
Abstract
Porcine epidemic diarrhea virus (PEDV) causes acute diarrhea, dehydration in pigs, and high mortality rates in piglets <3 weeks of age. Maternal immunity protects piglets, but information on vaccination before or after natural infection in endemically exposed sow herds is limited. Currently, the recovery goal in sow units infected with PEDV is to become fully naive again or use natural virus infection to develop immune gilts through a feedback program before introduction into the sow herd. Since neutralizing antibodies in the gut are critical for protection against enteric viral infections such as PEDV, we evaluated the effect of a conditionally licensed, adjuvanted inactivated PEDV vaccine on neutralizing antibody levels in milk and colostrum in both naive and previously naturally exposed sow herds. The results illustrate that intramuscular vaccination increased neutralizing antibody titers, and anti-PEDV IgA and IgG in milk and colostrum of sows that were previously infected. Thus, inactivated PEDV vaccines may provide increased protection to piglets nursing on previously infected sows against exposure to PEDV through increased delivery of lactogenic neutralizing antibodies to the enteric site of infection.
Introduction
P
PEDV replicates in the cytoplasm of epithelial villous cells of the small intestine (11). Clinical signs during PEDV infection include diarrhea, anorexia, depression, vomiting, and dehydration, and may vary depending on the previous exposure history and age of piglets (11). Diagnosis can be made from immunohistochemistry of small intestines or reverse transcriptase-polymerase chain reaction (RT-PCR) on tissue from the small intestine or fecal material shortly after infection (8,10). It has historically been spread by fecal–oral transmission and has been shown in proof-of-concept studies to be spread in feed (6). Infectious material was detected in air samples in air spaces with affected pigs, and genetic material was detected in air samples up to 10 miles downwind of a naturally infected farm (1). An effective amount of time and temperature that is required to inactivate virus is 80°C for 10 min or 20°C for 7 days (22). PEDV can be inactivated by standard disinfection processes; however, organic material will reduce inactivation efficacy (22). In Asia, chicken egg yolk immunoglobulin (IgY) was shown to decrease the mortality rates of piglets infected with PEDV, but this approach has not been widely utilized in the United States (13).
Feedback of fecal material from positive piglets to sows is effective in stimulating intestinal immune response and lactogenic immunity in sows; however, it presents the possibility of disease outbreaks within a herd. Transmission occurs readily within a facility by fecal contamination of boots, equipment, and other fomites. PEDV is a difficult disease to prevent from entering a sow farm, although external biosecurity improvements have been developed in the swine industry. Therefore, there is a need for an efficacious vaccine that stimulates a strong immune response without a threat of disease.
At this time, an efficacious vaccine for PEDV and many other porcine enteric viral pathogens is lacking (7). However, a vaccine product that boosted pre-existing maternal immunity from a previous natural infection could help in providing better protection to nursing piglets. In this article, we hypothesized that intramuscular administration of an inactivated PEDV vaccine would boost lactogenic neutralizing antibodies, an essential component of neonatal immune protection against enteric viruses, in previously infected sows.
Materials and Methods
Study design: naive breeding herd
A 1,000 sow, single site, farrow-to-finish breeding herd with no history of PEDV exposure was identified in the Midwestern United States as the source of naive sows. Sows were bred and gestated in pens and farrowed in stalls. Twenty pregnant sows were vaccinated two times intramuscularly at 5 weeks prefarrowing and again at 2 weeks prefarrowing. Twenty naive, negative control sows were injected with water at the same time. Each group consisted of 2 gilts and 18 sows of parity one or older.
Immune breeding herd
A second Midwestern U.S. 2,200-sow breed-to-wean herd was exposed to PEDV 7 months before study initiation and was the source of sows with pre-existing immunity. These breeding swine were bred, gestated, and farrowed in stalls. Twenty sows were vaccinated intramuscularly with two doses of PEDV-inactivated vaccine prefarrowing as already described. Twenty randomly selected sows were injected with two doses of sterile water as negative controls. Each group consisted of 2 gilts and 18 sows of parity one or older.
Before first vaccination, serum samples were collected from all sows in both groups in each herd. After farrowing, serum and colostrum samples were collected at 24 h, and milk samples were obtained between days 3 and 7. All other farm procedures followed were as normal/routine for the farm.
Vaccination and sample collection
Inactivated PEDV formulated with adjuvant was provided by Zoetis (Parsippany-Troy Hills, NJ) and used at a dose of 2 mL per sow intramuscularly in the neck at 5 and 2 weeks prefarrowing. Control animals received 2 mL sterile water intramuscularly at the same time. Serum was collected by venipuncture with restraint into Vacutainer serum separator tubes. Colostrum and milk samples were collected by expression of mammary glands into sterile tubes. Oxytocin (10 USP units per 0.5 mL dose; Bimeda-MTC Animal Health, Cambridge, Canada) was used if needed for milk let-down. Samples collected were <20 per group because of animal health and farm management reasons, including farrowing when personnel were not available to collect samples.
Clinical signs
Veterinary heath evaluation for PEDV was conducted by visual examination for diarrhea. Evaluation for adverse vaccination events was carried out by the veterinarian based on observation of animals for abnormal movement or behaviors after administration. The occurrence of injection site reactions was evaluated by visual observation at 1–2 days after administration.
PEDV RT-PCR
Environmental and fecal samples were submitted to the University of Minnesota Veterinary Diagnostic Laboratory for testing of PEDV RNA by RT-PCR.
Antibody isotype enzyme-linked immunosorbent assay
Total antibody isotype levels for IgG and IgA were determined by sandwich enzyme-linked immunosorbent assay (ELISA) in colostrum and milk. For ELISA, 96-well plates were coated with 500 ng affinity purified goat anti-IgG or anti-IgA (Bethyl Labs, Montgomery, TX), washed, incubated with serum (1:40,000 dilution for IgG, 1:50,000 dilution for IgA), colostrum (1:500,000 dilution for IgG, 1:120,000 for IgA), or milk (1:2,500 for IgG, 1:30,000 for IgA) for 1 h, washed, and reacted with horseradish peroxidase (HRP)-conjugated goat antiswine IgG or antiswine IgA diluted 1:100,000 for 1 h. Plates were washed and tetramethylbenzidine color development (KPL, Gaithersburg, MD) was performed for 15 min before reactions were stopped with 1 M phosphoric acid. Plates were read at 450 nm on an Epoch Absorbance Microplate Reader (BioTek Instruments, Winooski, VT). All steps were carried out at room temperature. Immunoglobulin concentrations were determined by comparison with a standard curve provided in the kit and run on the same plate, then adjusted for dilution.
Viral neutralization assay
The PEDV neutralization assay was performed essentially as described previously with modifications (14,16). Milk and colostrum samples were threefold serially diluted from 1:100 to 1:218,700 and incubated with PEDV at a multiplicity of infection of 0.01 for 1 h at 37°C. Confluent Vero-81 cell monolayers in 96-well plates were washed with phosphate-buffered saline (PBS) before mixtures of sample and virus, positive control virus, and blank controls were added in duplicate. After 1 h incubation at 37°C, plates were washed and incubated at 37°C in prewarmed media. After 24 h, plates were washed and fixed in 3.7% formaldehyde for 30 min, permeabilized with 0.1% Triton X-100 for 5 min, washed, and blocked with 5% nonfat dry milk in 50 mM carbonate buffer, pH 9.5, for 1 h at 37°C. Cells were incubated with anti-PEDV N protein monoclonal antibody (SD6-29) (Medgene Lab, Brookings, SD) diluted 1:1,000 in PBS containing 0.05% Tween-20 (PBST) for 1 h at 37°C, washed, and incubated with HRP-conjugated goat antimouse IgG heavy and light chains antibody (Bethyl Laboratories) diluted 1:10,000 in PBST. Plates were washed with PBST and developed using TMB development solution (KPL). Reactions were stopped with 1 M phosphoric acid and absorbance was measured at 450 nm by a spectrophotometer (BioTek). The concentration-dependent response curve was modeled by a sigmoidal four-parameter logistic fit of nonlinear regression models using GraphPad Prism 5.0 (GraphPad Software, San Diego CA). The background absorbance values varied in a dilution-dependent manner from 0.58 ± 0.25 (mean ± standard deviation) at a 1/100 serum dilution to 0.06 ± 0.01 at a 1/218,700 dilution. Percentage inhibition of viral infection at each dilution was compared with virus-only controls after background subtraction of absorbance from uninfected cells. Fifty percent neutralization titer was defined as the reciprocal of the highest serum dilution for which 50% inhibition of infection was reached.
Polypeptide-based ELISA
Polypeptides encoding PEDV nucleocapsid (N), membrane (M), and S2 fragment of the spike protein were produced and purified as described (18). Ninety-six-well EIA/RIA plates (Corning Incorporated, Corning, NY) were coated with 100 ng/well of recombinant polypeptides in carbonate buffer, pH 9.6, at 4°C overnight. Plates were washed three times with PBST (Fisher Scientific, Waltham, MA) and blocked with 5% nonfat dry milk in PBST, pH 9.6, for 1 h. Serum samples were diluted 1:50 in 5% nonfat dry milk (pH 7.4) in PBST, and each sample was applied in 100 μL to duplicate wells of an ELISA plate for 1 h. Plates were washed three times with PBST, and HRP-conjugated goat antiswine IgG or IgA (KPL) (1:5,000 dilution) was added for 1 h. Plates were washed and color development was performed with 100 μL of TMB peroxidase substrate (KPL) for 15 min. Reactions were stopped with 100 μL of 1 M phosphoric acid and plates were then read at 450 nm in a Thermo Max Microplate Reader (Molecular Devices, Sunnyvale, CA). All steps were carried out at room temperature.
Statistical methods
Treatment comparisons were analyzed by t-test or Mann–Whitney U test. Significance was set at p ≤ 0.05.
Results and Discussion
Clinical signs
The lack of clinical signs in the 2,200 sow herd before the start of the study as well as negative PCR tests from both environmental and fecal samplings supported the reasoning that PEDV had been eliminated from this site after the initial outbreak. The animals enrolled into the study did not have any adverse reactions to administration of the PEDV vaccine. All sows were reported on feed the day after the injection of vaccine. All sows continued to be clinically normal through their farrowing process and no adverse observations were documented in their offspring.
Effect of vaccination on neutralizing antibody responses
Intramuscular vaccination of previously infected, immune sows with inactivated PEDV at 5 and 2 weeks prefarrowing significantly increased viral neutralizing activity in colostrum (p < 0.0001) and milk (p = 0.0005) compared with nonvaccinated controls (Fig. 1A). The average titer in nonvaccinated sows was ∼1/20,000 in colostrum and 1/8,000 in milk. The average titers in vaccinated sows were ∼fivefold higher. Intramuscular vaccination of naive sows in a PEDV-negative herd induced viral neutralization activity in colostrum that was highly variable, ranging in titer from 1/100 to 1/7,000 (Fig. 1B). However, there was no effect of vaccination on neutralizing activity in milk, except for one outlier (Fig. 1B, solid triangles). Colostrum and milk from nonvaccinated naive animals lacked neutralizing antibodies, except for one outlier milk sample (Fig. 1B, open triangle). Increased neutralizing activity was because of anti-PEDV antibodies produced in response to vaccination, because total immunoglobulin levels were equivalent regardless of group or treatment, as shown in Table 1. Total IgG in the immune, vaccinated group was numerically higher, due primarily to two high values (9 and 40 mg/mL), as indicated by the large standard error. In previous, unrelated studies of isotype-specific immunoglobulin concentrations in sow serum, colostrum, and milk, the levels of IgG in milk were substantially higher than observed here, whereas the levels of IgA in milk were similar (Table 2). IgG levels in milk also showed a high level of variation compared with IgA. The variation suggests that the duration of IgG-rich colostrum delivery may be variable and prolonged beyond 24–48 h depending on the timing of fully developed lactogenesis (12).
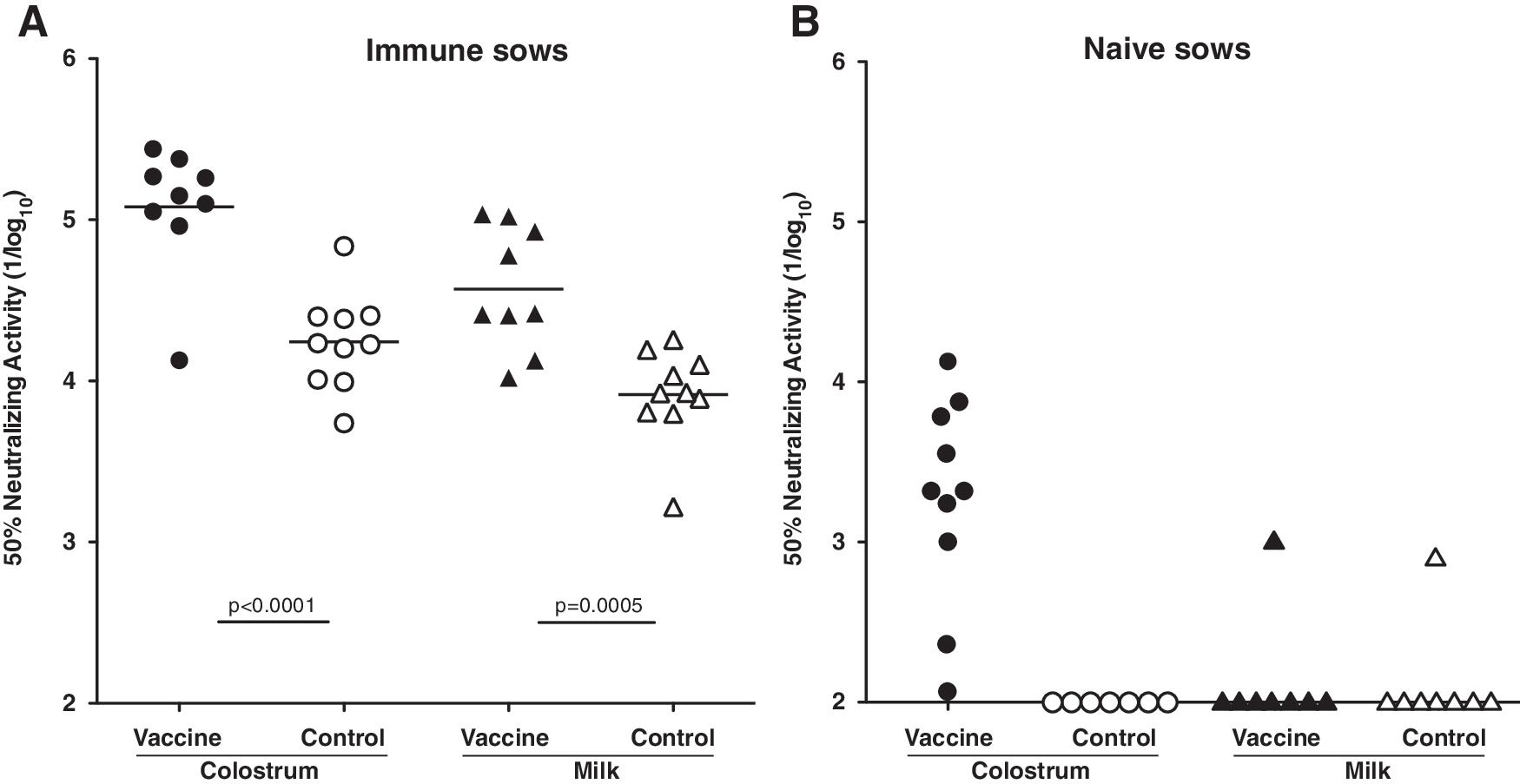
Effect of vaccination on neutralizing activity.
Concentration (mg/mL, median ± standard error).
Values are mean ± standard deviation (mg/mL).
Effect of vaccination on antibody levels in naive, nonimmune sows
ELISA antibody levels reactive with nucleocapsid (N), spike protein domain S2 (S2), and membrane protein (M) were measured in serum, colostrum, and milk to gain further insight into the impact of prefarrowing vaccination on maternal transfer of neutralizing activity. Antibody responses to the S1 spike protein domain were not assessed because the recombinant antigen was not expressed well. In naive sows, two doses of inactivated vaccine at 5 and 2 weeks before farrowing significantly increased IgG antibody responses in serum (p = 0.002, Fig. 2) and colostrum (p = 0.003, Fig. 3). However, vaccination did not induce a significant response to N in colostral IgA, milk IgG, or milk IgA (Fig. 3). Vaccination has no effect on antibodies to N, M, or S2 in serum (Fig. 2) or colostrum and milk (Fig. 3). p Values varied between 0.08 and 0.94. These results indicate that parenteral vaccination induced a systemic IgG response detectable in serum and colostrum but failed to elicit antibodies to structural proteins in colostrum or milk that could confer protection in the gut. It also suggests the possibility that the vaccine product may be enriched in N because the dominant response was to this protein.
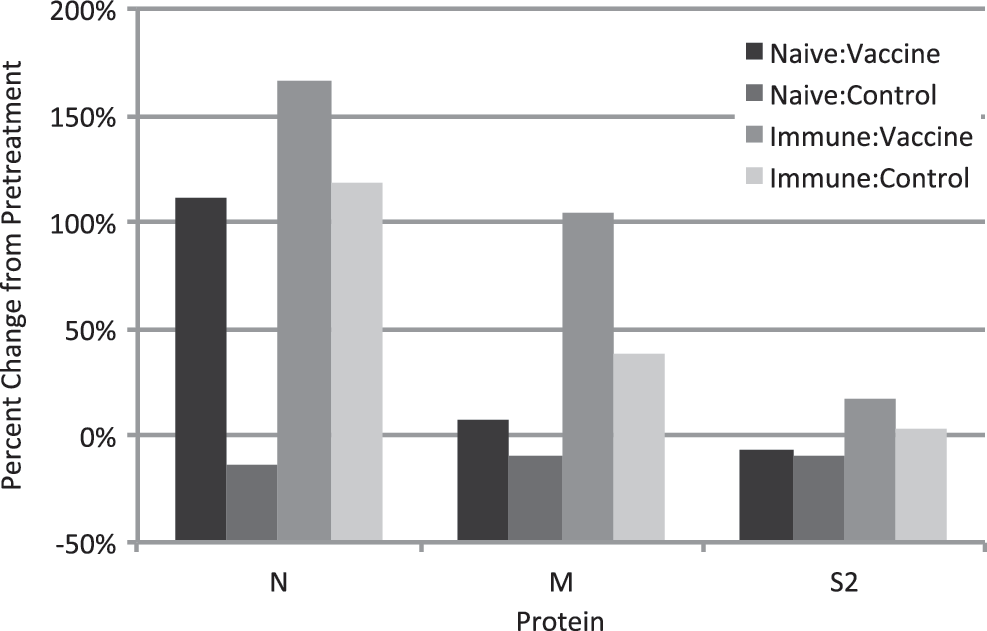
Effect of vaccination on anti-PEDV antibody responses in serum. Data are the percentage change in ELISA absorbance value in sow serum taken before the first treatment administration and day after farrowing. ELISA, enzyme-linked immunosorbent assay; PEDV, porcine epidemic diarrhea virus.
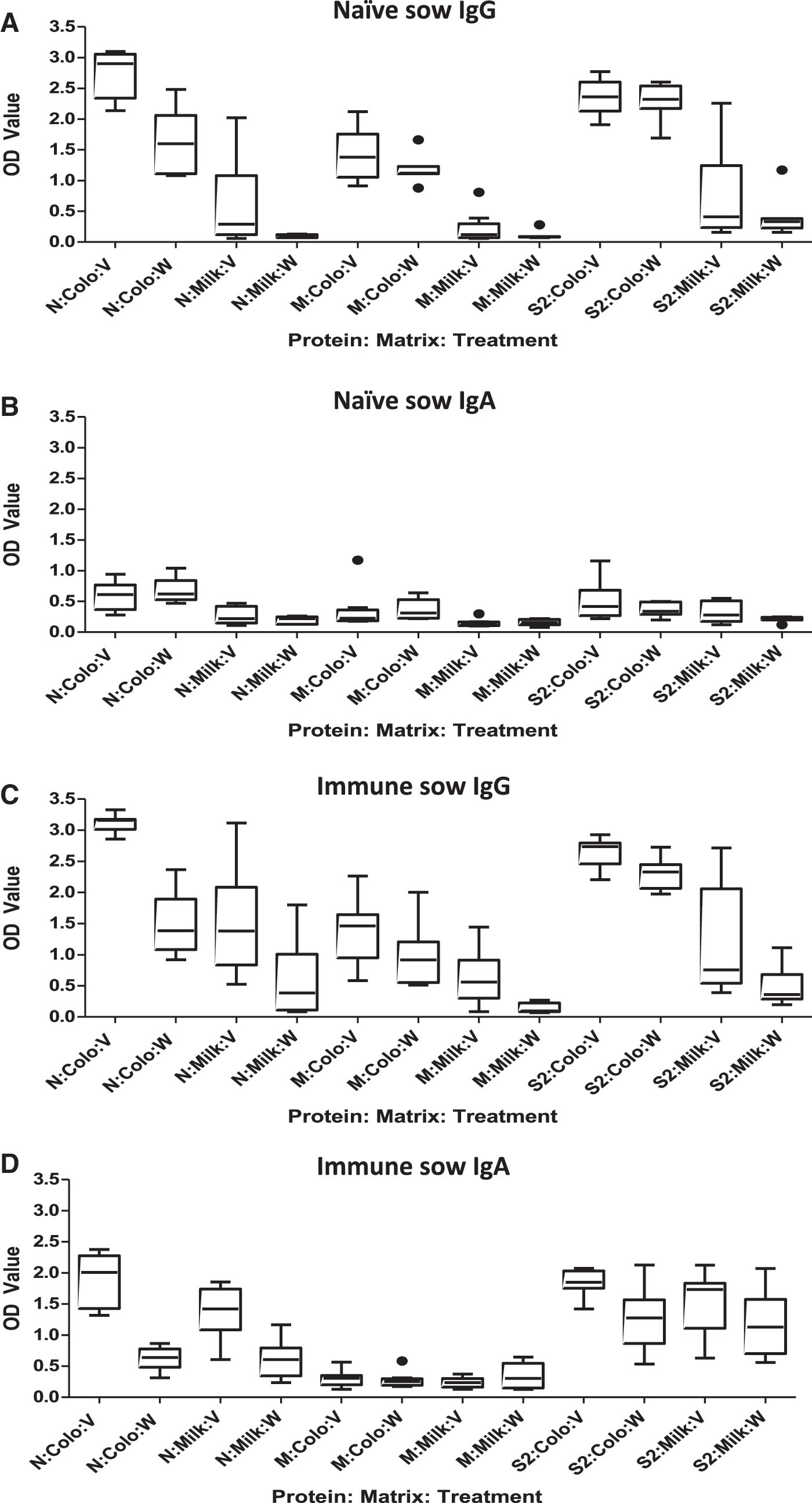
Effect of vaccination on anti-PEDV antibody responses in colostrum and milk. Data are shown as box and whisker plots in which the horizontal line represents the median, the box represents the 25th and 75th percentiles, and the whiskers represent the 5th and 95th percentiles. Dots represent outliers. On the X-axis in all panels, protein is N (nucleocapsid), M (membrane), and S2 (spike protein S2 fragment); treatment is V (vaccine) or W (water).
Effect of vaccination on antibody levels in previously infected sows
In immune sows that were previously exposed, vaccination induced immune responses detected in serum, colostrum, and milk. In serum, anti-M IgG was significantly increased (p = 0.0009). IgG antibody levels to N and S2 were numerically but not significantly increased (p = 0.20 and 0.08, respectively, Fig. 2). In colostrum, IgG and IgA antibodies to N and S2 were significantly increased (p ≤ 0.005 in all cases), but there was no significant response to M (Fig. 3). In milk, IgG antibodies were significantly increased to all three proteins (p ≤ 0.016), and IgA antibodies were increased for N only (p = 0.001) (Fig. 3). There was no effect of parity on antibody responses in naive or immune sows.
Systemic versus maternally derived antibody responses
The serological IgG responses were consistent with the systemic presence of PEDV-specific memory B cells in previously exposed sows that responded to restimulation because of vaccination, whereas in naive sows lacking PEDV-specific memory B cells, intramuscular vaccination was only partially successful in increasing circulating antibodies. IgA levels in serum were not tested because they were low in previous studies, comprising ∼2% of total immunoglobulins (Table 2).
Colostrum was dense and creamy compared with milk, was dominated by IgG isotype, and had a total immunoglobulin concentration in the range of 100 mg/mL. In practice, colostrum was collected at 24 h after farrowing. Milk was collected 3–7 days after farrowing, was watery, dominated by IgA isotype, and the total immunoglobulin concentration was in the range of 10 mg/mL. Collecting single samples of colostrum and milk may have contributed to variation in the results. For example, kinetic analysis of PEDV-neutralizing activity in sow colostrum and milk showed variation in neutralizing activity among animals and over time during lactation (18).
Maternally derived antibodies in colostrum are primarily of the IgG isotype and are derived from serum, whereas antibodies in milk are largely IgA isotype and are produced locally by plasma cells in the mammary gland (3). Thus, isotype analysis in colostrum and milk may give insights into the effect of intramuscular immunization with inactivated PEDV vaccine.
Intramuscular vaccination at 5 and 2 weeks induced IgG isotype antibodies in colostrum more so than in milk (Fig. 3A). By contrast, IgA isotype anti-PEDV antibodies were not significantly induced in colostrum or in milk (Fig. 3B). It appeared that intramuscular administration of inactivated PEDV induced a systemic immune response in naive animals that resulted in production of circulating IgG that was extracted from blood and concentrated in colostrum. It did not stimulate an IgA response in secretory tissue of the mammary gland for delivery into milk. A humoral response dominated by IgG and compartmentalized in systemic lymphoid tissue with limited impact at mucosal sites such as the gut is typical for a nonlive vaccine administered intramuscularly (2).
In previously infected, immune sows that have anti-PEDV immunological memory, vaccination markedly increased IgG antibodies to all PEDV structural proteins in both colostrum and milk (Fig. 3C). Vaccination also enhanced IgA isotype responses against N and S2 in both colostrum and milk (Fig. 3D). An increase in antigen-specific IgA in milk indicates that parenteral administration of antigen in the presence of immune memory is able to elicit localized B cell responses in the mammary gland. The mechanism by which it is accomplished remains to be determined.
Natural infection of sows with PEDV, an enteric pathogen, induces IgG antibodies in serum and secretory antibodies in mammary secretions that help protect suckling piglets. Systemic IgG is derived primarily from B cells in spleen and lymph nodes, whereas secretory IgA is produced locally in gut tissues and mammary glands (3, 4). The purpose of this study was to determine whether intramuscular administration of a nonreplicating vaccine antigen, which typically elicits a systemic response, could increase colostral and lactogenic antibodies with neutralizing activity to better protect piglets against PEDV. A successful outcome would help biosecurity by reducing the need for virulent virus infection, which introduces infectious virus into the sow herd. It also would show that immunity induced in a previous reproductive cycle might be enhanced in the absence of virulent virus.
The results here show that parenteral administration of an inactivated PEDV product elicits an immune response that is evident systemically in serum of naive sows and in colostrum. However, the immune response does not elicit neutralizing antibody responses in milk and only weakly in colostrum of previously unexposed sows. We conclude that parenteral administration of inactivated PEDV to naive sows is not likely to provide protective neutralizing antibodies to piglets.
A more robust antibody response occurred in immune sows. The parenteral administration increased anti-PEDV IgA levels in milk and colostrum, as well as IgG antibody levels in serum and colostrum. Neutralizing activity present in colostrum and milk of immune sows was significantly increased. Therefore, boosting existing immunity from prior infection with parenterally administered, inactivated virus may increase protection in nursing piglets against PEDV. Although vaccination in previously exposed sows was shown to increase maternally derived PEDV-neutralizing activity, the levels of activity needed to control or prevent neonatal infection are not yet known and might be dependent on piglet consumption.
Conclusions
The findings in this study show that intramuscular vaccination with an inactivated PEDV vaccine boosts existing colostral and lactogenic immunity 4–5 months after natural PEDV infection. Parenteral vaccination of naive sows without pre-existing immunity induced a serological antibody response and a low level of colostral neutralizing activity, but no response in milk. An infectious challenge study is needed to determine whether a parenterally administered inactivated virus preparation protects sows or piglets against virulent virus exposure.
Footnotes
Acknowledgments
Funding for the animal studies was provided by the USDA through the Indiana Board of Animal Health. Laboratory studies were supported at the University of Minnesota, in part, by a China Scholarship Council award to Q.S.
Author Disclosure Statement
No competing financial interests exist.