of the surface (SU) protein was found to be significantly associated with FFV serotype FUV-mediated neutralization. Taken together, our results add a new level of detail on the B cell epitope repertoire induced during FFV infection. Furthermore, our results provide a basis for current attempts to modify FV vectors to express and present vaccine epitopes for the directed induction of humoral immunity.
Introduction
R
Using membrane arrays spotted with overlapping peptides corresponding to the TM protein, epitope mapping was performed and epitopes were found in the fusion peptide proximal region (FPPR) and the membrane proximal external region (MPER) of the TM protein of FFV, however, a reactivity to the central part of the TM protein, which is characterized by extensive glycosylation, was not observed (19). Although these studies demonstrated that recombinant antigens were powerful in detecting FFV-specific antibodies, serological discrimination between the two main FFV serotypes, FUV and 951 (also designated as F17 serotype), is still dependent on cell-based neutralization assays exploiting the lack of substantial serum cross-neutralization between both viruses (9,27).
Alternatively, discrimination can be performed using polymerase chain reaction (PCR) on isolated viruses amplifying an about 200 bp stretch of the Env SU C-terminus, the only region found to be significantly different between both serotypes (26). Since exchange of the same domain changed the sensitivity to neutralizing antibodies specific for the corresponding serotype (27), it appears likely that this region is the main target for neutralizing antibodies. However, attempts to further define the epitope have not been undertaken so far. To extend these previous analyses, epitope mapping was performed using overlapping peptides corresponding to the entire Env and sera from animals infected with both serotypes.
Furthermore, we included sera from free-ranging pumas naturally infected with the FFV puma concolor (FFVpc) and sera from rats immunized with DNA encoding FFV Env, to identify B cell epitopes that might be typically induced after presentation of FFV Env to the immune system. Considering recent advances in using FV vectors for vaccine applications (15,16,20,23), such information could provide a rationale for the design of subunit vaccines by guiding the insertion of new or already defined pathogen-related protective linear epitopes into chimeric FV.
Materials and Methods
FFV Env expression vector
The expression vector pBC-Env was generated by cloning the complete FFV FUV Env sequence from the FFV molecular clone pCF-7 into the empty pBC vector backbone. To confirm Env expression, pBC-Env, pCF-7 with polyethylenimine (PEI) transfection reagent or PEI reagent alone (mock control) were added to 293T cells. Three days post transfection, cell lysates were analyzed by sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) and immunoblotted with a TM-specific goat antiserum (19). Equal loading was confirmed by immunostaining with a β-actin antibody (1:5,000; Sigma-Aldrich).
Animals
Wistar rats were used for immunization in accordance with a license from the Lageso (Berlin, Germany; Reference No. H0201/02). Sera from naturally infected cats were obtained from the veterinary laboratory LABOKLIN GmbH & Co.KG (Bad Kissingen, Germany), as leftovers from diagnostic testing of house and street cats. The laboratory is accredited according to DIN EN ISO/IEC 17025:2005. Puma capture and sampling were conducted with approved permits issued by the California Department of Fish and Game and the UC Davis Animal Care and Use Committee. Experimental infection of cats was performed in accordance with a license from the Regierungspräsidium Karlsruhe (application date February 27, 1998).
Sera
Sera from reference cat 8014 experimentally infected with FFV serotype FUV (1) and from three noninfected cats showing no reactivity for FFV antigens in ELISAs or in western blot analyses were used as positive and negative controls, respectively. Sera of group 1 (G1) were from 23 well-characterized naturally FFV-infected cats, previously tested positive by PCR, and ELISA-based screening for antibodies using recombinant Gag, Bet, and TM proteins or western blot analysis using FFV-infected cells as antigen (1,4,19,22,26). Group 2 (G2) comprised sera from six free-ranging North American pumas naturally infected with the FFVpc, which is highly related to FFV from domestic cats (12).
Immunization and antibody titration
Rats (group 3, G3) were immunized with the CMV immediate early promoter-driven expression plasmid pBC-Env, encoding the full-length Env protein of the FFV FUV serotype (27). The plasmid DNA was coated on 1–3 μm diameter gold particles (3 μg DNA/mg gold) and used for three successive ballistic immunizations (3 × 1 μg DNA per treatment) of Wistar rats with a Helios gene gun device (Biorad). Booster immunizations were performed at a 3-week interval and final antisera were obtained 21 days after the last injection. For the determination of FFV Env antibody titers, ELISA plates were coated with 100 ng/well of a 6xHis-tagged FFV Env antigen purified from stably transduced Crandell feline kidney (CRFK) cells by affinity chromatography. Antisera were serially diluted and bound antibodies detected with a horseradish peroxidase (HRP)-conjugated anti-rat antibody. Titers were expressed as the serum dilution exceeding the average background absorbance of the secondary antibody plus six standard deviations. All measurements were performed in triplicates.
Neutralization assay
To measure the neutralizing activity of the cat and rat sera, an improved neutralization assay using FeFAB indicator cells and FFV serotype FUV produced by transfection of 293T cells with the FFV infectious clone pCF-7 was conducted (19,27). The FeFAB indicator cells are genetically engineered FFV-permissive CRFK cells that carry the FFV long terminal repeat (LTR) promotor directing nuclear expression of β-galactosidase (β-Gal) (27). Sera were serially diluted, mixed with 200 focus-forming units of FFV FUV, incubated for 1 h at 37°C, and finally added to the cells seeded in 96-well plates. Two days later, infected FeFAB cells were fixed, stained, and scored for infection-mediated X-Gal expression from the FFV LTR β-Gal cassette using an ELISpot reader (AID, Strassberg, Germany). Neutralization titers were expressed as serum dilutions that showed more than 75% reduction of X-Gal-positive cells compared to the infected control. Since it is not dependent on syncytia formation and manual counting by microscopy but yields comparable neutralization titers, we consider this assay more sensitive and robust compared with previous assays (9,10,26). All measurements were performed in triplicates.
Epitope mapping
Epitope mapping was performed using 245 overlapping synthetic peptides (15-mers overlapping by 11) corresponding to the entire FFV Env, comprising the virion-associated N-terminal Env leader protein (Elp), SU protein, and TM protein. The peptides were synthesized and anchored on microarrays (JPT, Berlin, Germany). The application of a multiplex-format with 24 arrays per chip, each containing two replicate sub-arrays and IgG controls, allowed simultaneous screening of sera from various sources. Serum samples were incubated on the arrays using a 24 multi-well hybridization chamber (ArrayIt, Sunnyvale, CA) following the protocol recommended by JPT and analyzed in a GenePix 4000 microarray scanner (Molecular Devices, Sunnyvale, CA). Signal intensities were normalized to the input level using the spotted anti-cat and anti-rat IgG controls and epitopes were defined as central amino acids shared by overlapping peptides with signal intensities above a 3,000 relative light units (RLU) threshold.
Results
To perform epitope mappings, sera from different sources with characteristics summarized in Table 1 were analyzed using microarrays with overlapping peptides. In addition to FFV-negative cat sera and a positive control serum (cat 8014), another 23 cat sera with previously confirmed natural FFV infection (group G1) were screened. In addition, sera from free-ranging American pumas (group G2), being positive for FFVpc, which is closely related to FFV from domestic cats, were analyzed. To generate antisera against FFV Env by immunization, an expression plasmid encoding the entire Env protein of the FFV FUV serotype was constructed (Fig. 1a) and production of the Env protein was demonstrated in transiently transfected 293T cells (Fig. 1b). Similar to the expression pattern observed for the parental FFV molecular clone pCF-7, we detected the uncleaved gp130 FFV Env precursor, glycosylated gp48 TM protein, and cell lysate-associated TMCL protein in cells transfected with pBC-Env but not with the mock control (Fig. 1b). When rats were immunized three times with this vector, antisera from these animals (group G3) showed solid titers of Env-specific antibodies (0.9–2.2 × 104) 3 weeks after the last immunization as confirmed by ELISA using a recombinant FFV Env antigen (Fig. 1d). Together, these results demonstrated the success of the applied immunization regimen.
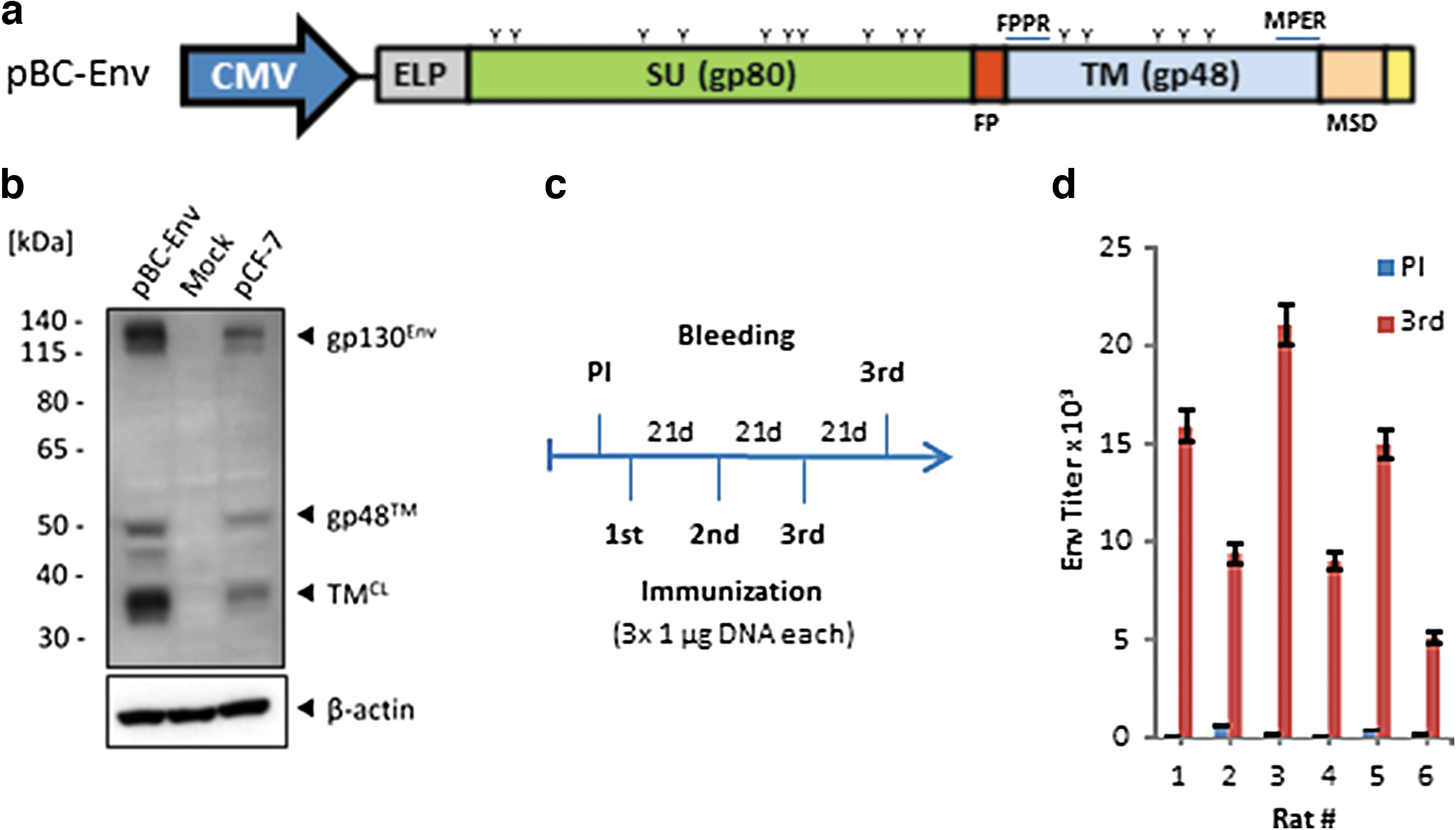
Induction of FFV Env-specific antibodies with a DNA-based immunization regimen.
Serotype derived from neutralization profile.
FFV, feline foamy virus; FFVpc, FFV puma concolor; −, negative; n.t., not tested due to limited serum availability.
Sera from all three groups were analyzed in epitope mappings using overlapping peptides corresponding to the full-length Env protein of FFV. A representative epitope map and the corresponding RLU diagram are shown in Figure 2a and b, respectively, and an overview of all samples with a colour-based intensity index is given in a heat map (Fig. 2c). The results show that all FFV-positive animals produced antibodies binding to Env with complex recognition patterns (Fig. 2c). Overall, there was stronger reactivity to the Elp/SU subunits when signal intensities in all groups were compared to that of the TM protein (p = 0.001, two-tailed Student's t-test, 95% confidence interval [CI]).
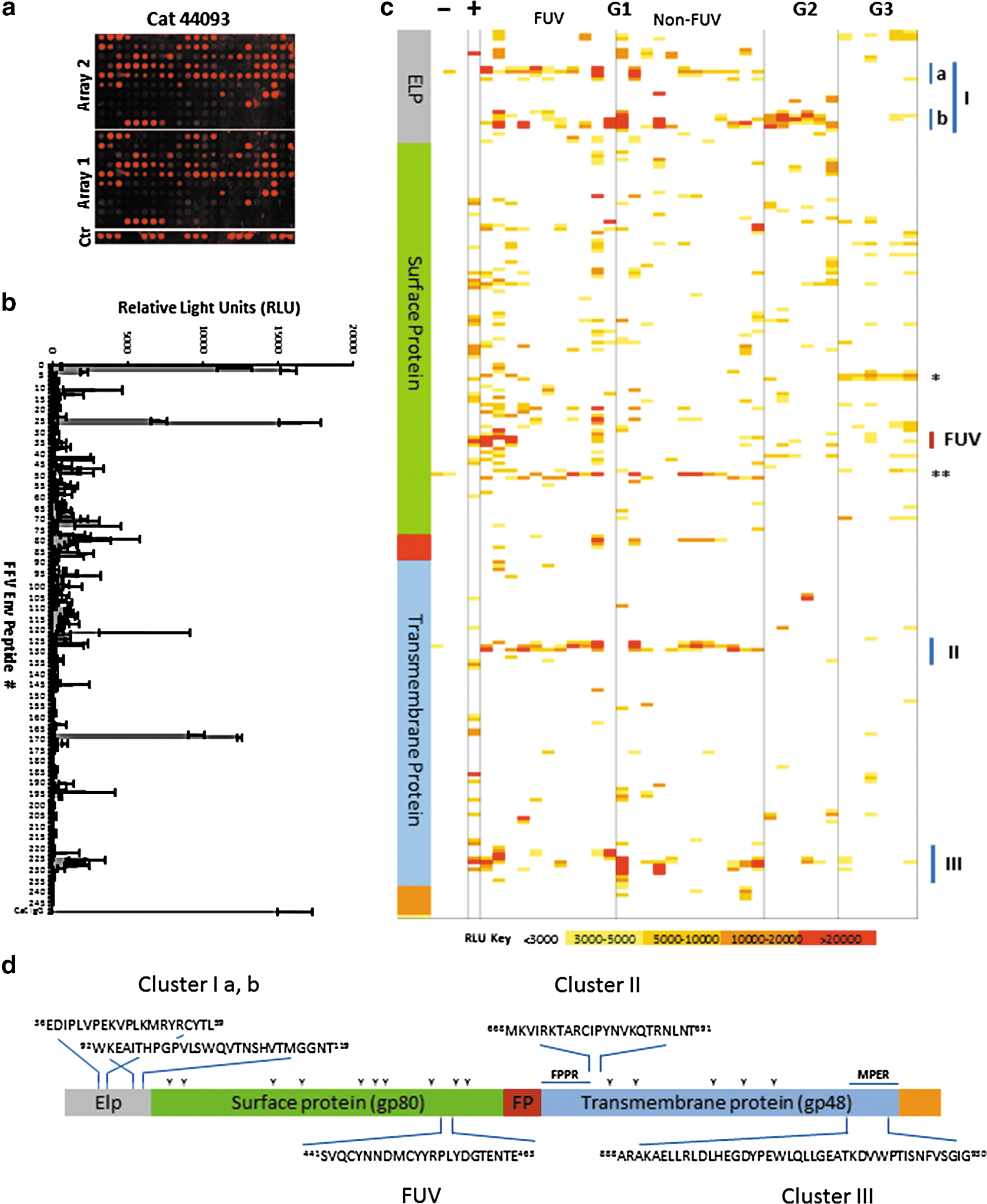
Mapping of FFV Env-specific antibody responses using microchips spotted with overlapping peptides.
The antibody response to the SU subunit was directed against different epitopes in the molecule, whereas the antibody response to the Elp and TM subunits was directed against three main epitope clusters. Cluster I, located in Elp was allocated to two sub-clusters (Ia and Ib) comprising residues 36EDIPLVPEKVPLKMRYRCYTL59 and 92WKEAITHPGPVLSW-QVTNSHVTMGGNT119 of FFV FUV Env, respectively (numbering according to Uniprot entry O56861). The epitopes are located in the N- and C-terminal from the hydrophobic membrane anchor of Elp and were recognized mainly by sera from cats and also FFVpc-infected pumas (G1 and G2, Fig. 2c, d). The cluster II was localized in the N-terminal helical region of the TM protein (aa 668MKVIRKTARCIPYNVKQTRNLNT691) and was predominantly recognized by sera from naturally infected cats (G1 in Fig. 2c, d). Epitope cluster III was localized in the MPER of the TM protein (aa 888ARAKAELLRLDLHEGDYPEWL-QLLGEATKDVWPTISNFVSGIG930). Reactivity to this domain was observed in the experimentally infected cat 8014 (+), several of the naturally infected cats (G1) and weakly in some animals from the FFVpc-infected puma cohort and the immunized rats (Fig. 2c, d). Next to these main clusters, an epitope in the SU subunit (aa 381NMPRAVLLLNK391) was almost exclusively detected by sera from rats after immunization with the FFV Env expression vector (marked *, Fig. 2c). Finally, strong responses against a single SU peptide with the sequence 485IWKKNYTLSSVLAEC499 were observed in samples from naturally infected cats (marked **, Fig. 2c). Since this epitope was also detected by sera from two of the three control animals (−) and a reactivity to a single peptide in a panel of overlapping peptides is unlikely, it was considered to be nonspecific binding.
Since neutralizing antibodies against retroviruses have been shown to react with linear and with complex conformational epitopes (11), the neutralizing activity of the cat and rat sera were analyzed using a neutralization assay based on FeFAB cells. The FFV FUV-specific 8014 control cat serum and 11 out of 23 group G1 cat sera neutralized FFV FUV at a dilution of 1:100 and higher including cat sera 10 and 12 also known to be FFV FUV-specific (Table 1) (26). In contrast, the remaining sera including the FFV 951-specific sera from cats 14 and 26 (26) showed low or no neutralization (Table 1). Based on previous findings demonstrating no substantial cross-neutralization between the two major serotypes, we thus concluded that sera with FFV FUV neutralizing activity of 1:100 and more are FUV-specific while those with weak or without neutralizing activity most probably corresponded to the 951 serotype of FFV (Fig. 2c, and Table 1). Correlating the binding reactivity to individual peptides with neutralization activity, only cat sera recognizing one cluster of three overlapping peptides in the SU domain, comprising residues 441SVQCYNNDMCYYRPL-YDGTENTE463 (marked FUV in Fig. 2c, d) were significantly associated with neutralization by the FFV FUV serum samples, but not by the non-FUV serum samples (p = 0.0475, 0.0249, and 0.0273 for peptides 111, 112, and 113, two-tailed Student's t-test, 95% CI). Considering the intensity of the complete FUV stretch (peptides 111–113 combined) the significance of the differences in binding was further improved (p = 0.0056, two-tailed Student's t-test, 95% CI) (Fig. 3). As expected, there were some neutralizing sera that did not recognize linear epitopes in this cluster region, indicating that there are also other epitopes in Env associated with neutralization, probably complex conformational epitopes.
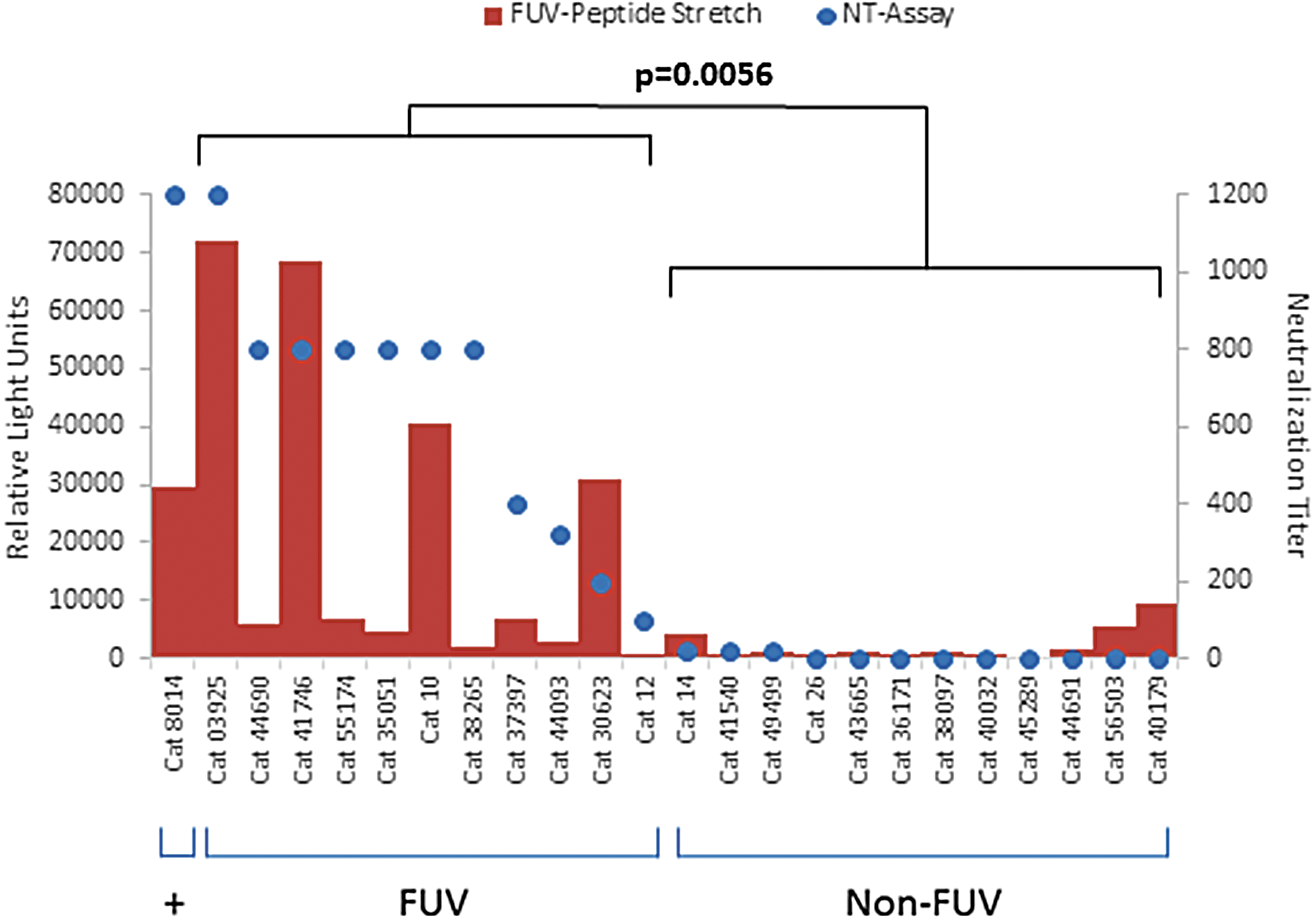
Correlation of the intensity of binding and neutralization. The sum of relative light units of the FFV-FUV peptide stretch (peptides 111, 112, 113) observed in microarrays is plotted against the neutralizing capacity of the individual cat sera of the FUV (left) and non-FUV serotypes (right) as indicated below the graph. Comparing both serotypes, strong reactivity to the three peptides was observed in most of the FUV but not the non-FUV sera (p = 0.056). +Represents reactivity of the FFV FUV infected reference cat 8014.
Discussion
This is the first comprehensive epitope mapping of antibody responses to a complete FV Env, the FFV Env protein including Elp. In general, we observed a stronger immunogenicity of FFV Env ELP/SU protein when compared to the TM subunit. This correlated with results obtained during immunization studies with the TM and SU proteins of different gammaretroviruses, where in all cases a much stronger binding and neutralizing activity was induced with the SU protein (7,8,14).
Infected cats and pumas strongly recognized epitopes in the Elp domain, mainly cluster 1b. The data confirm and extend previous findings showing a strong reactivity of sera from cats infected with FFV serotypes 951 or FUV to a recombinant Elp/SU antigen in an ELISA-based screen (4). The strong responses to Elp of sera from all cat and puma sera in this study (Fig. 2c) might explain the failure to discriminate between the two serotypes in the previous study (4). In addition, the epitope Ia is of certain interest. Although epitope Ia is part of the Elp domain that is localized in the cytoplasm of the virus-producing cell and, correspondingly, is present in the interior of FFV particles (10), it efficiently raised strong humoral immune responses. Release by disrupted virus particles or by infected apoptotic cells could be an explanation for this finding. Alternatively, antibody responses to this epitope might be triggered after uptake and proteolytic degradation of virus particles, MHC class II presentation, and activation of B cells through CD4+ T cells.
The reactivity against the TM protein described here is in line with our previous results, in which this antigen was used for detection of FFV infection (19). However, although the epitopes in the MPER, here cluster III, were nearly identical, the main epitopes detected in the FPPR were shifted more to the C-terminus. In the previous study (19) membranes spotted with 15-mer peptides overlapping by 13 amino acids corresponding only to the TM protein were used and bound antibodies were detected by peroxidase-conjugated secondary antibodies and enhanced chemiluminescence. Here, glass chips spotted with 15-mer peptides overlapping by 11 amino acids corresponding to the entire Env were used and the primary antibodies were detected by fluorescent secondary antibodies. The length of the peptides or the method of detection may be the reasons for the observed differences.
The induction of MPER-specific antibodies during FFV infection resembled immune responses described in HIV-1-infected patients and observed after immunization with the TM proteins of different gammaretroviruses, including the porcine endogenous retrovirus, the feline leukemia virus, and the Koala retrovirus (6 –8,14,18,25). In contrast to the situation described for HIV-1 and gammaretroviruses where antibodies binding to linear epitopes in the MPER were associated with neutralization (6 –8,14), we did not detect any correlation of antibodies recognizing linear peptides in the MPER of FFV and neutralization, however, we do not know whether conformational neutralizing antibodies bind to this region.
Notably, we were unable to induce FFV neutralizing antibodies with the DNA immunization regimen applied in this study and the induced epitope pattern was quite divergent from that observed during natural infections (e.g., epitope *, Fig. 2c). In immunization studies using recombinant FFV protein such differences were attributed to the lack of glycosylation of the antigen expressed in Escherichia coli (19,21), this is somewhat unexpected since the protein expressed here after DNA delivery should closely mimic the authentic antigen presented in vivo. Together, this may suggest that during FFV infection additional factors, such as incorporation of Env into virus particles, continuous replication, coordinated passage of assembled particles through the secretory pathway, and concomitant Env modifications or systemic immune activation, may further shape the induced antibody responses. In addition, our data indicate that the antiviral immunity to FFV Env is markedly influenced by the species to which this antigen is presented. This might be explained by the genetic differences of the hosts and their ability to mount distinct immune responses based on the available B cell receptor repertoire, on the expression of certain MHC class II molecules or on the responsiveness of T helper cells at the time of antigen encounter.
Analyzing the epitope reactivity and in parallel the neutralizing capacity of the sera, we were aware of the technical limitation not to detect conformational epitopes that are likely also contributing to neutralization. On the other hand, neutralizing antibodies binding to linear peptides, for example in the MPER, have been described also in the case of HIV-1 and other retroviruses (6 –8,14,18). Blocking neutralization in the case of HIV-1 using specific peptides or isolating neutralizing antibodies using immobilized epitope peptides usually failed, because a defined conformation of target molecule appears to be important for neutralization. Binding antibodies against HIV-1 often contained two subpopulations, one only binding and the other binding and neutralizing (Morozov et al., in preparation). This also explains why immunizations with HIV-1-derived peptides or recombinant proteins corresponding to the epitope failed to induce neutralizing antibodies despite the fact that the induced antibodies recognized the epitope peptide (24). It is important to underline that the gene gun expression and also the vaccination with the TM envelope protein of FFV that was purified under non-denaturing conditions (19,21) did not induce neutralizing antibodies.
Nevertheless, we identified an epitope stretch in the Env SU domain that was significantly associated with an increased sensitivity to neutralization of the FUV serotype (Fig. 2c, d). These data are supported by the finding that the corresponding sequence 441SVQCYNNDMCYYRPLYDGTENTE463 has only 45% identity with the sequence of the 951 serotype (residues without similarity are underlined). Furthermore, this region that is molecularly defined by our previous article largely overlaps with the sense primer used for serotype-specific detection of FFV-FUV but not FFV 951 by PCR on isolated viruses (26). In secondary structure predictions, the 445YNNDM449 motif forms a disordered coil and is flanked by two antiparallel beta sheets, similar to the structure of the HIV-1 V3 loop co-receptor engagement site (2,5). Furthermore, the flanking cysteines residues are predicted to be in a disulfide bonded state, potentially stabilizing the structure (data not shown). Thus, this domain might represent a loop element required for interaction with FFV's cellular receptor(s). This hypothesis is also supported by the finding that the discontinuous simian FV receptor binding domain is located in almost the same region of Env (3). Based on our data, it would be tempting to evaluate, whether a serologic discrimination between both FFV serotypes, which currently can be accessed only by neutralization assays or sequence-based methods (26), is possible by using peptides of this epitope.
Footnotes
Acknowledgments
The authors gratefully acknowledge funding by the Volkswagen Foundation, Christa-Maria Schmidt and Martina Keller for technical assistance, and Susan VandeWoude, Department of Microbiology, Immunology and Pathology, Colorado State University (Fort Collins, CO) for the puma sera.
Author Disclosure Statement
No competing financial interests exist.