
Abstract
Detection of respiratory syncytial virus (RSV) in blood, including mononuclear leukocytes and organs other than the lung, suggests that RSV disseminates outside the respiratory tract. In this study, the role of platelets in host defense against RSV was explored using an in vitro model. Platelets, also produced in the lungs, are increasingly recognized as an important part of host immune responses and may therefore play a role in modulating lung infections and clearing RSV viremia. In human peripheral blood mononuclear cells (PBMCs), platelets significantly reduced RSV infection of monocytes, monocyte activation, and interferon (IFN)α/γ production. Direct contact of platelets with PBMCs modulated the immune response when stimulated with Poly I:C (TLR3) and R848 (TLR7/8), Toll-like receptors (TLRs) involved in the recognition of RSV, and led to an enhanced IFNα/γ production. This suggested that reduction in RSV infection of monocytes in the presence of platelets could be IFN dependent; blocking IFNα receptor 2 (IFNAR2) on PBMCs indeed increased RSV infection. In addition, IFNs were not detected when PBMCs were stimulated with inactivated RSV, indicating that infection of monocytes was important for the induction of IFN responses and that the platelet-mediated reduced RSV infection was responsible for the decreased IFN levels. Furthermore, platelets could internalize RSV reducing the amount of viral particles that could infect monocytes. Our findings suggest that platelets may play a role in the clearance of RSV viremia by internalizing viral particles and by enhancing type I IFN production from PBMCs, which subsequently exert antiviral effect on host cells.
Introduction
R
While the host immune response to RSV has been studied before (1,8), no data on the putative role of platelets in RSV pathogenesis are available. Increasing evidence indicates that platelets play an important role in host defense in general and more specific in the pathogenesis of viral infections through various mechanisms (5). Recently, the lung has been identified as second site, apart from bone marrow, of platelet biogenesis from megakaryocytes (10) and may therefore have a function in modulation of lung infections. The interaction between platelets and leukocytes and the subsequent alteration in cytokine production (19) may be one of the mechanisms that also play a role in host defense against RSV. Of special importance is the role of type I interferons (IFNs), which play a crucial role in RSV host defense (6,18).
Although many viral infections are accompanied by thrombocytopenia, thrombocytosis is commonly found in children with RSV infection (9). In contrast, thrombocytopenia was found to be a risk factor for viremia and increased mortality from RSV infection in patients with hematopoietic cell transplantation (23). Interestingly, analysis of gene expression profiles from infants with LRTI of different viral etiologies showed that overexpression of platelet related genes is specific for RSV (11). All these data indicate that significant changes in platelets occur during RSV infection while we have no insight whether this also affects host defense against RSV. The aim of our present study was therefore to determine the role of platelets and platelet–leukocyte interaction in innate immune responses against RSV infection.
Materials and Methods
Isolation of peripheral blood mononuclear cells and platelets
Peripheral blood mononuclear cells (PBMCs) were isolated from buffy coats obtained from the Sanquin blood bank Nijmegen while platelets were isolated from citrate-anticoagulated venous blood collected from healthy adult volunteers as described previously (21). Briefly, PBMCs were isolated by density gradient centrifugation on Ficoll-Hypaque (Pharmacia Biotech, Uppsala, Sweden). PBMCs were washed five times with normal saline then resuspended in RPMI-1640 Dutch Modification culture medium (ICN Biomedicals, Costa Mesa, CA) and adjusted to 5 × 106 cells/mL.
For platelet isolation, 3.2% sodium-citrate-anticoagulated whole blood (Becton Dickinson, Franklin Lakes, NJ) was centrifuged for 15 min at 156 g without brake to obtain platelet-rich plasma (PRP). PRP was further centrifuged at 330 g for 15 min to obtain platelet pellets and resuspended in HEPES tyrode buffer.
Culture of RSV (GFP-RSV)
Green fluorescent protein (GFP) expressing RSV-A2 (rgRSV30) was kindly provided by Dr. M.E. Peeples (7) and was cultured on HeLa cells. Subsequently the virus was purified by ultracentrifugation over 30% sucrose cushion as described previously (22). To quantify the number of infectious virus particles, HeLa cells (80–90% confluent) were infected with fivefold viral dilutions for 20–22 h. The virus titer was determined by counting wells with ≥10 and ≤100 infected cells/view (Olympus CKX41 microscope) where each data point was the average of an experimental duplicate (22). Inactivated rgRSV30 was produced by incubating the virus with 0.025% beta-propiolactone (BPL) (Acros Organics) for 16 h at 4°C under continuous gentle shaking. Next, the stock was incubated for 4 h at 37°C to hydrolyze the BPL and stored at −80°C until further use.
RSV infection and stimulation of PBMCs
PBMCs at a concentration of 5 × 106 cells/mL were dispensed into a round-bottomed 96-well plate (Thermo scientific, Roskilde, Denmark) and infected with live or BPL inactivated rgRSV30 at multiplicity of infection (MOI) of 10 in the presence and absence of platelets. For the transwell system assay, PBMCs were dispensed into the lower compartment of a flat-bottomed 24-well plate with a 0.4 μm filter membrane (Greiner Bio-one, Frickenhausen, Germany) and stimulated with 100 μg/mL Poly I:C (InvivoGen, Toulouse, France) or 5 μg/mL R 848 (InvivoGen) in the presence of platelets or physically separated platelets dispensed into the upper compartment of the transwell. PBMC to platelet ratio used was 1:150. Negative controls included stimulated PBMCs with Roswell Park Memorial Institute medium (RPMI) in presence or absence of platelets. IFNα receptors on PBMCs were blocked using anti-IFNAR2 antibodies (PBL assay science, Piscataway, NJ) or IgG2A isotype control (eBioscience, San Diego, CA) for 1 h at 37°C before adding platelets or RSV. Cells were incubated for 48 h at 37°C and 5% CO2, and supernatant was collected after centrifugation and stored at −20°C.
Flow cytometry
To determine the percentage of monocytes infected with RSV and expression of CD16 on monocytes, PBMCs were cultured with RSV for 48 h in presence and absence of platelets as described above. Cells were washed with cold PBS and then labeled for monocyte markers with anti-CD14 (anti-CD14 ECD; Beckman Coulter, Fullerton, CA) and anti-CD16 (anti-CD16 PE; eBioscience) antibodies for 30 min, washed again, and fixated with paraformaldehyde (1%). Cells were analyzed using the FC500 flow cytometer with CXP software (Beckman Coulter). Mean fluorescent intensity (MFI) or percentage of RSV infection and expression of CD16 on monocytes were determined by gating on the population of cells double positive for CD14 and RSV and CD14 and CD16, respectively.
To study whether platelets internalize RSV, washed platelets at a concentration of 5 × 105 cells/mL were incubated with RSV-A2 that does not express GFP at MOI of 5 for 2 h at 37°C, 5% CO2 on a shaker.
After incubation cells were washed twice using bovine serum albumin (BSA) buffer and then stained for CD61 (anti-CD61; Beckman Coulter) as platelet identification marker and for surface expression of platelet alpha granule protein P-selectin (anti-CD62p; BioLegend, San Diego, CA) as platelet activation marker for 30 min at 4°C. Thereafter cells were washed and resuspended in fixation/permeabilization solution for 20 min followed by perm/wash buffer (both obtained from BD Biosciences, Breda, The Netherlands) for 10 min at 4°C. Cells were then stained for intracellular expression of RSV nucleoprotein (NP) using FITC labeled anti-NP antibodies (Abcam, San-Francisco) diluted in perm/wash buffer for 30 min at 4°C. After a wash step cells were analyzed on a BD LSR II flow cytometry machine. Platelets were identified based on their forward and side scatter characteristics, from which populations double positive for platelet marker anti-NP and CD62p were identified.
Cytokine determination
Cytokines in collected supernatants were quantified using commercially available enzyme-linked immunosorbent assay (ELISA) for IFN gamma (IFNγ) (obtained from Sanquin, Amsterdam, Netherlands) and IFN alpha (IFNα) (obtained from R&D, Abingdon, United Kingdom) according to the manufacturer's instructions.
Statistical analysis
Statistical analysis was done using GraphPad prism 5 software. Data were analyzed using Kruskal–Wallis and Mann–Whitney tests or student t-test (data on expression of RSV NP in platelets). Data are presented as mean of independent samples unless stated otherwise. Flow cytometry analysis was done using Beckman Coulter Kaluza software, version 1.2.
Results
We first investigated whether platelets modulated RSV infection of monocytes. PBMCs were infected with RSV at MOI of 10 in the presence and absence of platelets. Addition of platelets significantly decreased RSV infection of monocytes after 48 h of incubation (Fig. 1A and Supplementary Fig. S1; Supplementary Data are available online at
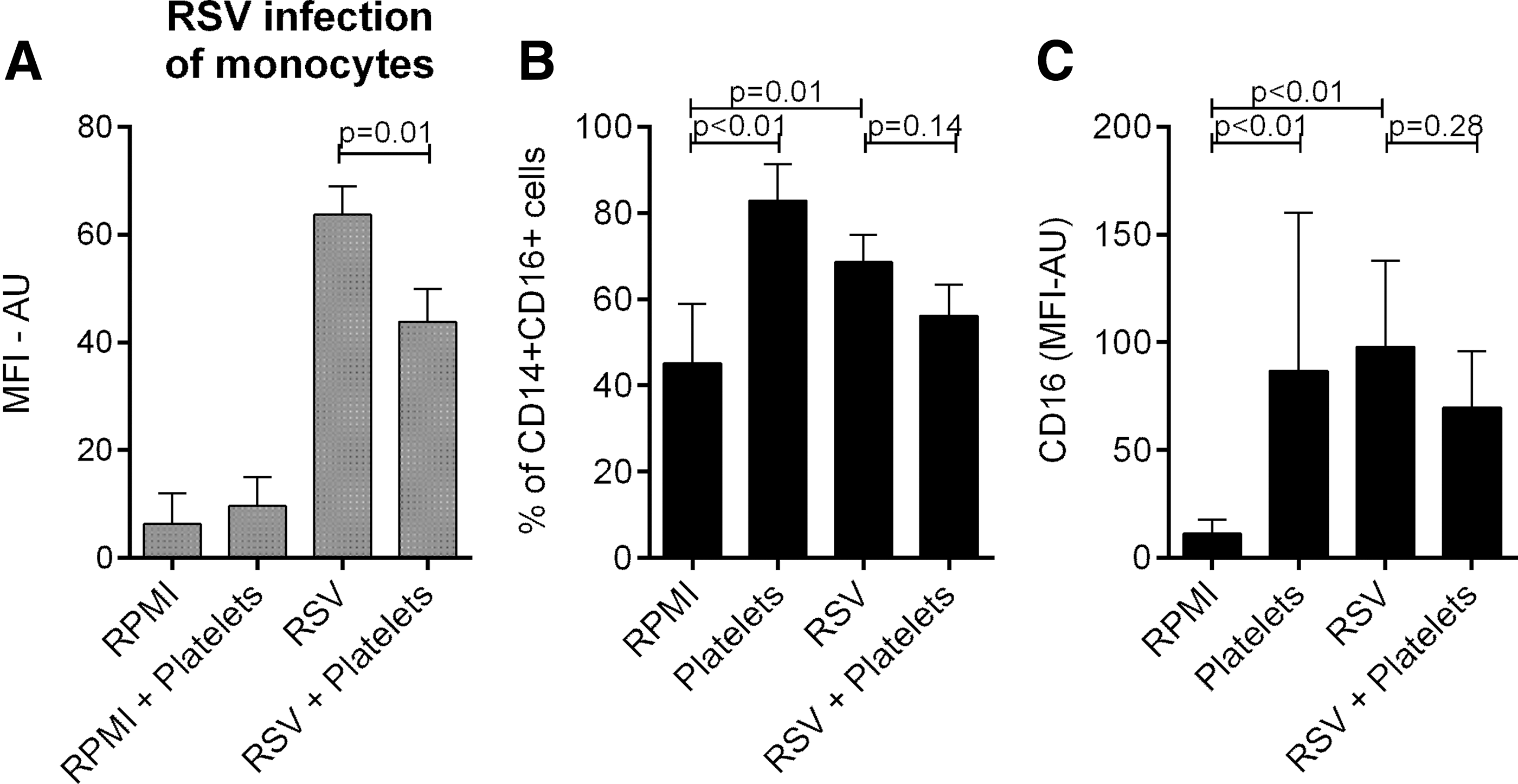
RSV infection of monocytes and expression of CD16 on monocytes (CD14+ cells) after stimulation of PBMCs with RSV.
We next explored whether the reduced monocyte infection and activation rate resulted from increased production of antiviral cytokines IFNα and IFNγ. Concentrations of these cytokines in the culture supernatant were lower when PBMCs were stimulated with RSV in the presence of platelets. In addition, BPL inactivated RSV did not induce production of IFNα/γ from PBMCs (Fig. 2). To avoid confounding by monocyte infection rate, we stimulated PBMCs with the synthetic TLR3 and TLR7/8 ligands poly I:C and R848. RSV is a known ligand for these Toll-like receptors (TLRs) (8). IFNα and IFNγ production by PBMCs upon stimulation with these TLR ligands was increased in the presence of platelets (Fig. 3); however, differences were not significant for IFNγ production after stimulation with R848. These effects could be reversed by physical separation of PBMCs and platelets by a transwell system (Fig. 3). Blocking IFNAR2 on PBMCs using anti-IFNAR2, but not IgG2A isotope control, increased RSV infection of monocytes and monocyte activation (CD16 expression) (Fig. 4 and Supplementary Figs. S1 and S2).
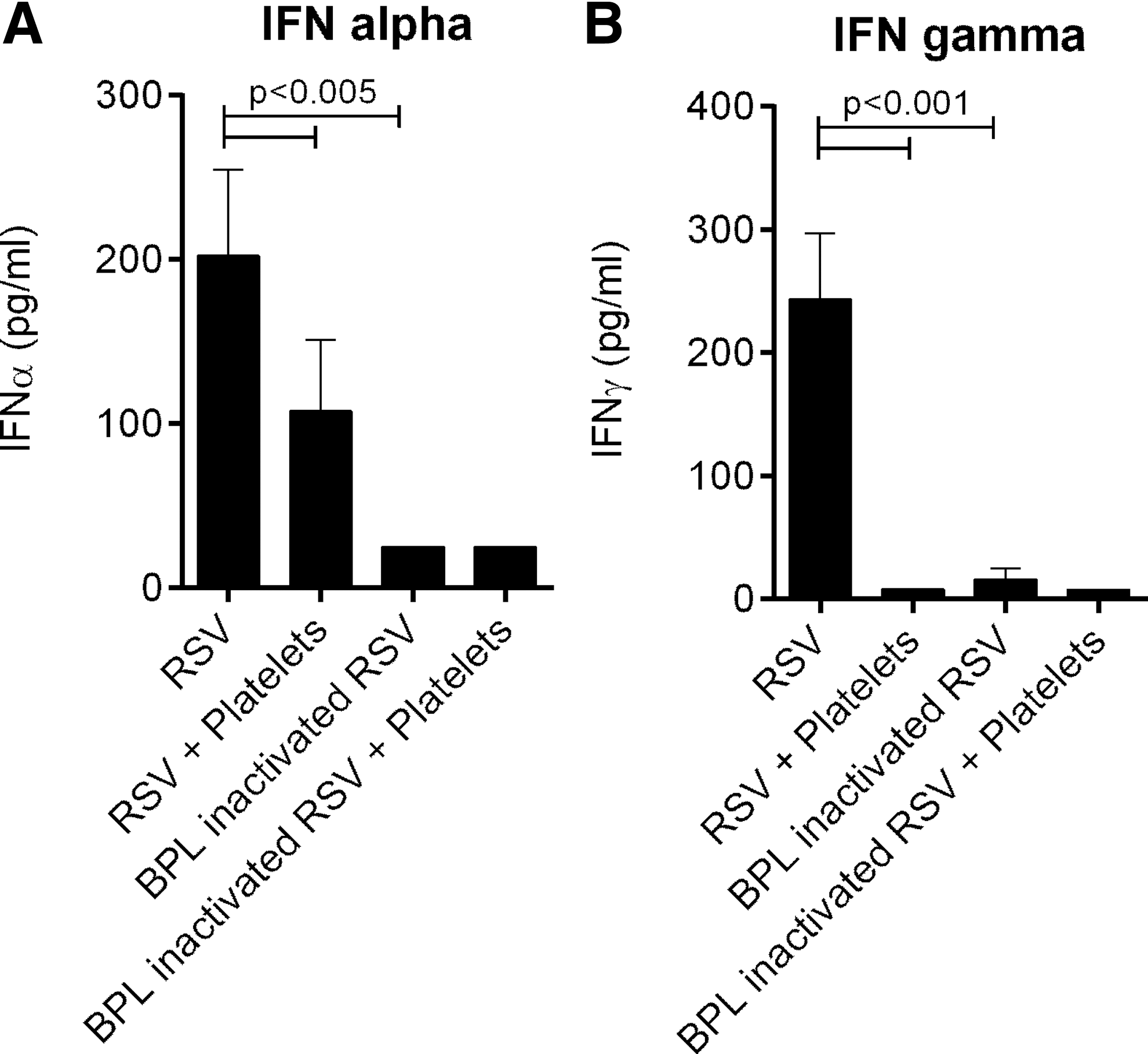
IFN production after stimulation of PBMCs with RSV.
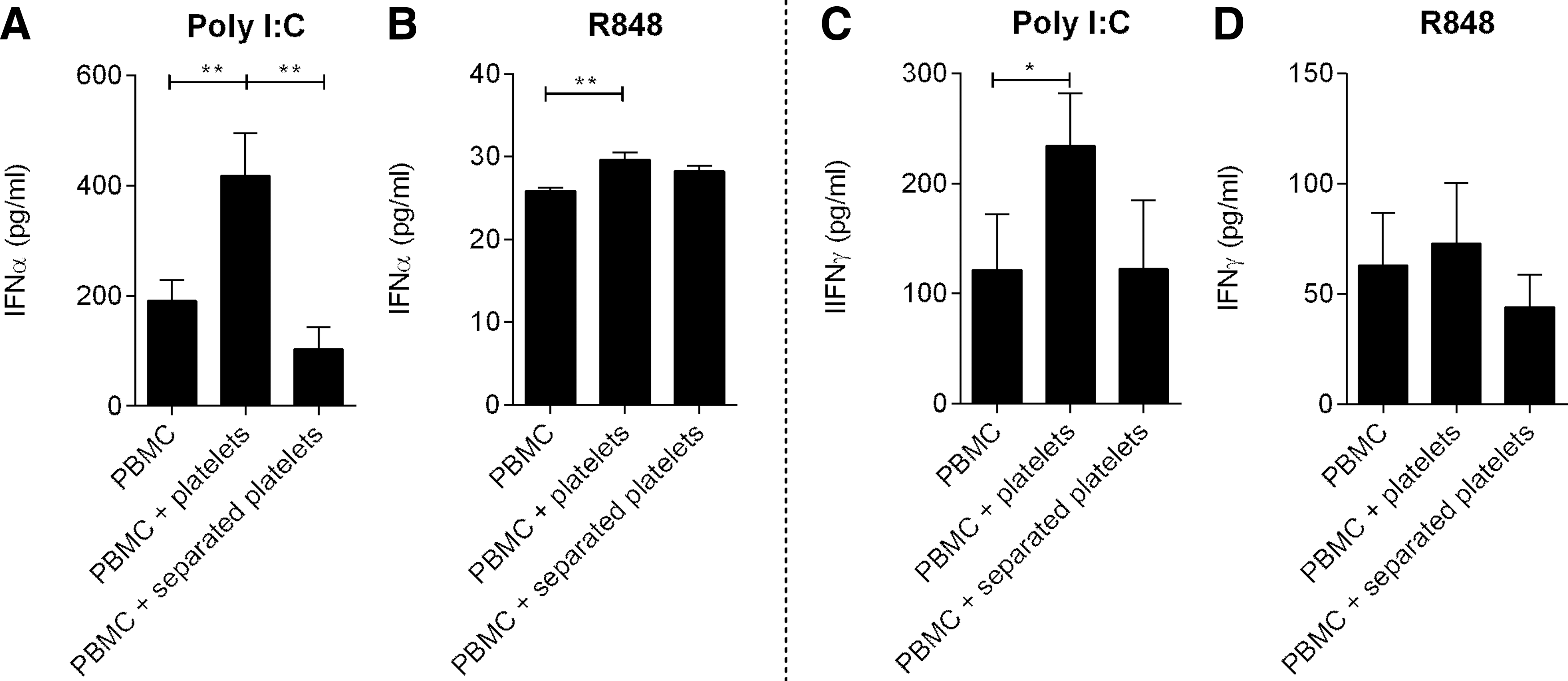
IFN production after stimulation of PBMCs with Poly I:C and R848. IFNα and IFNγ produced after stimulation of PBMCs using Poly I:C (TLR3 ligand)

RSV infection of monocytes and expression of CD16 on monocytes (CD14+ cells) after blocking IFNAR2 on PBMCs.
To study whether the reduced monocyte infection with RSV resulted also from internalization of RSV by platelets, washed platelets were incubated with RSV for 2 h. Platelets incubated with RSV had significantly higher MFI of intracellular RSV NP and higher percentages of platelets expressing RSV NP (Fig. 5A and Supplementary Fig. S3). Surface P-selectin expression was also increased in RSV exposed platelets (Fig. 5B and Supplementary Fig. S3). These results indicate that platelets can internalize RSV, a process that leads to platelet activation and degranulation.
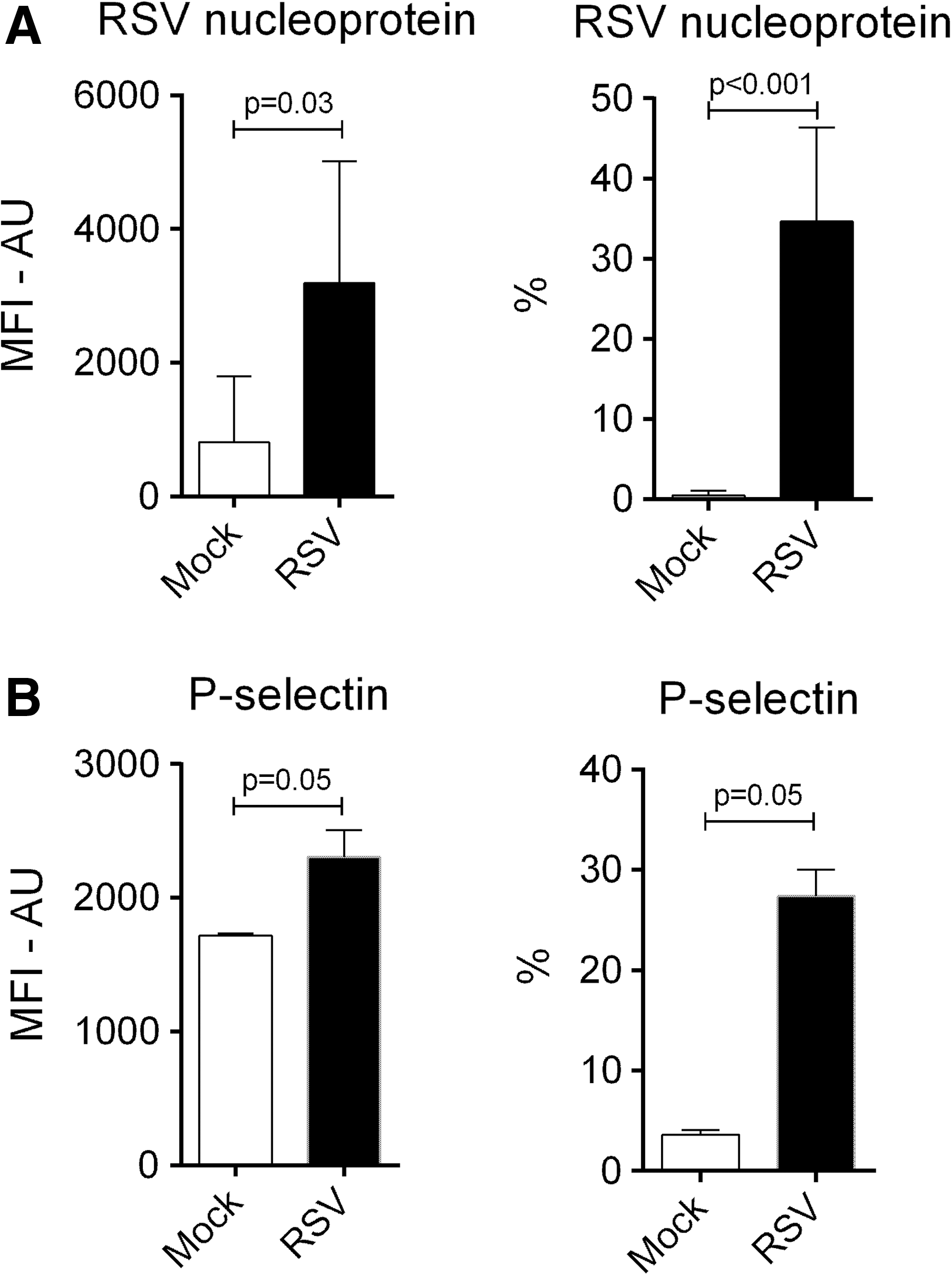
Expression of RSV NP and P-selectin on platelets. MFI-AU of and percentages of platelets expressing RSV NP
Discussion
Increasing evidence supports the idea that platelets play a role in host defense against infections. Our data show that platelets can reduce RSV infection of monocytes and monocyte activation by enhancing IFN production from leukocytes and by internalizing viral particles.
IFNs induce antiviral responses when pattern recognition receptors in the cytoplasm of host cell detect viral associated motifs and are dependent on the capacity of the virus to infect a host cell (17). Our results show that platelets can enhance antiviral responses by enhancing production of IFNs from PBMCs following stimulation with viral antigen. This IFN dependent antiviral effect was exhibited by reduced RSV infection of monocytes in the presence of platelets and confirmed by increased infection after blocking IFNα receptor2 on PBMCs. RSV infection of monocytes is a well-known inducer of IFNα/γ (6,17). This observation explains the simultaneous reduction of IFN production with reduced RSV infection of monocytes observed in our study. In addition, inactivated RSV could not induce production of IFNα/γ from PBMCs. Increased IFN production from PBMCs following stimulation of TLR3 and TLR7/8 in the presence of platelets provides further evidence of the link between RSV infection rate and IFN production. The transwell system portrays the importance of direct contact between platelets and leukocytes in these effects. Platelet effects mediated by direct platelet–leukocyte interaction have also been described earlier (15,21).
Platelets can also have inherent ability to directly recognize and clear pathogens and release immune mediators (14). Platelet mediated reduction of monocyte infection with RSV could also be explained by the internalization of RSV by platelets (2). Put together, our findings indicate that platelets enhance type I IFN production from PBMCs exerting antiviral effect on host cells; and second, platelets internalize viral particles and, hence, reduce the number of particles that can infect monocytes, a process that may play a role in clearing RSV viremia.
RSV primarily infects the lung and it is possible that platelets also interact with alveolar macrophages. A limitation of our study is that we did not use alveolar macrophages. However, PBMCs have been proposed as a robust and reproducible in vitro model to study both innate and adaptive responses to RSV (22). Furthermore, platelet mediated reduction of monocyte activation and IFNα during RSV infection as observed in our study may be an important factor in preventing lung inflammation in vivo.
Data presented are based on an in vitro model using a recombinant RSV strain and isolated PBMCs. The protective effect of platelets to alveolar macrophages and epithelial cells, which are the main targets for RSV in vivo, poses need for further studies.
Conclusion
Collectively, our current study shows that platelets play an important role in host innate defenses against RSV by reducing RSV infection of monocytes and monocyte activation in a type I IFN dependent manner. Platelets can also internalize viral particles and may therefore be important in preventing RSV dissemination and disease exacerbation.
Footnotes
Acknowledgments
The authors thank Jop Jans and Wouter van der Heijden for laboratory assistance. This project was funded by the Netherlands Fellowship Programme.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.