
Abstract
The attenuated C-strain vaccine against classical swine fever virus (CSFV) is one of the safest and most effective attenuated vaccines. However, little is known of the host immune response after vaccination with the C-strain vaccine. Blood samples from vaccinated pigs were collected to evaluate the number of immune cells, the level of specific CSFV antibody, and related cytokines induced by the vaccination of C-strain vaccine. The C-strain nucleic acid was gradually removed and specific antibody to vaccine kept increasing; the amount of the lymphocyte, Tc cell, and Th cell increased; some inflammatory cytokines such as interleukin (IL)-1 and tumor necrosis factor-α mainly showed downregulated trends, but IL-6 and IL-8 were upregulated greatly; IL-2, IL-4, IL-5, IL-12p40, IL-13, interferon (IFN)-I, and Toll-like receptors (TLRs) kept high expression level after 28 days postvaccination (dpv); IFN-γ was upregulated slightly at 5 and 9 dpv, respectively. These results suggest that the C-strain vaccine induces a Th2 cell response to produce the specific antibody. The vaccine virus replicates at very low level. C-strain vaccine burden has close relationship with the expression of TLRs. The overexpression of TLRs initiates the innate immune system to clear up the vaccine. Meanwhile, ILs expressed by immune system induce the differentiation of B cells and produce specific antibody.
Introduction
T
Materials and Methods
Animals and vaccine
Thirteen healthy 30-day-old Large White/Landrace cross male piglets were purchased from a local commercial source. Before vaccination, the animals were tested negative for CSFV antibody and bovine viral diarrhea virus antibody by means of the neutralization test. The animal study proposal was approved by the Animal Care and Use Committee of the China Institute of Veterinary Drug Control. All experimental procedures involving the piglets were performed in accordance with the Regulations for the Administration of Affairs Concerning Experimental Animals approved by the State Council of People's Republic of China. The C-strain vaccine, whose trade name is Classical Swine Fever Live Vaccine (Cell Line Origin), was kindly provided by Winsun Pharm. For vaccination with the piglets, the vaccine was reconstituted in vaccine diluent as described by the manufacturer.
Vaccination with the C-strain vaccine
The piglets were randomly divided into two groups: the vaccinated group and control group. The immunized group, including eight piglets, was vaccinated with the C-strain vaccine (one dose each). The control group, including five piglets, was immunized with the same amount of sterile saline.
Blood sampling
Blood was collected from precaval vein at 0, 1, 2, 5, 7, 9, 12, 16, 20, 23, 28, 30, 35, and 43 days postvaccination (dpv) from each piglet. Four milliliters was injected into an EDTA-K2 anticoagulant tube to separate leukomonocytes and detect C-strain nucleic acid load, and other 2 mL was used to separate serum to detect CSFV antibody.
Detection of CSFV nucleic acid load and antibody
Total RNA was extracted from 200 μL EDTA-blood using a MeDiPro Tissue RNA Auto Kit as described by the manufacturer (Formosa Biomedical Technology Corp). The level of CSFV nucleic acid load was analyzed by real-time reverse transcription-polymerase chain reaction (RT-PCR) using porcine β-actin as an internal control (13,17). Using the first detectable Ct value as the basic value, relative quantification data of CSFV nucleic acid load were analyzed by the 2−ΔΔCT method (14). Serum was examined by a CSFV Antibody Kit as described by the manufacturer (IDEXX Europe B.V.).
Detection of routine blood cells, immune cells, and related cytokines
EDTA-blood (100 μL) was counted by a semiautomatic blood cell counter to quantify the total number of leukocytes, lymphocytes, neutrophil granulocytes, erythrocytes, and platelets. Lymphocytes isolated from 100 μL EDTA-blood were resuspended in fluorescence wash buffer and stained with FITC-conjugated anti-CD3, CD4, and CD8 monoclonal antibodies (mAbs) (Southern Biotechnology Association, USA) for 30 min at 4°C in the dark. Samples were washed with fluorescence wash buffer and analyzed by a flow cytometer. CD3+ CD4+ CD8− T cells were defined as Th cells, CD3+ CD4− CD8+ T cells were defined as Tc cells, CD3− CD4− CD8+ cells were defined as NK cells, CD3+ CD4+ CD8+ cells were defined as activated memory T cells, and CD3+ CD4− CD8− cells were defined as γδT cells (17).
For exploring the cytokine gene expression levels after vaccination, total RNA was extracted from peripheral blood mononuclear cells (PBMCs) using a MeDiPro Tissue RNA Auto Kit as described by the manufacturer (Formosa Biomedical Technology Corp). cDNA was synthesized using random primers (Invitrogen). First-strand cDNA was amplified by real-time PCR using a SYBR® Premix Ex Taq™ (Perfect Real Time) Kit (Takara Biotechnology, Co., Ltd.) with the housekeeping gene porcine β-actin as an internal control. Gene expression at 0 dpv was used as baseline for comparison. All of the primer pairs used are previously described (3).
Statistical analysis
For each independent gene expression experiment, a one-way analysis of variance (ANOVA) was used to compare the experimental vaccinated group to the expression level at 0 dpv. ANOVA analyses were performed using GraphPad Prism 5 (Prism 5 for Windows, Version 5.01; GraphPad Software, Inc. La Jolla). Statistical significance was defined as p < 0.05.
Results
The levels of CSFV nucleic acid load and antibody in the vaccinated group
The CSFV nucleic acid load was first detected at 7 dpv, increased to a peak at 23 dpv, and then decreased. Minimal virus RNA was detectable in the peripheral blood cells at the end of the experiment (43 dpv). The CSFV antibody level increased from 5 dpv and kept increasing to the end of experiment. Seroconversion occurred at 12 dpv. All control animals were negative both in CSFV nucleic acid and CSFV antibody during the experiment (Fig. 1).

The levels of the C-strain nucleic acid load and specific CSFV antibodies induced by C-strain vaccine in piglets. Nucleic acid of C-strain was amplified by real-time RT-PCR using the porcine β-actin gene as an internal control. Nucleic acid was first detected at 7 dpv. Specific CSFV antibodies induced by C-strain vaccine or sterile saline in serum of pigs were detected by a commercial ELISA Kit. Data are expressed as the mean for each group of pigs, and error bars represent the SEM. CSFV, classical swine fever virus; dpv, days postvaccination; RT-PCR, reverse transcription-polymerase chain reaction.
Routine blood cell and T lymphocyte subset analysis
There were no significant differences between the vaccinated group and control group concerning the number of neutrophil granulocytes, erythrocytes, platelet, γδT cells, and activated memory T cells. The lymphocyte levels began to increase at 5 dpv, peaked at 12 dpv, and then decreased to the control level. Tc cells transiently increased in the vaccinated group at 12 dpv. Th cells increased from 5 dpv, peaked at 9 dpv, which is 2.4-fold greater than the control group, and then decreased to control levels (Fig. 2).
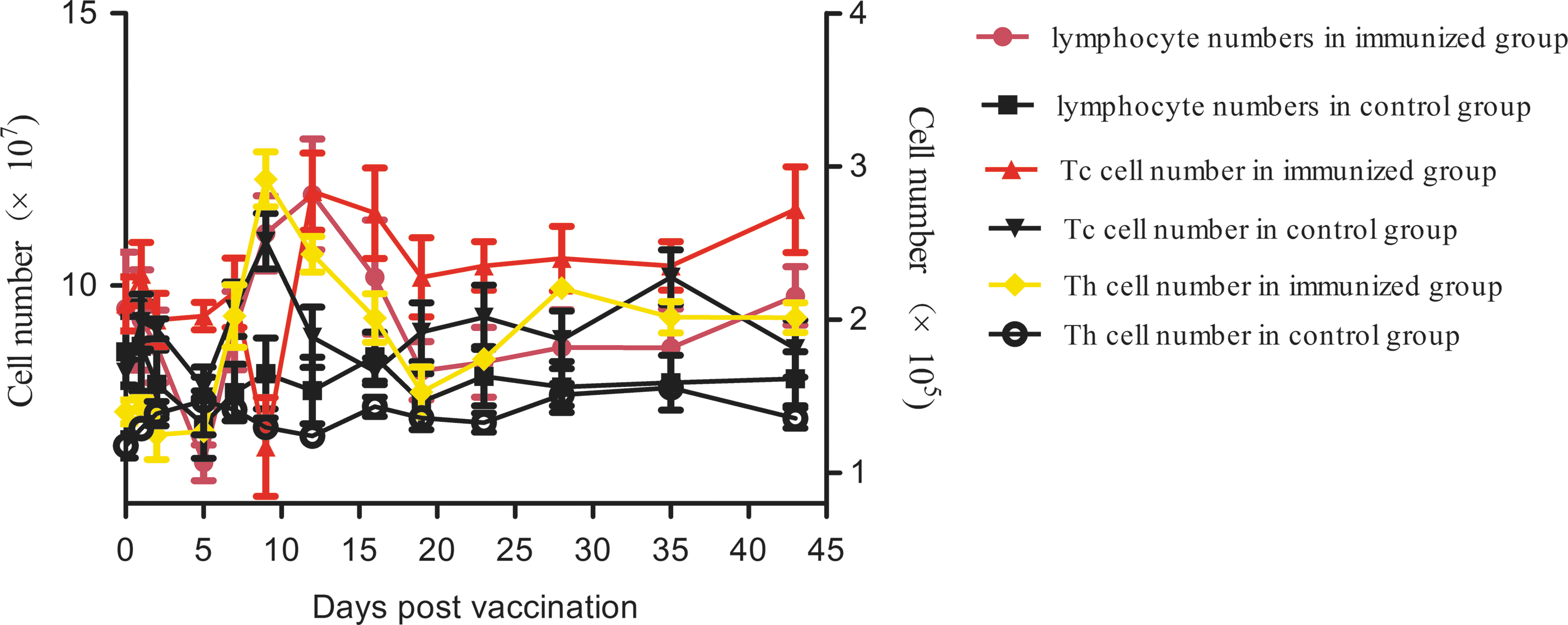
The changes of lymphocyte, Tc cell, and Th cell numbers after immunization with the C-strain vaccine. Lymphocyte numbers were counted by a semiautomatic blood cell counter. Lymphocytes stained with FITC-conjugated anti-CD3, CD4, and CD8 mAbs were counted by flow cytometry. CD3+ CD4− CD8+ T cells were defined as Tc cells and CD3+ CD4+ CD8+ T cells were defined as Th cells. mAbs, monoclonal antibodies.
The gene expression level of cytokines and Toll-like receptors in control group
The gene expression level of cytokines and 10 Toll-like receptors (TLRs) kept stable in control group. There were no obvious changes during the experiment.
The gene expression level of cytokines and TLRs in vaccinated group
The gene expression levels of interleukins
The gene expression levels of the inflammatory cytokines interleukin (IL)-1α and IL-1β irregularly changed before 20 dpv and then decreased from 20 to 43 dpv. IL-6 level was upregulated from 5 dpv, increasing 1,000-fold at 16 dpv, and then decreased to control levels. Chemokine IL-8 levels were transiently upregulated at 5, 9, and 16 dpv, corresponding to 4,570-, 30-, and 20-fold increase, respectively (Fig. 3a). Five cytokines (IL-2, IL-4, IL-5, IL-12p40, and IL-13) displayed nearly the same trends in that all remained upregulated from 28 dpv after a transient upregulation at 1, 5, 9, and 20 dpv, respectively (Fig. 3b).

The gene expression levels of TNF-α and IFNs
TNF-α was downregulated 4.3- and 98-fold at 7 and 23 dpv, but was upregulated 14-fold at the end of the experiment. The gene expression levels of IFN-I (IFN-α and IFN-β) irregularly changed before 20 dpv. However, at 20 dpv, both IFN-α and IFN-β were transiently upregulated 13- and 14-fold, respectively, and kept increasing from 28 dpv. IFN-II (IFN-γ) was upregulated 5.5- and 5.6-fold at 5 and 9 dpv and downregulated thereafter (Fig. 4a).

The gene expression levels of TLRs
The gene expression levels of 10 TLRs were examined, and all of them have the same trends, that is, upregulated at 1, 9, and 20 dpv, respectively, and kept upregulated from 28 dpv to the end of the experiment (Fig. 4b).
Discussion
C-strain virus replicates in lymphoid tissues, especially in tonsil, and only trace of virus nucleic acid could be detected in blood (18). After 6 to 7 weeks postvaccination, vaccine virus is removed completely (31). But after infection with the virulent CSFV, virus was detected in many tissues and organs from 1 day postinfection (dpi), and virus load increased with the infection continued (13). In this study, in accordance with a previous study (31), vaccine nucleic acid was detectable from 7 dpv and reached to a peak at 23 dpv, which was only 101.28 ± 0.054 fold to 7 dpv, and then decreased. This result showed that C-strain vaccine replicated at a very low level in vaccinated pigs and it showed a trend from accumulation gradually to clearance gradually.
Lymphocytopenia is one of the most typical symptoms of CSFV infection. After infection with virulent CSFV, the number of the platelets, NK cells, Th cells, Tc cells, γδT cells, and CD3− CD4− CD8− and CD3+ CD4+ CD8+ lymphocytes are significantly decreased (32). In this study, the number of lymphocytes increased from 5 to 20 dpv. Through analysis of the lymphocyte subsets, we found that the number of Tc cells transiently increased at 12 dpv, but the number of Th cells increased obviously from 5 to 16 dpv. These results indicate that the C-strain vaccine stimulates the proliferation of Th cells. Th2 cells mainly secrete IL-4, IL-5, IL-6, IL-8, IL-10, and IL-13 and stimulate B cell to multiply and induce B cell antibody class switching, as well as improve the neutralizing antibody production. In this study, the number of Th cells increased from 5 to 20 dpv, meanwhile the levels of IL-6, IL-8, and CSFV antibody increased. These results indicate that cells are activated and Th0 cells differentiate into Th2 cells in this phase.
Inflammatory cytokines, such as TNF-α, IL-1, and IL-6, regulate the immune system and cause specific inflammatory reaction, which is the important cause of anti-infection and tissue damage. Many studies have proven that either middle or highly virulent CSFV can cause lymphocyte apoptosis, which leads to lymphocytopenia and thrombocytopenia (3,19,20). TNF-α released from CSFV-infected macrophages may induce apoptosis in both infected and uninfected bystander cells (5). IL-1 is another important inflammatory cytokine against infection, which causes fever, hyperalgesia, vasodilation, and hypotension (6). In this study, vaccination with the C-strain vaccine did not increase the expression levels of TNF-α and IL-1, thereby avoiding the damage caused by inflammation. IL-6 is a pleiotropic pro-inflammatory cytokine (10). Both in vivo and in vitro studies have demonstrated that IL-6 is overexpressed in the infected cells and it is related to the damages of immune cells and tissues (2,16,19). However, in addition to its function in inflammation, IL-6 is also involved in a wide variety of biological functions such as essential roles in the final differentiation of B cells into Ig-secreting cells and in induction of Th2 cell response (22). In this study, IL-6 levels began to increase at 7 dpv, peaked at 16 dpv with a 1,000-fold upregulation, and decreased thereafter. Considering the changes in antibody levels, the period of 7–30 dpv was the critical period for antibody secretion and T cell activation. Thus, our results indicated that the overexpression of IL-6 did not induce obvious inflammatory damages in the vaccinated pigs; however, it played an important role in immune regulation after vaccination with the C-strain vaccine.
The innate immune system is the first and fast activation of the innate immune response against pathogens; the major objective of the innate response is to induce pro-inflammatory cytokines and IFNs and to set up a bridge of communication subsequent to the onset of adaptive immunity (11). Not only virus has tactics to evade host immune system but also the host initiates an immunosuppressive program that actively suppresses antiviral T cell responses and facilitates persistent infection. The expression of suppressive factors is tightly linked to viral burden, suggesting the presence of an immunologic sensory system that continually measures the magnitude and duration of viral replication and then dynamically modulates the balance between antiviral immunity and immune exhaustion (30). In our study, vaccine nucleic acid was detectable from 7 dpv, reached to a peak at 23 dpv, and then decreased. Before 23 dpv, the expression level of IFNs, most of ILs, and TLRs did not increase persistently. This result suggests that at early phase after vaccination with C-strain vaccine (before 23 dpv), virus replicates so slow that host immune system does not sense the presence of virus, so host initiates an immunosuppressive program that actively suppresses antiviral T cell responses. When the virus accumulates to a certain extent at late phase of vaccination (after 23 dpv), the immunosuppressive program is stopped, host initiates the immune response.
TLRs, which are expressed on the surface of immune cells, initiate and regulate innate immunity and induce adaptive immunity (12). TLR3, 7, 8, and 9, the most important pattern recognition receptors for viral nucleic acid, activate special signal transduction pathways to induce the production of IFN-I and IFN-II (15,21). Highly virulent CSFV downregulates the expression levels of TLRs in macrophages (13). In this study, the expression levels of 10 TLRs displayed the same trends, which were in accordance with the expression levels of IFN-I. Our results indicate that when it accumulates to a certain extent, C-strain virus could upregulate the expression levels of TLRs and then induce the upregulation of IFN-I and some cytokines to clear the virus.
IFN-I signals possess antiviral and immune stimulatory potential required for the clearance of infection, but when virus cannot be controlled, these sustained IFN-I signals induce overexpression of inflammatory cytokines and chemokines, which may do harm to immune cells and immune organs, and lead to immunosuppression and persistent infection (27,30). Several studies showed that IFN-α is detectable in blood 2 dpi with highly virulent CSFV, with maximum levels at 3–5 dpi. Meanwhile, the onset of severe lymphopenia is concomitant with the IFN-α response, and all infected animals display depleted B and T lymphocytes. These facts indicate that the apoptosis of lymphocytes is closely related to the untimely overexpression of IFN-α (22). In this study, there was no obvious IFN-I response before 20 dpv, indicating that IFN-I might have no effect on antiviral immunity. The vaccine virus load continually increased during that period. However, after 28 dpv, the expression of IFN-I and other cytokines increased and the virus nucleic acid load decreased. These results suggested that IFN-I increased at a proper time point not only to protect immune cells and organs from damaging but also to avoid the vaccine virus being cleared too early. So the C-strain virus replicated slowly at the early stage after vaccination and gently stimulated the immune system, therefore avoiding damage to immune cells and immune organs caused by the overexpression of IFN-I.
Type II IFN (IFN-γ) is released by antigen-active T cells, playing an important role in immune regulation (1). Many studies have demonstrated that unimmunized animals cannot produce IFN-γ after infection with virulent CSFV (26). However, for the immunized animals, infection motivates a rapid and high-level IFN-γ response, which provides protection to immunized animals. Abundant IFN-γ contributes to CSFV control, at least until the emergence of neutralizing antibody (26). In this study, IFN-γ levels were transiently upregulated twice at 5 and 9 dpv, respectively. Similar results were described elsewhere (23). These low-level transient upregulations illustrated that T cells transiently responded to vaccination and entered a sensitization status. There is a close temporal correlation between T cell IFN-γ responses and rapid protection induced by C-strain vaccination (8). Once they were stimulated by virulent virus, T cells could release plenty of IFN-γ to protect the animals against infection (7,8). These two temporal upregulations reveal that T cells responded to the vaccine, but the response did not sustain. We speculate that C-strain vaccine only triggers the cellular immunity but does not induce strong response. In such a situation, C-strain virus could replicate slowly and stimulate the immune system continually.
In conclusion, we speculate that the poor infectivity and slow replication abilities are important reasons for the attenuation of C-strain. C-strain virus replicates very slowly in limited tissues, so it could escape the recognition of TLRs temporarily, avoiding the untimely overexpression of IFNs and pro-inflammatory cytokines and protecting the immune cells and organs indirectly. When it accumulates to a certain extent, virus induces the expression of TLRs which initiates the innate immune system and then virus is cleared up. Meanwhile, ILs expressed by immune system induces the differentiation of B cells and production of specific antibody.
Footnotes
Acknowledgments
This work was supported by the “National Natural Science Foundation of China” (Grant No. 31372434) and Projects in the National Science and Technology Pillar Program during the Twelfth Five-year Plan Period (No. 2015BAD12B04).
Author Disclosure Statement
No competing financial interests exist.