
Abstract
Antigen-specific immune responses determine the efficacy of herpes simplex virus type 2 (HSV-2) vaccines. To optimize the immunogenicity of the antigen gD2, we developed the gD2ΔUL25 DNA vaccine encoding HSV-2 glycoprotein D and UL25 gene encoding viral capsid vertex proteins in this study. The gD2 and gD2ΔUL25 DNA vaccines were compared with formalin-inactivated HSV-2 (FI-HSV-2), and results showed a greater protective immune response induced by gD2ΔUL25 than by gD2. Therefore, gD2ΔUL25 was chosen to evaluate further using the IL28B adjuvant. Immunization with gD2ΔUL25/IL28B elicited stronger humoral and T cell immune responses than with gD2ΔUL25 alone. Compared with controls, gD2ΔUL25/IL28B decreased HSV-2 viral loads and induced protective effects against genital tract lesions generated by HSV-2. These findings demonstrated that the prophylactic DNA vaccine gD2ΔUL25 with IL28B adjuvant could enhance the humoral and T cell immune responses, and improve the protective immune response against HSV-2 in female mice compared with FI-HSV-2.
Introduction
H
Unfortunately, no commercial vaccine candidate for HSV-2 is currently available, and developing a safe and effective vaccine is imperative. Many studies have reported on outcomes of prophylactic and therapeutic vaccine approaches against HSV-2, including changing viral targets, altering vaccine platforms, adjusting inoculation dosages, and application of adjuvants. However, all vaccine candidates have failed in clinical trials to date. The HSV-2 glycoprotein gD2/gB2 subunit vaccine with the MF-59 adjuvant was demonstrated to induce strong neutralizing antibody titers in both prophylactic and therapeutic HSV-2 vaccine clinical trials, but it provided no protection from HSV-2 infection based on seroconversion (25). The gD2-subunit vaccine with MPL-alum adjuvant is currently in a large Phase III trial in HSV-1 and HSV-2 doubly seronegative women, and this strategy improved only the prevention of HSV-1 genital infection (6). The gH deletion virus vaccine (single cycle) and ICP10DPK replication competent virus both failed in Phase II trials with no significant effects on recurrence (10,14). Studies of other vaccines entering Phase I/II trials are also under way, such as the gD2-ICP4 subunit vaccine with M2 Matrix adjuvant, as well as the replication-defective HSV529 virus and plasmid DNA vaccine with Vaxfectin adjuvant (14).
Vaccination with plasmid DNA is a novel strategy for protection from HSV-2 infection. Unlike traditional vaccines, DNA vaccines encode antigen proteins for expression in eukaryotic cells. They are easy to manufacture with high yield and are safe and stable (11). In this study, we chose the DNA vaccine platform and the antigen proteins HSV-2 gD2 and UL25. The gD2 protein binds to herpes virus entry mediator HveA of host cells directly to mediate HSV-2 entry (13). As it is an essential viral molecule, gD2 epitopes have been used in many vaccine studies as the viral target and shown to induce strong neutralizing antibodies that block infection (39). In this study, another chosen target encoded by the UL25 gene is the capsid protein located on the viral capsid vertices and is associated with pentons (32). HSV-2 UL25 contains T cell epitopes that have been demonstrated to elicit antigen-specific CD8+T cell responses (22,31). Both gD2 and UL25 epitopes can stimulate the host to produce humoral and cellular immune responses. While both types of immunity can play an important role in antiviral defense, cell-mediated immune responses, especially antigen-specific CD8+ T cells, are necessary for the elimination of replicating virus and have been shown to reduce HSV-2 latency in some sensory neurons (37). Meanwhile, antigen-specific CD4+ T cells can provide helper functions to facilitate antibody-dependent cell-mediated cytotoxicity of virus-infected cells and secrete high levels of interferon gamma (IFN-γ) (41).
Although DNA vaccines alone are known to be weakly immunogenic, their effects can be enhanced by adjuvants such as the IL28B cytokine. IL28B was shown to potentiate both cellular and humoral responses to immunogens in mice, representing a promising molecular adjuvant (18). In the early characterization of IL28B, the cytokine was demonstrated to have an antiviral activity against a number of viruses. Subsequent studies with administration of recombinant IL28B to mice or expression of IL28B by a recombinant virus demonstrated an antiviral activity of IL28B (24). The antiviral activity of IL28B against vaginal HSV infection was shown to be dependent on IL-28Ra expression only on nonhematopoietic cells, and in this compartment, vaginal epithelial cells responded to IL28B, which directly restricted virus replication. These reports showed that recombinant IL28B could potently inhibit HSV replication in a murine model for herpes genitalis (16).
In our study, three plasmids, pgD2, pgD2ΔUL25, and pΔUL25, were constructed, and their immunogenicities were compared with that of formalin-inactivated HSV (FI-HSV) in the first experiment. In the subsequent experiment, the potential of pIL28B to enhance the immunogenicity of the DNA vaccines in BALB/c mice also was investigated.
Materials and Methods
Animals
In this study, female BALB/c mice aged 6–8 weeks were purchased from Changchun Institute of Biological Products. All animal studies were approved by the University Committee on the Use and Care of Animals of Jilin University.
Cells, media, and viruses
African green monkey kidney (Vero) cells (ATCC CCL-81, Manassas, VA) and human embryonic kidney 293 (HEK293) cells (cat#114 CRL-1573; ATCC) were grown and maintained in Dulbecco's modified Eagle's medium containing 10% (vol/vol) fetal bovine serum, 100 U/mL penicillin G, and 100 μg/mL streptomycin sulfate for use. Vero cells were cultured in a 37°C incubator with 5% CO2.
HSV-2 strain G (cat# VR-734; ATCC) was propagated by infection of confluent Vero cell monolayers at a multiplicity of infection of 0.1 in a 34°C incubator with 5% CO2 for 72 h. At harvest, most cells showed cytopathic effects (CPEs; 90–100%) as previously described (15). The HSV-2 titer determined on Vero cells was 1 × 108 plaque-forming units (PFU)/mL. A portion of the virus stock was stored at −80°C for challenge experiments, while another portion was prepared as inactivated HSV-2 viruses (FI-HSV-2) as a positive control. This latter portion of the virus was purified by discontinuous sucrose gradient centrifugation at 60,000 g for 3 h at 4°C and inactivated in 0.1% formaldehyde for 72 h at 37°C before storage at −80°C.
Peptides
In this study, HSV-2 gD253-65 (LTDPPGVKRVYHI) (30) and UL25364-372 (FLARGHNLF) peptides (5 μg/mL) were used. The UL25364-372 sequence was determined to be a CD8+ T cell epitope based on analysis with the SYFPEITHI algorithm (Institute for Cell Biology, Heidelbeng, Germany) and predicted to bind to H-2d MHC-I molecules.
Construction of recombinant DNA plasmids and adjuvant
A human codon-optimized nucleotide sequence was derived from the HSV-2 HG52 amino acid sequence (GenBank accession number: Z86099.2). The gD2ΔUL25 fusion gene fragment, including the gD2 (amino acids 26 to 306) and ΔUL25 (amino acids 310 to 585) with a flexible peptide (Gly4Ser)3 linker gene between gD2 and ΔUL25, was synthesized and the plasmid was named pGH (plasmid of genital herpes)-gD2ΔUL25 (GENTEC, Shanghai, China). The gD2, ΔUL25, and gD2ΔUL25 (fusion gene) sequences were polymerase chain reaction (PCR) amplified with the template pGH-gD2ΔUL25 and primers (Table 1) and cloned into the vector pcDNA3.1(+) to generate plasmids pgD2, pΔUL25, and pgD2ΔUL25, respectively (Fig. 1A–C). A human codon-optimized nucleotide sequence encoding IL-28B (GENTEC) was also synthesized and PCR amplified, and cloned into the pVAX1 plasmid backbone to generate the plasmid pIL28B (Fig. 1D).
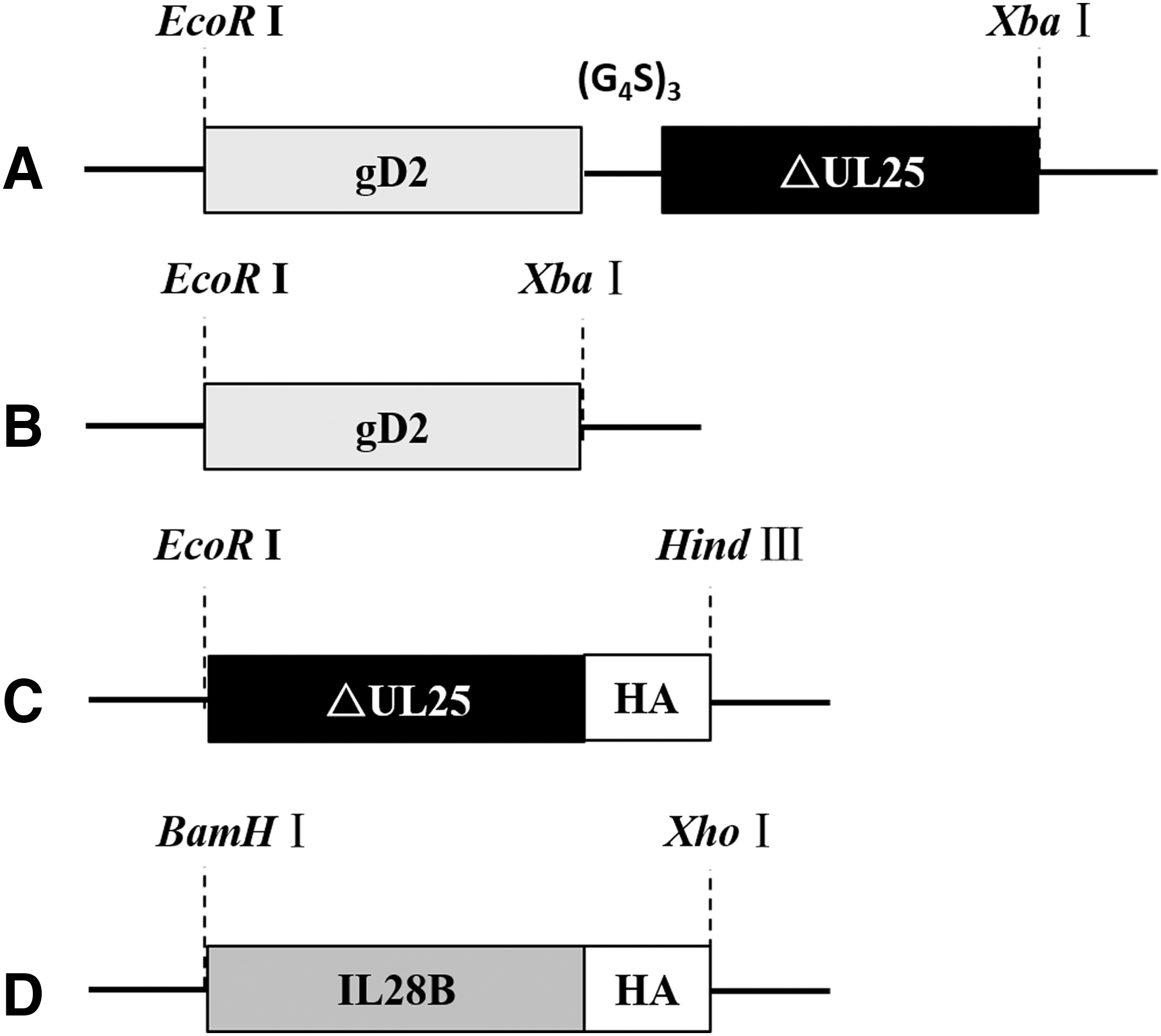
Construction of recombinant DNA plasmids and adjuvant.
Eukaryotic expression of gD2, ΔUL25, gD2ΔUL25, and IL-28B
Purified plasmids were transfected into confluent HEK293 cell monolayers by using Lipofectamine 2000 and incubated at 37°C with 5% CO2 for 3 days (17). HEK293 cells were harvested when most cells showed CPEs, and the lysates were obtained by freeze-thawing cells twice. Proteins from cell extracts were determined by Western blotting. Expressed antigens (gD2, UL25, and gD2ΔUL25) and cytokine adjuvant protein (IL-28B) were separated by electrophoresis on a 13.5% sodium dodecyl sulfate–polyacrylamide gel under denaturing conditions (12). Separated proteins were transferred to a nitrocellulose filter. The gD2 polyclonal antibody produced by our laboratory and 1:1,000 anti-HA tag antibody (Abcam, Inc., Shanghai, China,) were used for Western blot assays as described previously (26).
Mouse immunizations and viral challenge
Thirteen female BALB/c mice were randomly assigned to each group. To evaluate the immune responses to the pgD2, pgD2ΔUL25, and pΔUL25 constructed in this study, they were compared with FI-HSV-2 as a positive control in experiment 1 (Table 2). To determine if IL28B could enhance the immune responses to these DNA vaccines, mice were immunized as shown in Table 3 for experiment 2 (Table 3). Each vaccine was administered by intramuscular or subcutaneous injection of the left and right hind limbs in a volume of 50 μL per injection (7,9). All animals were immunized thrice, 14 days apart. The immunized animals were bled from saphenous veins on the 10th day after every immunization, and serum samples were collected and stored at −80°C. Five mice of each group were sacrificed at 2 weeks after the last immunization.
FI-HSV-2, formalin-inactivated herpes simplex virus type 2; PBS, phosphate-buffered saline.
Mice were injected subcutaneously with medroxyprogesterone acetate (Pharmacia Upjohn, Kalamazoo, MI) at a dose of 2 mg/animal a week before viral challenge. All animals were anesthetized and challenged intravaginally with lethal dose 1 × 105 PFU (5 × LD50)/animal of the HSV-2 G strain 2 weeks after the third immunization. The immunization and viral challenge experiments were repeated thrice.
Clinical observations
The mice were scored daily for 20 days after challenge for the severity of vaginal disease according to a previously described (38) scoring system: 0 = no lesions; 1 = redness or swelling; 2 = several vesicles; 3 = urine or fecal retention and hind limb paralysis; and 4 = moribund. Disease scores were recorded every day postchallenge. To confirm vaginal viral shedding, vaginal swab samples were taken with a nylon-tipped applicator to collect vaginal secretions every 2 days.
Antigen-specific antibody titers
Antibody titers were detected by standard antigen-specific enzyme-linked immunosorbent assay (ELISA). In this study, HSV-2 gD26-306 protein was used to coat 96-well plates at 100 ng/well diluted in a coating buffer. The gD26-306 protein was expressed from Escherichia coli and purified using the Ni-NTA affinity column (Invitrogen, Carlsbad, CA) under denaturing conditions. Mouse serum samples were threefold serially diluted with the first dilution at 100-fold. Absorbance values in ELISAs were obtained as previously described (33). The endpoint titer was defined as the reciprocal of the serum dilution, when the absorbance of samples at 450 nm was twice that of the negative control.
Neutralization analysis
The neutralization capacity was tested in a microneutralization assay as previously described (19). Briefly, serum from all mice in each group was heat inactivated (30 min, 56°C), and 200 PFU of the HSV-2 strain G was mixed with twofold serially diluted serum samples at a 1:20 starting dilution. Each sample was tested in duplicate on Vero cell monolayers. The neutralizing antibody titer was defined as the reciprocal of the highest plasma or serum dilution that showed no CPE compared with the virus control.
Enzyme-linked immunospot assay
Two weeks after the last immunization, mice were sacrificed and their spleens were collected to prepare single-cell suspensions. The splenocytes were added at 1 × 106 cells/well with HSV-2 gD253-65 (LTDPPGVKRVYHI) and UL25364-372 (FLARGHNLF) as stimulating peptides to 96-well filter plates that were precoated with purified anti-mouse IFN-γ mAb (BD 51-2525KC). Spotting-out was processed according to the manufacturer's protocol (BD Biosciences, Franklin Lakes, NJ). IFN-γ spots stimulated by peptides in the enzyme-linked immunospot (ELISPOT) are presented as the number of spot-forming cells (SFC) per 106 splenocytes.
Multiplexed microsphere cytokine assay
Results of the multiplexed microsphere cytokine assay for detection of Th1/Th2 cytokines were analyzed according to the manufacturer's instructions for the kit (BioLegend) as previously described (19). In our study, we detected Th1 (IFN-γ and IL-2) and Th2 (IL-4, IL-5, and IL-10) cytokines secreted by each group of splenocytes stimulated with the gD2 protein. Briefly, 1 × 107 cells/well of splenocytes were added to a sterile 96-well round-bottom plate and incubated with 2.5 μg of gD26–306 for 3 days. The culture supernatant was collected, and 100 μL of each sample was analyzed on the Luminex® 100 ISTM System (Luminex Corporation, Austin, TX).
Isolation of vaginally shed virus
Vaginally shed virus was isolated from swab samples that were suspended in 200 μL HASS buffer as previously described (42). The HSV-2 genome was extracted by using a DNA extraction Kit (Qiagen, Hilden, Germany) according to the instructions of the kit and stored at −20°C until analyzed.
Real-time quantitative PCR of viral DNA
The number of HSV-2 DNA copies was quantified by a Real-time Quantitative PCR Detecting System (Invitrogen) with the following primers specific for HSV-2 gG2 (glycoprotein G2) DNA: forward, 5′-CCCACACCCCAACACATC-3′; and reverse, 5′-CCAAGGCGAC CAGACAAAC-3′ (1). Tenfold serial dilutions of the gG2-Teasy recombinant plasmid were used as standards to quantify the HSV-2 copy number. Each 20 μL reaction was performed in duplicate and contained 2 μL of DNA,10 μL of 2× TransStart Top Green qPCR SuperMix, 0.5 μL of (each) forward and reverse primers, 0.4 μL of passive reference dye (50 × ), and 6.6 μL of ddH2O. The reactions were amplified for 40 cycles (30 sec at 94°C, 30 sec at 55°C). The PCR products were analyzed with the Bio-Rad CFX Manager™, version 2.1. The limit of detection was determined to be one copy per 20 μL reaction, with excellent linearity (R = 0.98) over 5 logs of HSV-2 genomic DNA content.
Statistical analyses
Statistical analyses were performed using ANOVA and Student's t-test with GraphPad Prism 5.0 software (GraphPad, San Diego, CA). All tests were three-tailed, and a p value <0.05 was considered to indicate a significant difference.
Results
Expression of recombinant DNA plasmids and IL28B adjuvant
Western blot analysis was performed to characterize gD2 and gD2ΔUL25 protein expression in extracts of HEK293 cells (Fig. 2A) using gD2 polyclonal antibody as the primary antibody. A band of ∼32 kDa was visualized that represented the gD2 protein (lane 2), and the other band of ∼65 kDa was detected that represented the gD2ΔUL25 protein (lane 3). Nontransfected HEK293 cells served as negative controls (lane 4). The ΔUL25 protein expressed in HEK293 cells also was confirmed by Western blot analysis, exhibiting a band of 32 kDa, as shown in Fig. 2B (lane 2). Adjuvant IL28B protein from the eukaryotic expression plasmid pGH-IL-28B was visualized as shown in Figure 2C. Culture supernatant of transfected HEK293 cells showed a band of 25 kDa in lane 2, while IL28B protein from cell extracts was detected at the same size. These results indicated that the DNA plasmids that transfected into HEK293 cells could express the corresponding proteins correctly, and recombinant proteins were expressed both intracellularly and extracellularly since the sequence of IL-28B contains a secretion signal peptide (bases 1 to 57).
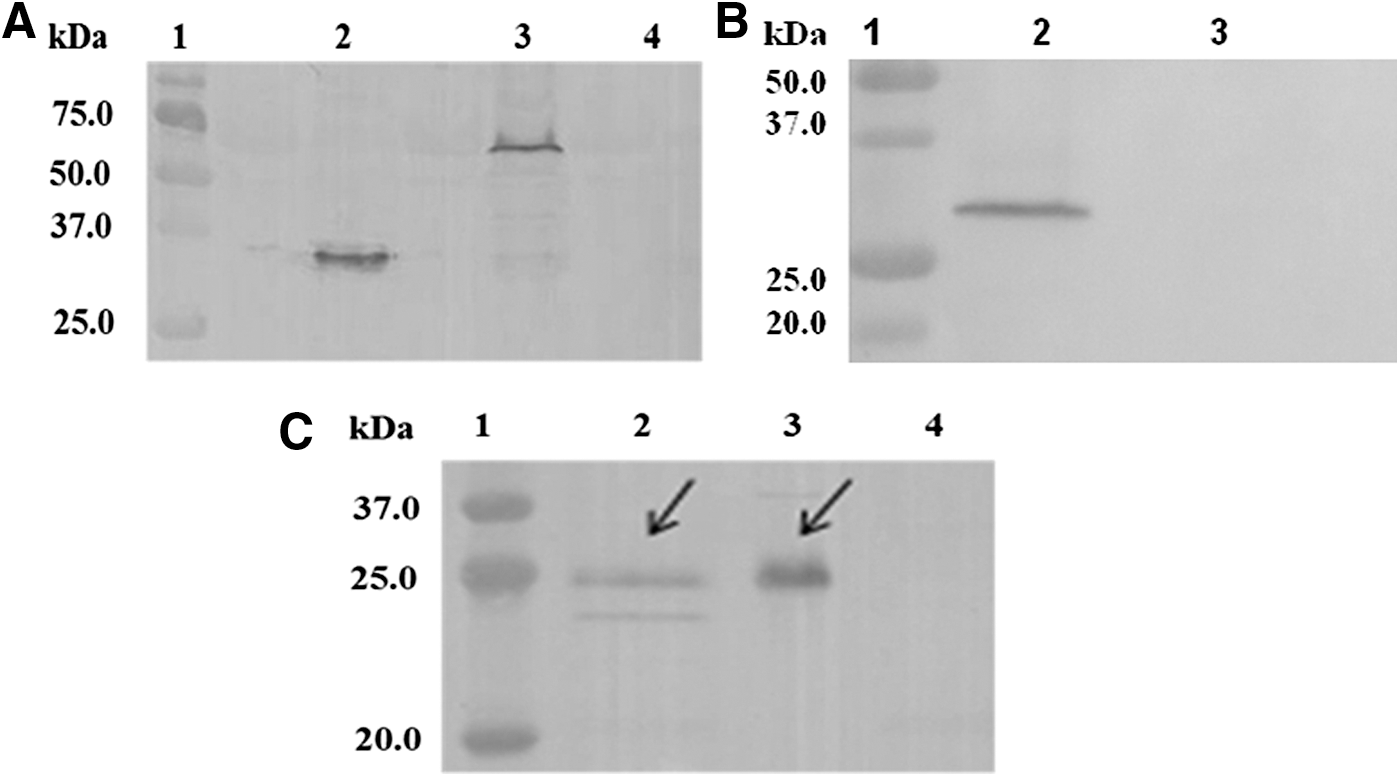
Western blot analysis of expression of recombinant DNA plasmids and IL28B adjuvant. A gD2 polyclonal antibody produced by our laboratory and anti-HA-tag monoclonal antibody were used in Western blot.
Evaluation of specific immune responses and protection of gD2ΔUL25 vaccine in BALB/c mice
DNA vaccines against HSV-2 are generally weakly immunogenic. A nonadjuvanted gD2 DNA vaccine was reported previously to provide partial protection against genital herpes (8), but it yielded low neutralizing antibody levels. To optimize the immunogenicity of the gD2 DNA vaccine, we developed a fusion gene fragment consisting of ΔUL25 (T cell epitopes) linked to the gD2 sequence in experiment 1. In this part, we compared the specific immune responses and protective efficacy of recombinants pgD2ΔUL25 and pgD2 in BALB/c mice.
ELISA and neutralization analysis
Sera were collected from all mice on day 10 after every immunization in experiment 1, and the gD-specific IgG titers were analyzed by antigen-specific ELISA. The results in Figure 3A showed that immunization with pgD2, pgD2ΔUL25, and FI-HSV-2 clearly elicited gD-specific IgG levels higher than those in the ΔUL25 group. The gD-specific IgG levels elicited by immunization with pgD2 and FI-HSV-2 showed significant differences, while immunization with pgD2ΔUL25 markedly raised the specific IgG titer compared with pgD2 and also lower than FI-HSV-2. Immunization with pΔUL25 also did not induce gD-specific IgG responses, as the titer was close to those of sham-immunized controls.
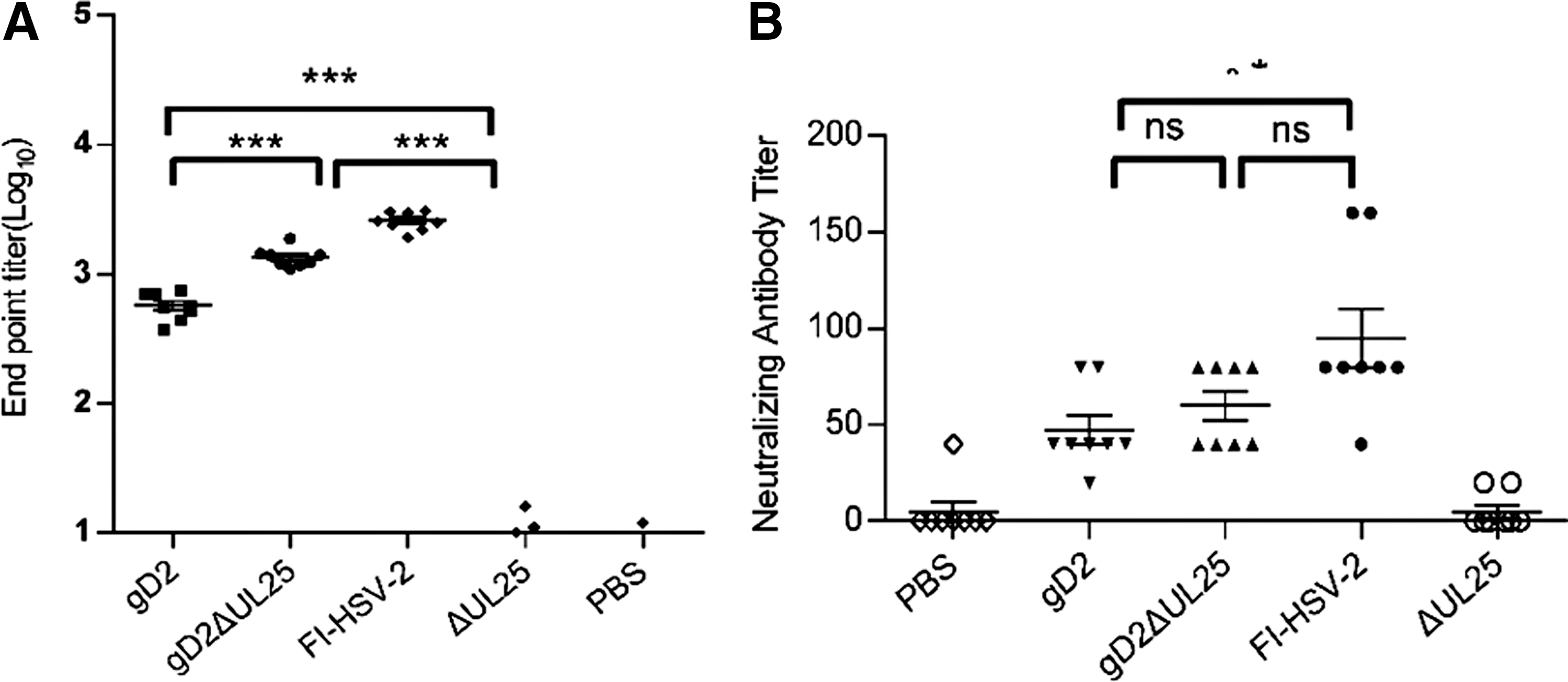
Humoral responses in mice immunized with pgD2ΔUL25, pgD2, pΔUL25, FI-HSV-2, or PBS.
Antibody-mediated neutralization of viral infectivity is reflected by antibody titers measured in neutralization assays. The results in Figure 3B showed that pgD2, pgD2ΔUL25, and FI-HSV-2 vaccines induced high neutralizing antibody titers compared with phosphate-buffered saline (PBS), while the pΔUL25 vaccine presented negative results. The neutralizing antibody titer of pgD2 vaccine (M = 60) was shown to be lower than the FI-HSV-2 vaccine (Fig. 3B). The neutralizing antibody titer of pgD2ΔUL25 did not show significant differences with the FI-HSV-2 vaccine. Compared with sham-immunized controls, mice immunized with pgD2, pgD2ΔUL25 generated a notable level of antibody neutralization, and the neutralizing capacity of pgD2ΔUL25 was similar to the FI-HSV-2 vaccine (Fig. 3B).
The results indicated that the ability of pgD2ΔUL25 vaccine to induce pgD2-specific IgG levels or neutralization titer was greater compared with the pgD2, but lower compared with the FI-HSV-2 vaccine. Meanwhile, immunization with ΔUL25 alone failed to induce gD-specific IgG responses or neutralization.
ELISPOT analysis of IFN-γ responses
IFN-γ production of splenocytes from female BALB/c mice immunized with pgD2, pgD2ΔUL25, and FI-HSV-2 was examined. The HSV-2 gD253-65 (CD4+ T cell epitope) and UL25364-372 (CD8+ T cell epitope) peptides could stimulate T cells to secrete IFN-γ (Fig. 4A, B). In our experiment, all experimental groups demonstrated positive results, while splenocytes from the PBS groups for all tests yielded below 15 SFC/106 cells. By detection of IFN-γ secretion from splenocytes upon HSV-2 gD53-65 stimulation, the pgD2ΔUL25-immunized group produced ∼440 SFC/106cells, which was twice that of the pgD2-immunized group and notably higher compared with the FI-HSV-2-immunized group (Fig. 4A). With UL25364-372 stimulation of splenocytes from the pgD2ΔUL25-immunized group, the number of IFN-γ-secreting cells was 206 SFC/106 cells, and the pgD2 was 19.5 SFC/106 cells, while FI-HSV-2 and ΔUL25 groups were 58 SFC/106 cells and 75 SFC/106 cells, respectively (Fig. 4B). The results showed that the number of SFC was highest in the group immunized with pgD2ΔUL25 upon stimulation with HSV-2 gD53-65 (CD4+ T cell epitope) or UL25364-372 (CD8+ T cell epitope) peptides.
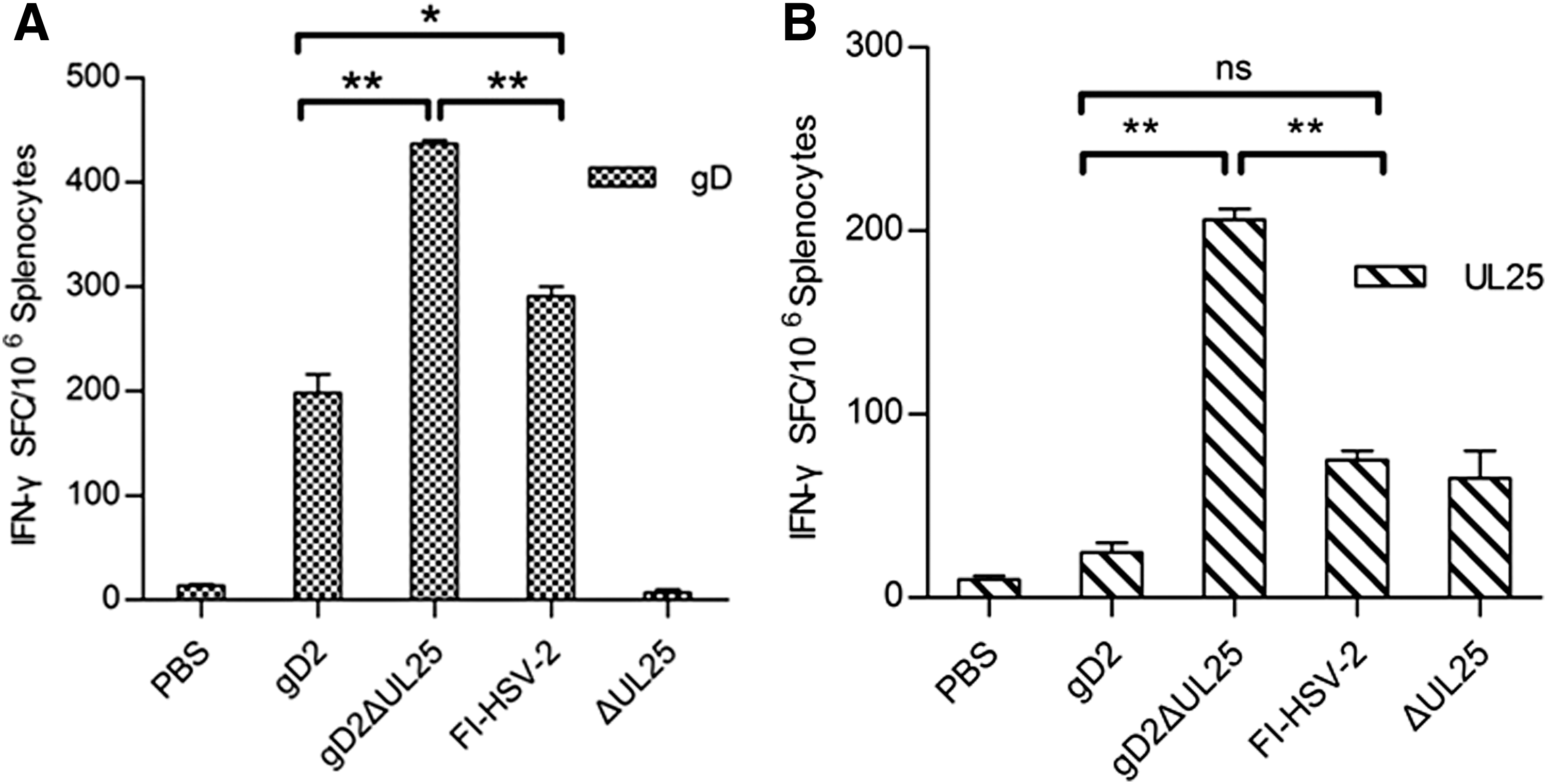
Frequency of IFN-γ-producing splenocytes induced by pgD2ΔUL25, pgD2, pΔUL25, FI-HSV-2, or PBS vaccination in BALB/c mice.
Cytokine multiplex bead assay of Th1-type and Th2-type cytokines
To evaluate the immune profile induced by the various vaccines, we evaluated gD-specific Th1- and Th2-like cellular immune responses by measuring the production of Th1-associated (IL-2 and IFN-γ) and Th2-associated (IL-4 and IL-10) cytokines by using a multiplex bead assay (Fig. 5). Splenocytes from PBS- and pΔUL25-immunized groups produced cytokine levels below 25 pg/mL. As shown in Figure 5A and B, the pgD2- and pgD2ΔUL25-immunized groups both secreted higher levels of Th1 cytokines (IL-2, IFN-γ) than those of the FI-HSV-2-immunized group. As shown in Figure 5C and D, the pgD2- and pgD2ΔUL25-immunized groups produced lower levels of Th2 cytokines (IL-4, IL-10) than those of the FI-HSV-2 group. While the pgD2-and pgDΔUL25-immunized groups produced Th1-biased immune responses, pgD2ΔUL25 notably induced higher secretion levels of IL-2, IFN-γ, IL-4, and IL-10 than pgD2 did (Fig. 5).
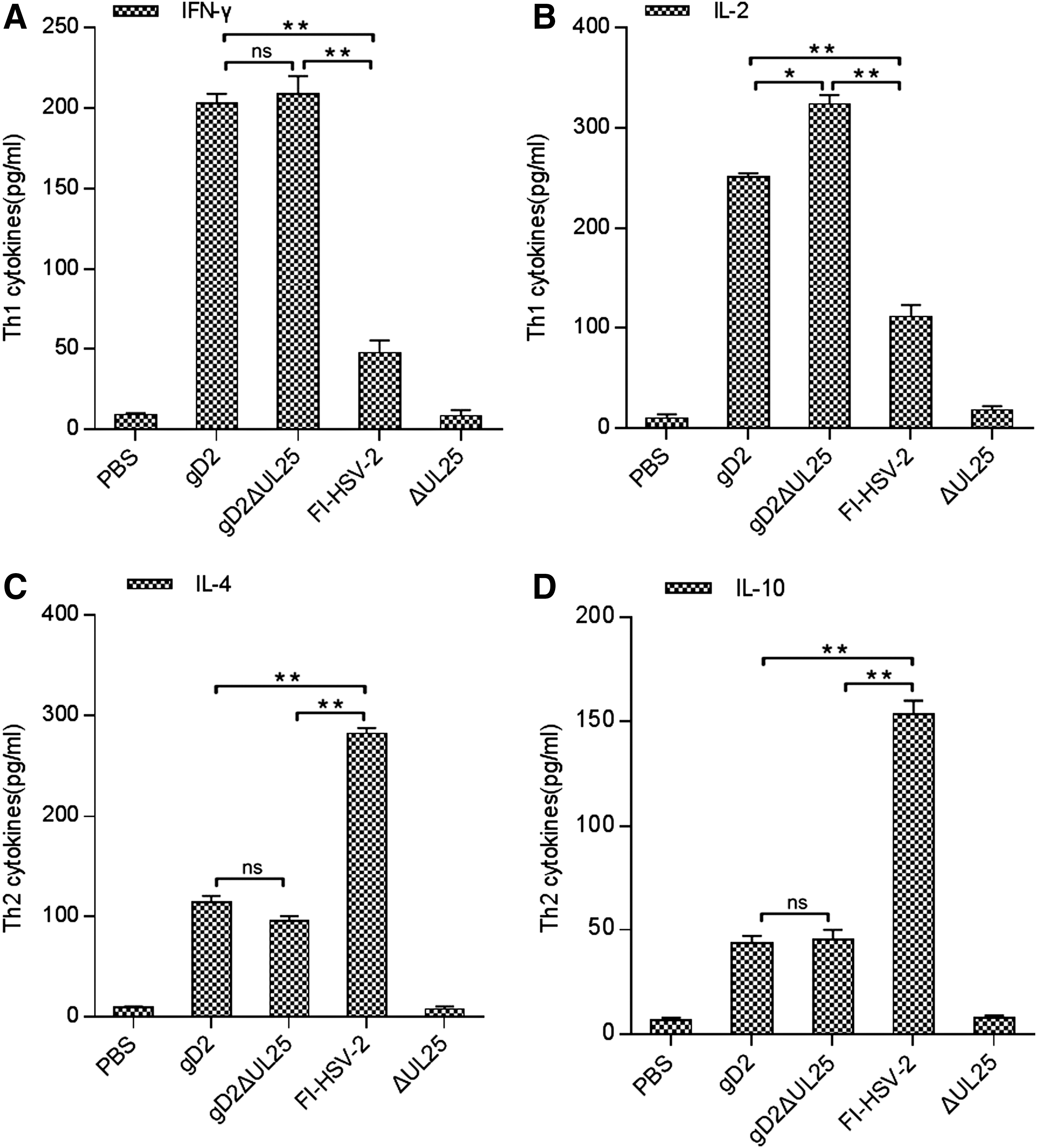
Th1/Th2 cytokine profiles produced by splenocytes from BALB/c mice vaccinated with pgD2ΔUL25, pgD2, pΔUL25, FI-HSV-2, or PBS. Splenocytes were pooled for each test group (n = 5 mice/group) and incubated with gD26–306 at a final concentration of 5 μg/mL.
Protective effects of pgD2 or pgD2ΔUL25 against HSV-2 challenge
Mice were immunized thrice subcutaneously with 100 μg/animal of pgD2 or pgD2ΔUL25 or 12.5 μg/animal of FI-HSV-2. Two weeks after the third immunization, female BALB/c mice were challenged with the HSV-2 strain G at the dose of 1 × 105 PFU/animal.
Virus shedding
Swab samples were taken on day 2, 4, 6, 8, and 10 postchallenge. Vaginal virus shedding that was reflected by the HSV-2 DNA copy number was tested by real-time quantitative PCR (qPCR). PBS-immunized mice did not exhibit any reduction on viral DNA copies, and pΔUL25-immunized group showed a similar trend. By contrast, viruses DNA copies of FI-HSV-2-immunized group and pgD2-immunized group showed 2.9 log and 3.9 log reductions, respectively, on day 10 postchallenge. Moreover, pgD2ΔUL25-immunized group showed a remarkable 4.7 log reduction in HSV-2 DNA copy number down to 103.4 copies on day 10 postchallenge (Fig. 6A).

Survival rates and clinical disease scores in mice immunized with pgD2ΔUL25, pgD2, pΔUL25, or FI-HSV-2 and challenged intravaginally with a highly lethal dose of wild-type HSV-2 strain G. Mice were monitored and scored daily for 20 days postchallenge.
Survival rate after challenge
Animal survival rates postchallenge were monitored for 18 days (Fig. 6B). The survival rate of mice was 37.5% in the pgD2-immunized group and 50% in the pgD2ΔUL25-immunized group, as well as the FI-HSV-2-immunized group. Meanwhile, no mice survived in the PBS- and pΔUL25-immunized groups.
Protection against genital skin lesions
Disease scores were recorded every day postchallenge (Fig. 6C). Genital skin lesion scores of PBS-immunized group and pΔUL25-immunized group were recorded for only 11 days because all mice had died in these two groups. Mice in other groups developed severe skin lesions. On day 13, the mean scores of pgD2, FI-HSV-2, and pgD2ΔUL25 groups reached the peak values of 4, 3.75, and 3.33, respectively. After that, the mean scores showed downward trends. On day 19, the mean scores of the pgD2 group and FI-HSV-2 group decreased to 2.25 and 2.5, respectively. The pgD2ΔUL25 group showed the earliest decline trend on day 14, and its mean score had declined to 2 on day 18. However, no significant difference was observed between the pgD2, FI-HSV-2, and pgD2ΔUL25 groups based on analysis of genital skin lesions. These results indicated that both pgD2ΔUL25 and pgD2 could provide protection against HSV-2 postchallenge in female BALB/c, but without significant differences between them.
Together, results of experiment 1 indicated that immunization with pΔUL25 alone yielded negative results similar to sham-immunized controls, while the pgD2-, pgD2ΔUL25-, and FI-HSV-2-immunized groups showed boosts in humoral and cellular immunity compared with sham-immunized controls. Among these groups, the neutralizing capacity of pgD2ΔUL25 did not show significant differences with the FI-HSV-2 vaccine. The pgD2ΔUL25 group also generated balanced Th1- and Th2-gD-specific T cell responses. However, in the evaluation of protection against HSV-2 postchallenge, neither pgD2 nor pgD2ΔUL25 showed a notable enhancement compared with the FI-HSV-2-immunized group. However, pgD2ΔUL25 showed the higher gD-specific IgG titers, producing a higher level of neutralization, and elicited a satisfactory level of IFN-γ-producing splenocytes compared with pgD2.
Enhancement of pgD2ΔUL25 vaccine effects by IL28B adjuvant
From results of experiment 1, we determined that compared to pgD2, pgD2ΔUL25 more effectively induced gD-specific IgG titers and capacity of neutralizing antibodies, as well as remarkably raised the cellular immunity. However, improving the protection by pgD2ΔUL25 against HSV-2 postchallenge still required more investigation. In experiment 2, the effect of pIL28B as an adjuvant with DNA vaccine pgD2DUL25 was examined.
ELISPOT analysis of IFN-γ responses
IL-28B is a member of the IFN-γ family, which has been identified with antiviral properties similar to IFN-α and IFN-β. IL28B treatment has been reported to enhance HSV-induced expression of IFN-γ, a cytokine known to participate in the response against this virus. To determine if IL28B could enhanced cellular and humoral immune responses in this study, we examined IFN-γ production from splenocytes of female BALB/c mice in all immunization groups [35]. Both HSV-2 gD253-65 (CD4+ T cell epitope) and UL25364-372 (CD8+ T cell epitope) peptides stimulated T cells to secrete IFN-γ (Fig. 7A, B). Splenocytes separated from pIL28B- and PBS-immunized groups for all tests yielded below 15 SFC/106 cells. Under gD253-65 stimulation, the pgD2ΔUL25-/pIL28B-immunized group produced ∼670 SFC/106 cells, which was 1.5 times higher compared with the pgD2ΔUL25-immunized group (456 SFC/106 cells) and 2.1 times the FI-HSV-2-immunized group (311 SFC/106 cells) (Fig. 7A). Under UL25364-372 stimulation, the pgD2ΔUL25-immunized group had 206 SFC/106cells, while the FI-HSV-2-immunized group showed only 88 SFC/106 cells. The pgD2ΔUL25-/IL28B-immunized group produced up to 584 SFC/106cells (Fig. 7B). According to these results, the number of SFC obtained by immunization with pgD2ΔUL25/pIL28B was significantly higher compared with pgD2ΔUL25 alone and FI-HSV-2, both under stimulation with HSV-2 gD253-65 (CD4+ T cell epitope) and UL25364-372 (CD8+ T cell epitope) peptides. These findings indicated that IL28B satisfactorily enhanced the antigen-specific IFN-γ secretion from splenocytes of DNA-immunized mice as expected.
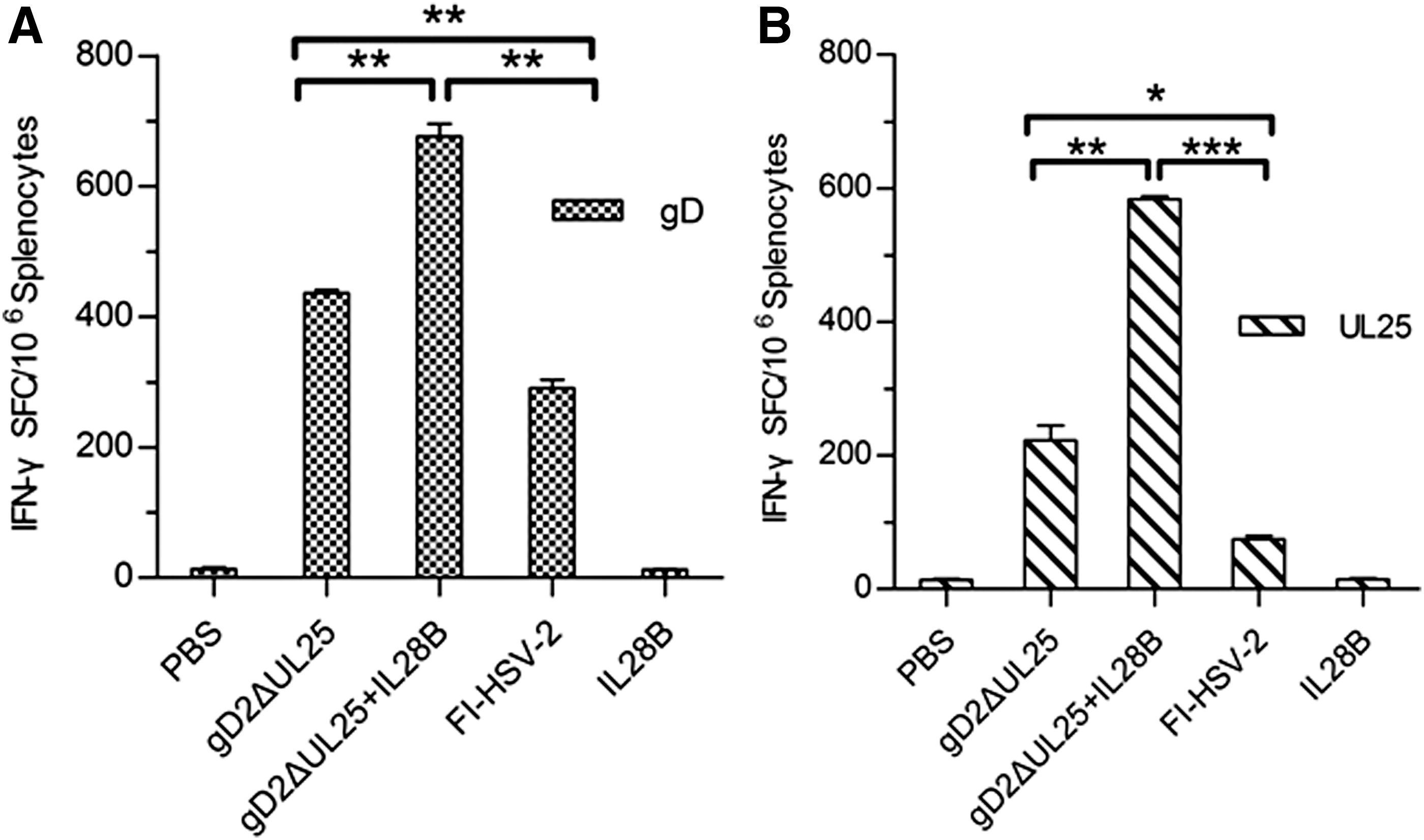
Frequency of IFN-γ-producing splenocytes induced by pgD2ΔUL25, pgD2ΔUL25/IL28B, pIL28B, FI-HSV-2, or PBS vaccination in BALB/c mice.
Cytokine multiplex bead assay of Th1-type and Th2-type cytokines
We next evaluated gD-specific cellular immune responses by measuring the production of Th1-associated (IL-2, IFN-γ) and Th2-associated (IL-4, IL-10) cytokines using a multiplex bead assay (Fig. 8). Levels of cytokines tested in the PBS and IL28B groups were all below 20 pg/mL. The FI-HSV-2 group showed Th2-biased immune responses, with Th2 cytokines levels higher than pgDΔUL25 and pgDΔUL25/IL28B (Fig. 8A, B) and Th1 cytokines levels lower than those of the pgDΔUL25 and pgDΔUL25/IL28B group (Fig. 8C, D). While both pgDΔUL25 and pgDΔUL25/IL28B DNA induced high Th1 cytokine levels, pgDΔUL25/pIL28B induced significantly greater gD-specific levels of IL-2 and IFN-γ. Also, the pgDΔUL25/IL28B group induced higher Th2 cytokine levels than the pgDΔUL25 group, but a little lower than the FI-HSV-2 group. Therefore, following immunization with gDΔUL25 combined with the IL28B adjuvant, T cell-mediated immune responses and humoral responses both upregulated, especially significant in T cell-mediated immune responses.
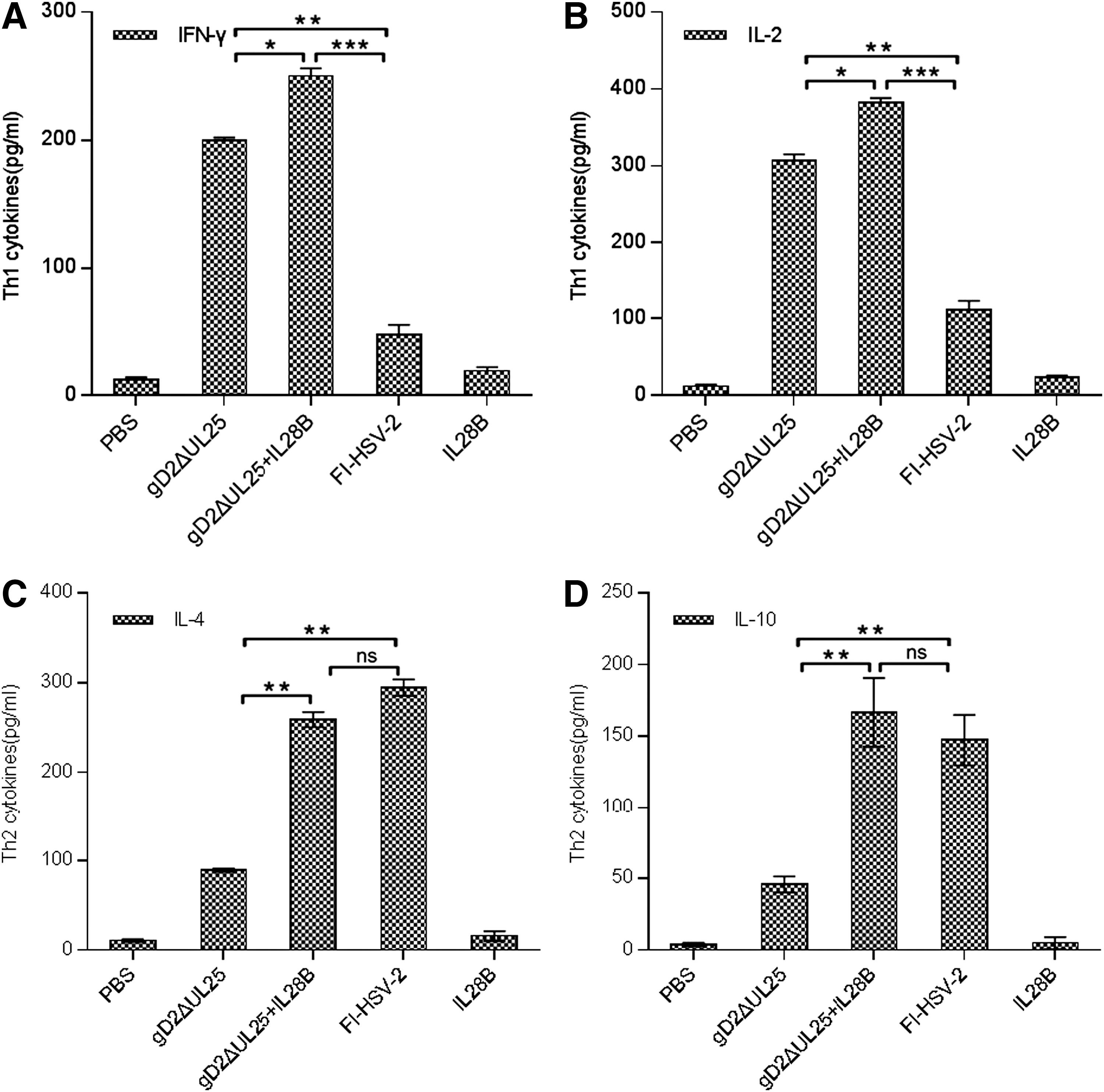
Th1/Th2 cytokine profiles produced by splenocytes from BALB/c mice vaccinated with pgD2ΔUL25, pgD2ΔUL25/IL28B, pIL28B, FI-HSV-2, or PBS. Splenocytes were pooled for each test group (n = 5 mice/group) and incubated with the peptide gD26–306 at a final concentration of 5 μg/mL.
ELISA and neutralization analysis
We also measured gD-specific IgG endpoint titers by ELISA (Fig. 9A). The pIL28B group failed to elicit gD-specific IgG; all the other experimental groups, pgD2ΔUL25, pgD2ΔUL25/pIL28B, and FI-HSV-2, exhibited high gD-specific IgG levels. gD-specific IgG of pgD2ΔUL25/pIL28B was clearly higher than pgD2ΔUL25 group, but showed no significant difference with FI-HSV-2.
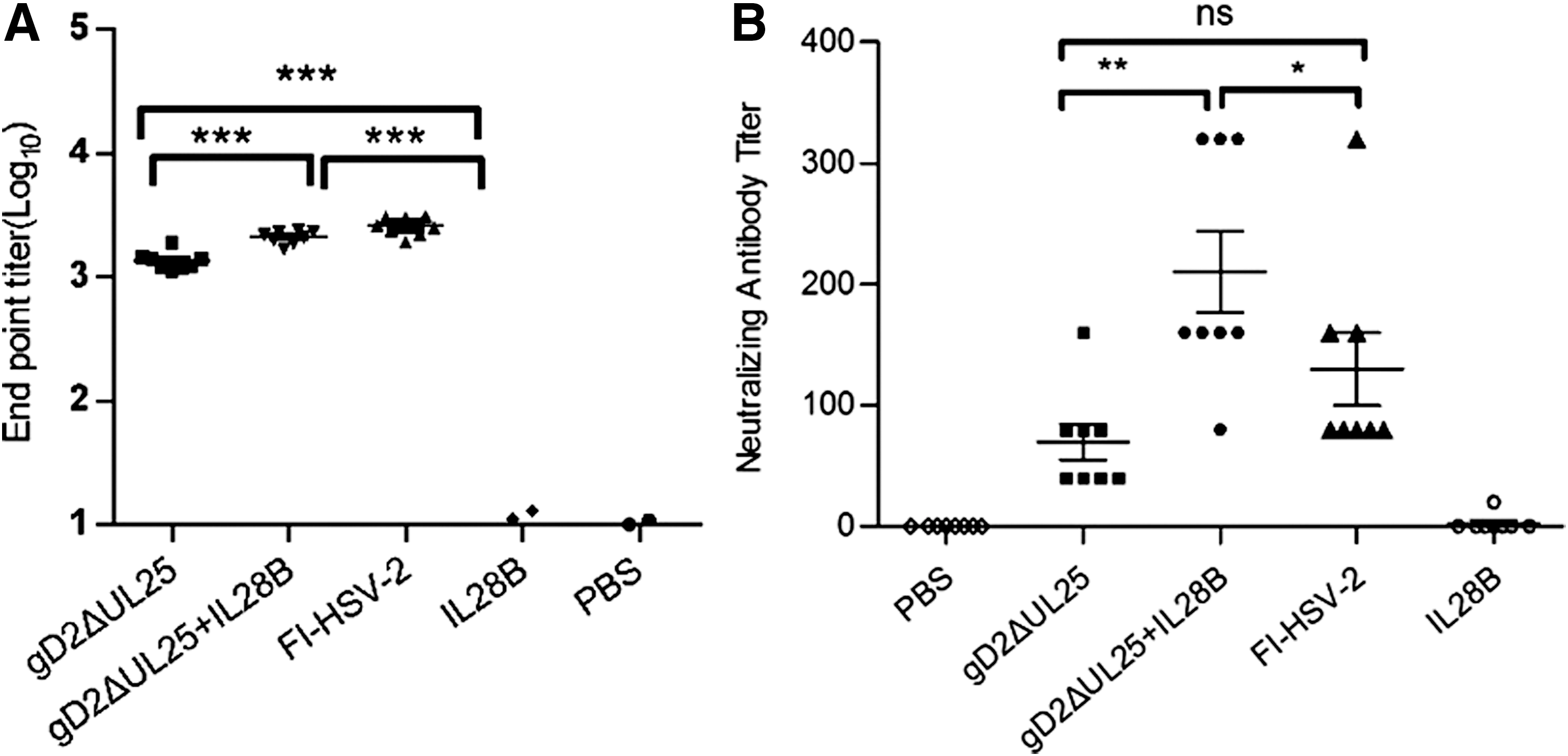
Humoral responses in mice immunized with pgD2ΔUL25, pgD2ΔUL25/IL28B, pIL28B, FI-HSV-2, or PBS.
Antibody-mediated neutralization of viral infectivity by serum samples was measured in vitro and represented by neutralizing antibody titers. As shown in Figure 9B, the neutralizing antibody titer of the FI-HSV-2-immunized group was 130, while that of the pIL28B and PBS groups was ∼0. The neutralizing antibody titer of the pgD2ΔUL25/pIL28B group (M = 190) was thrice higher than the pgD2ΔUL25 group (M = 60) (Fig. 9B).
The results showed that immunization with pgD2ΔUL25/pIL28B led to higher levels of gD-specific IgG and neutralization than with pgD2ΔUL25 alone; meanwhile, results of pIL28B alone were nearly like the PBS group. These findings indicated that IL28B, as a molecular adjuvant, could be applied effectively with the pgD2ΔUL25 DNA vaccine to enhance humoral immunity.
Protection of pgD2ΔUL25/pIL28B against HSV-2 challenge
Mice were immunized thrice intramuscularly with pgD2ΔUL25 (100 ng/animal), pgD2ΔUL25/pIL28B (100 ng/animal), or pIL28B (10 ng/animal), or thrice subcutaneously with FI-HSV-2 (12.5 μg/animal). Two weeks after the third immunization, female BALB/c mice were challenged with the HSV-2 G strain at the dose of 1 × 105 PFU/animal.
Virus shedding
HSV-2 DNA copies of FI-HSV-2-immunized mice were reduced to 104 on day 10 postchallenge, while the pIL28B- and PBS-immunized groups did not show downward trends. The viral DNA copies of the pgD2ΔUL25-/pIL28B-immunized group were reduced to 103.6 copies, and this reduction was more significant than that in FI-HSV-2-immunized mice (Fig. 10A). Compared with pgD2ΔUL25 immunization alone, the pgD2ΔUL25/pIL28B resulted in a slightly, but not significantly effective clearance of virus.
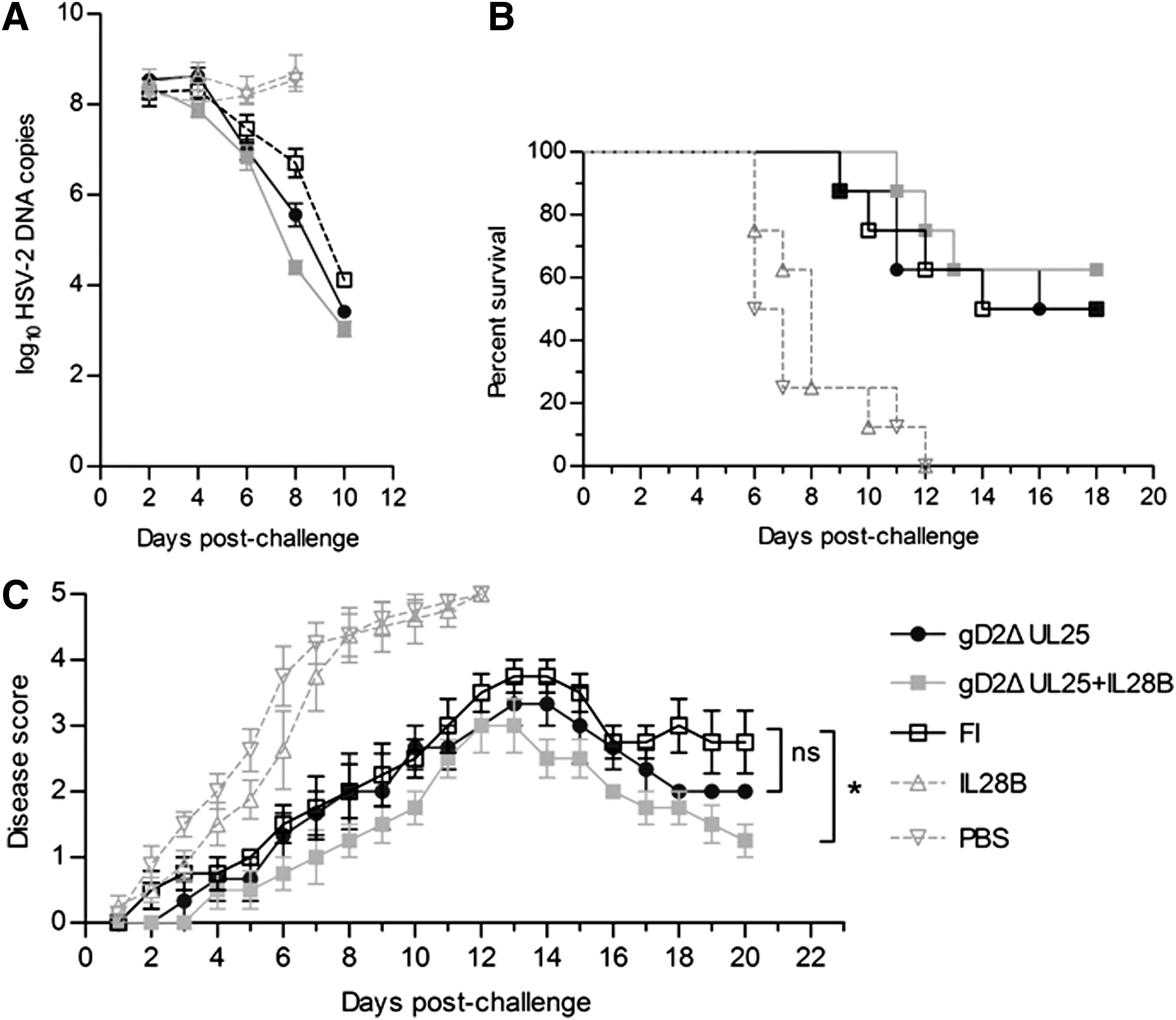
Survival rates and clinical disease scores in mice immunized with pgD2ΔUL25, pgD2ΔUL25/IL28B, pIL28B, FI-HSV-2, or PBS and challenged intravaginally with a highly lethal dose of wild-type HSV-2 strain G. Mice were monitored and scored daily for 20 days postchallenge.
Survival rates after challenge
Survival rates were monitored for 18 days after challenge (Fig. 10B). On day 18, 50% of mice survived in the FI-HSV-2 group, while mice in PBS- and IL28B-immunized groups were all dead. Meanwhile, the survival rate was 50% in the pgD2ΔUL25-immunized group and 62.5% in the pgD2ΔUL25pIL28B-immunized group. pgD2ΔUL25pIL28B-immunized group represented the highest survival rate.
Protection against genital skin lesions
Disease scores were recorded every day postchallenge (Fig. 10C). Mice in PBS-and IL28B-immunized groups developed severe skin lesions with the mean score of 4.375 on day 8, while those of other groups were all under 2. Genital skin lesion scores in the PBS-and pIL28B-immunized groups were only recorded for 12 days due to the death of all mice in the two groups. Skin lesions of pgD2ΔUL25-/pIL28B-immunized group showed the lowest peak with the mean score of 3 on day 12. The FI-HSV-2 and pgD2ΔUL25 groups reached peak mean scores of 3.75 and 3.33, respectively. On day 13 and after that, FI-HSV-2 and pgD2ΔUL25 groups showed downward trends. The mean score of the pgD2ΔUL25 group declined to 2 on day 18, while that of the pgD2ΔUL25/pIL28B group declined to 1.75 on day 18 and to 1.25 on day 20. The score of the pgD2ΔUL25/pIL28B group showed a significant difference when compared with the FI-HSV-2 group, but not with the pgD2ΔUL25 group. These results indicated that the IL28B adjuvant showed tendencies toward improved protection against HSV-2 challenge as determined by virus shedding, survival and clinical symptoms, but these trends did not reach statistical significance.
Discussion
Some previous studies have indicated that cellular immune responses play important roles in the control of infection and transmission (4). During the past few years, recombinant live-attenuated, replication-deficient viruses and viral-vectored vaccines have been extensively developed as vaccine platforms (3,20,21). Currently, research is focused on some DNA vaccines with T cell epitopes in the fields of intracellular infectious diseases and tumor biotherapy (23), and the DNA vaccines might provide critical clues in protein vaccine design or in prime and boost strategy. DNA vaccines, which can induce systemic T cell responses, including polyfunctional cytokine-secreting CD4+ and CD8+ T cells, were chosen as a platform in this study.
We first designed recombinant DNA vaccines pgD2, pgD2ΔUL25, and pΔUL25 and compared their immunogenicities, and a subcutaneous immunized FI-HSV2 vaccine was used as positive control in experiment 1. pΔUL25 yielded positive results when detecting IFN-γ-secreting splenocytes, indicating that it expressed T cell epitopes, which could elicit antigen-specific CD8+ T cell responses. The fusion vaccine pgD2ΔUL25 led to higher specific IgG levels and neutralization titers than pgD2, but lower than FI-HSV-2, and induced greatest number of IFN-γ-secreting splenocytes. The pgD2ΔUL25 vaccine produced Th1-biased immune responses compared with FI-HSV-2, which means that, for cellular immunity, pgD2ΔUL25 was good candidate. In the protection experiments, pgD2 showed the weaker protection or immunogenicities than both pgD2ΔUL25 and FI-HSV-2, while pgD2ΔUL25 showed the same level of protection as subcutaneous immunized FI-HSV-2.
The results in experiment 1 indicated that the DNA fusion vaccine formulation could elicit good immune responses. Also, the pgD2ΔUL25 vaccine induced higher levels of cellular immune responses than pgD2, but the same level of protection against HSV-2. Among various means of improving vaccine efficacy, the adoption of adjuvants to promote immunogenicity of an antigen is one of the most simple, yet most effective ways. As a member of the IFN-λ family, IL28B has been reported to have an antiviral activity against influenza A virus, hepatitis C virus, and HSV (29). IL-28B was shown to affect not only humoral responses but also produce a potent effect on cellular responses by modulating the Th1/Th2 cytokine balance (4).
In experiment 2 of this study, IL28B used as an adjuvant in combination with pgD2ΔUL25 (pgD2ΔUL25/pIL28B) led to a higher gD-specific IgG level and neutralization titer than observed with pgD2ΔUL25 alone, and similar to subcutaneous immunized FI-HSV-2. Meanwhile, immunization with IL28B alone showed negative results. These findings indicate that the IL28B adjuvant could remarkably raise the HSV-2 neutralization capacity of antisera raised by DNA immunization.
In the evaluation of cellular immunity, antigen-specific IFN-γ responses to pgD2ΔUL25/pIL28B were greatest both in CD4+ and CD8+ T cell subsets. Th1/Th2 cytokines secreted by activated immunocytes after immunization play a pivotal role in controlling HSV-2 infection and disease progression in animal models. IL28B was able to enhance the secretion level of all cytokines, both Th1/Th2 cellular responses. These results showed that IL28B as an adjuvant could generate balanced Th1/Th2 cellular responses.
Effects of pgD2ΔUL25/pIL28B vaccine were evaluated by clearance of HSV-2, disease scores recorded every day postchallenge, and survival rate compared to the pgD2ΔUL25 and FI-HSV-2 vaccines. We demonstrated that pgD2ΔUL25/pIL28B could strengthen the protection after challenge compared with pgD2ΔUL25 alone, but these trends did not reach statistical significance. In experiment 1, without IL28B as adjuvant, pgD2 and pgD2ΔUL25 vaccines showed similar or lower results on disease scores, survival rate, and copies of HSV-2 with FI-HSV-2 vaccines. In experiment 2, IL28B as adjuvant recombinant with pgD2ΔUL25, the score of the pgD2ΔUL25/pIL28B group showed a significant difference when compared with the FI-HSV-2 group, but not with the pgD2ΔUL25 group. These findings indicate that the IL28B adjuvant could enhance immune responses and protection against HSV-2 challenge, but using the strategy of DNA vaccine with adjuvant designed as pgD2ΔUL25/pIL28B, we expected more significant immune responses.
The lower than desired humoral and cellular immune responses induced by pgD2ΔUL25/pIL28B may be attributed to several reasons. First, the amount of antigen proteins that can be expressed by DNA vaccines in an animal model is limited. As the DNA vaccines showed weak immunogenicity, as reflected by humoral immune responses in BALB/c mice in this study, the best animal model should be further determined. Second, since IL28B could influence cellular responses by modulating the Th1/Th2 cytokine balance, but did not provide a significant level of protection against HSV-2, the dose and immunization route may need to be optimized. Third, protection after challenge is the result of a complex interaction of local immune factors and responses, and the mechanism of immunomodulation by DNA vaccination is not completely understood. Moreover, one reason that our DNA vaccine was not optimal was that it was administered without a delivery enhancing technology such as electroporation that could enhance the antigen expression at least 100-fold (2,28,35,40). Nevertheless, IL28B was demonstrated to be functional and a promising adjuvant for HSV-2 DNA vaccines in our study. Further exploration of the optimal immunization route, animal model, as well as immune pathways may enhance its effects.
In summary, two DNA vaccines pgD2 and pgD2ΔUL25 were developed and evaluated for immunogenicity in this study, and the pgD2ΔUL25 vaccine was chosen to be optimized with the IL28B adjuvant. Immunization with pgD2ΔUL25/pIL28B induced a high specific IgG level, robust neutralization titer, and a remarkable level of IFN-γ secretion, as well as enhanced Th1/Th2 cellular immune responses. In addition, pgD2ΔUL25/pIL28B significantly decreased HSV-2 viral loads and provided some protective effects against to genital tract lesions generated by HSV-2 compared with controls. The DNA fusion vaccine formulated with the IL28B adjuvant elicited stronger humoral and T cell immune responses than those without the adjuvant. Thus, the findings confirmed our hypothesis that IL28B could enhance the immunogenicity of DNA vectors in BALB/c mice and may be developed as a promising adjuvant for HSV-2 vaccines.
Footnotes
Acknowledgments
This work was supported by Funding: National Natural Science Foundation of China (Grant No. 31200692); The Jilin Province Science and Technology Development Plan (Grant No. 20140309007YY).
Author Disclosure Statement
No competing financial interests exist. Informed consent: No human samples were used in this study.