
Abstract
Aleutian mink disease (AD) is a chronic viral infection that causes autoimmune disorders in minks and presents a significant economic burden on mink farming. Despite the substantial challenges presented by AD, no effective vaccine is available and only partial protection has been achieved. We constructed a whole-gene nucleic acid vaccine from an isolated virulent Aleutian mink disease virus (ADV) strain (pcDNA3.1-ADV). Based on this whole-gene nucleic acid vaccine, we generated truncated mutant constructs by removing portions of the ADV VP2 gene using overlap extension polymerase chain reaction. pcDNA3.1-ADV-428 lacks nucleotides encoding VP2 amino acid residues 428–466, and pcDNA3.1-ADV-428–487 harbors additional deletion of nucleotides coding for VP2 amino acid residues 487–501. We also generated nucleic acid vaccines for the ADV NS1 gene, truncated ADV NS1 gene, ADV VS2 gene, and truncated ADV VS2 gene: pcDNA3.1-NS1, pcDNA3.1-NS1-D, pcDNA3.1-VP2, and pcDNA3.1-VP2-D, respectively. The immunogenicity of the seven DNA vaccines was confirmed by immunofluorescent evaluation. Sixty female minks were divided into 10 groups: seven groups were immunized with the DNA vaccines, one control group was injected with phosphate-buffered saline, one group was immunized with pcDNA3.1 empty vector, and one group was immunized with inactivated ADV-G virus. ADV antibody levels, percentage of CD8+ cells in blood, and levels of γ-globulin and circulating immune complexes in the serum were evaluated longitudinally over 36 weeks after ADV challenge. Minks that were immunized with the pcDNA3.1-ADV-428–487 nucleic acid vaccine produced ADV antibodies. After ADV challenge, the minks immunized with pcDNA3.1-ADV-428–487 nucleic acid vaccine had lower γ-globulin content and lower CIC in serum compared to other immunization groups. Although the pcDNA3.1-ADV-428–487 nucleic acid vaccine did not demonstrate complete protection against ADV, it demonstrated marked efficacy and could potentially be used as a vaccine to prevent losses in mink populations due to ADV. Discovery of effective means to vaccinate mink against ADV will not only improve overall health of mink populations but will also reduce the economic impact of ADV.
Introduction
A
Here we explore the possibility of generating a vaccine against Aleutian disease virus. In this study, we isolated and characterized virulent strains of ADV, and constructed a whole-genome nucleic acid vaccine for a candidate strain, ADV-125. Based on this whole-gene nucleic acid vaccine, we generated truncated mutant constructs by removing portions of the ADV VP2 gene using overlap extension polymerase chain reaction (PCR). pcDNA3.1-ADV-428 lacks nucleotides encoding VP2 amino acid residues 428–466, and pcDNA3.1-ADV-428–487 harbors additional deletion of nucleotides coding for VP2 amino acid residues 487–501. Based on the previous work of the laboratory, six truncated vaccines were generated, including an ADV NS1 gene vaccine, an ADV NS1-D mutant gene vaccine, an ADV VP2 gene vaccine, and an ADV VP2-D mutant gene vaccine. Groups of minks were immunized with the seven different nucleic acid vaccines, and the immunogenicity of these seven vaccines was analyzed.
Materials and Methods
ADV isolation
Kidneys (n = 372) were extracted from minks with suspected AD infection from mink farms in northern China (Heilongjiang, Jilin, Liaoning, and Shandong provinces). Samples were collected beginning in November 2012 and ending in December 2013. Crandel feline kidney (CRFK) cells were used for virus isolation. After three blind passages, cells and cell supernatants underwent three freeze/thaw cycles and were stored at −70°C for later use.
Determination of virulence of isolated virus
Because the virus from the subcultured CRFK cells produced little or no, we used an immunoperoxidase assay to identify isolated virus and to calculate tissue culture infective dose (TCID50). The five AVD strains demonstrating the highest TCID values were selected as candidate strains.
Eighteen female minks that had naive immune status confirmed through convection immunoelectrophoresis were randomly divided into six groups. Individuals in groups one to five received an intraperitoneal inoculation of 3 mL of isolated virus (104.0 TCID50/mL). Individuals in the sixth group served as controls and received an intraperitoneal inoculation of 3 mL of saline solution. All individuals were quarantined after inoculation and temperature, respiration, response to stimuli appetite, and behavior were monitored daily for 60 days. The ADV-DL125 strain was selected for further experimentation due to the obvious morbidity demonstrated by infected mink.
Whole-genome amplification and mutation of isolated virus strains
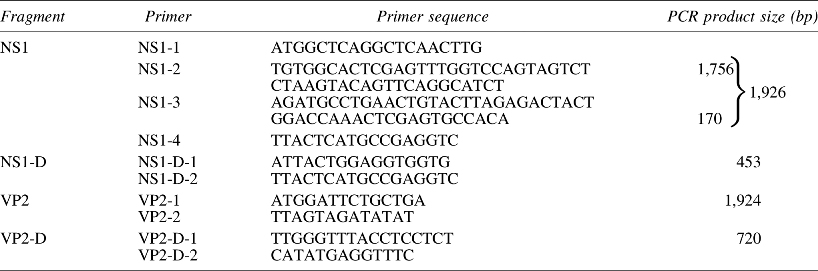
Using Primer6.0 software, we designed a total of 12 pairs of primers to isolate conserved regions of the AD virus, based on published AD viral sequences (GenBank). The primer sequences of each PCR, the location relative to the complete AD virus sequence (ADV-G strain), and primer annealing temperatures are shown in Table 1. The annealing temperatures for the 12 primer sets were 55.6°C, 52.6°C, 51.1°C, 54°C, 51.5°C, 57°C, 54.9°C, 54.2°C, 52.6°C, 56°C, 58.7°C, and 55.7°C, respectively.
The annealing temperatures in the 12 different amplification reactions were 55.6°C, 52.6°C, 51.1°C, 54°C, 51.5°C, 57°C, 54.9°C, 54.2°C, 52.6°C, 56°C, 58.7°C, and 55.7°C, respectively.
PCR, polymerase chain reaction.
PCR amplification products were ligated into the pMD18-T carrier vector and positive clones were sent to Sangon for sequencing. Sequencing results showed that specific amino acid residues of the VP2 protein from the ADV-125 strain (residues 352, 395, and 534) were valine, glutamic acid, and aspartic acid, respectively; these are identical to the same residues in the VP2 protein from the ADV-U strain (10). Using overlap extension PCR technique, mutation constructs were generated. The VP2 protein residues 352, 395, and 534 were mutated to isoleucine, histidine, and histidine, respectively; these residues are identical to the residues from the ADV-G strain. Primers used for overlap extension PCR mutation are shown in Table 2.
Mutations in italics.
Construction of seven DNA vaccines
The whole genome of ADV-125 was cloned and inserted into pcDNA3.1 vector by overlap extension PCR to construct the mutant nucleic acid vaccine (pcDNA3.1-ADV). The nucleotide sequences encoding amino acids 428–446 in the ADV VP2 gene were removed from pcDNA3.1-ADV by overlap extension PCR to generate the mutant recombinant plasmid pcDNA3.1-ADV-428. The nucleotide sequences encoding amino acids 487–501 in the ADV VP2 gene were also removed from pcDNA3.1-ADV-428 by overlap extension PCR to construct the whole-gene mutation recombinant plasmid pcDNA3.1-ADV-428–487. Primers used to generate the mutant constructs are shown in Table 3. Based on previous work in our laboratory, four pairs of primers were designed to generate the ADV NS1 full-length gene, ADV NS1 truncated gene, ADV VP2 full-length gene, and ADV VP2 truncated gene vaccines (primers are shown in Table 4). The four segments of the ADV gene truncations were amplified from the whole ADV-125 genome vector pcDNA3.1-AVD. The truncated genes were then ligated into the pcDNA3.1 eukaryotic expression vector to construct the ADV truncated nucleic acid vaccine.
ADV, Aleutian mink disease virus.
Immunofluorescent evaluation of ADV gene products by DNA vaccines
CRFK cells were seeded in a cell culture plate and cultured overnight at 5% CO2 and 37°C. When the cell density reached 80% to 90%, CRFK cells were transfected with recombinant plasmids with the FuGENE® HD Transfection Kit (Code No. E2311; Promega), according to the manufacturer's instructions. Purity of the recombinant plasmids was analyzed by optical density (OD260/280 ratio ranging from 1.7 to 1.9). Plasmids and transfection reagents were transfected into CRFK cells at 1:2. Transfected CRFK cells were cultured at 37°C and 5% CO2. Transfection control groups include transfection with empty plasmid and mock transfection groups. Expression products were detected by routine indirect immunofluorescence: transfected cells were fixed in ice-cold acetone and incubated for 45 min at 37°C with ADV-positive serum. Cells were washed with phosphate-buffered saline (PBS) and incubated for 45 min at 37°C with fluorescein isothiocyanate (FITC)-labeled rabbit anti-mink IgG antibodies (Code No. bs-0777R-FITC; Bioss Antibodies, China) diluted 1:100 in the above solution. Cells were washed again with PBS and mounted in a microscope slide using 5 μL of antibleaching DABCO solution (1,4-diazobicyclo-[2,2,2]-octane, dissolved at 100 mg/mL in 90% glycerol in 1× PBS).
Vaccination protocol
Seven recombinant plasmids were extracted and purified using the EndoFree plasmid Giga kit (QIAGEN, Germany), following the manufacturer's instructions. Vaccination grade plasmid DNA was obtained by the following adjustments to the protocol: plasmids were resuspended overnight at 4°C in endotoxin-free water, precipitated, washed with 70% ethanol, and finally resuspended overnight at 4°C in endotoxin-free PBS. Plasmid concentrations were adjusted to 1 mg/mL using endotoxin-free PBS and stored at −20°C.
Sixty 6-month-old female minks that were confirmed to be AVD naive were included in this vaccination study. Sixty minks were randomly divided into 10 groups (n = 6). Animals were allowed to adapt to their environment for 15 days. Minks were injected three times (21-day intervals) with 800 μg of plasmid before virus challenge. The specific immunization groups are shown in Table 5 (6). Seven groups of minks received vaccine immunization, one group received PBS control injections, one group received pcDNA3.1 empty vector control injections, and one group received injections of inactivated ADV virus.
PBS, phosphate-buffered saline.
One month after the end of the immunization protocol, the minks were challenged with 105 ID50 of type 4 ADV (9). Immediately before each vaccination, mink were anesthetized with 0.1 mL xylazine 20 mg/mL (Narcoxyl Vet., A/S Rosco, Taastrup, Denmark) and 0.6 mL ketamine 50 mg/mL (Ketaminol Vet., Intervet Scandinavia AS, Skovlunde, Denmark).
The study protocol was reviewed and approved by the Ethics Committee of the Jilin Institute of Endemic Disease Control and Prevention. All the minks were treated in strict accordance with the Guidelines for the Use and Care of Laboratory Animals from the Chinese CDC and the Rules for the Implementation of Laboratory Animal Medicine (1998) from the Ministry of Health, China. The protocols followed for the use of the animals were approved by the National Institute for Communicable Disease Control and Prevention, China. All surgeries were performed under ether anesthesia, and all efforts were made to minimize suffering.
Flow cytometry
Blood was collected in heparinized capillary tubes. Red blood cell lysis was performed on 250 μL of blood by incubation of blood for 6 min in 11 mL erythrocyte lysing solution (0.15 M NH4Cl, 10 mM KHCO3, 1 mM disodium EDTA, pH 7.3); 2 mL PBS was added after incubation (4). Peripheral blood lymphocytes (PBLs) were centrifuged for 5 min at 800 g and washed once in PBS. Cells were washed in 1% normal mink serum/PBS with 0.02% azide and incubated on ice for 1 h with a mouse monoclonal antibody to human CD8+ lymphocytes that is cross-reactive with mink CD8 (cat. No. CRL 8014; ATCC) (1,11). An FITC-conjugated F(ab′)2 fragment of a rabbit antibody to mouse IgG was used as a secondary antibody (cat. No. 313; DAKO, Glostrup, Denmark) (2).
Plasma electrophoresis
γ-globulin quantitation was performed by agarose electrophoresis (LSA agarose, Litex, Copenhagen), using the Tris-barbital buffer system adjusted to pH 7.5. Serum (0.2 mL) was added with 0.02 mL amido-black staining, 37°C water bath for 30 min. The rectangular hole was placed at the end of the agarose gel, and the stained serum was added to the hole. Four layers of filter paper were used for electrophoresis. When the stained serum reached the farthest distance, the protein fractions were quantitated by densitometry (LKB 2202 ultrascan densitometer) (15).
ADV enzyme-linked immunosorbent assay
Enzyme-linked immunosorbent assay (ELISA) plates were coated with ADV. The plates were incubated with increasing dilutions (1:1,000, 1:3,000, 1:9,000, 1:27,000, 1:81,000, and 1:270,000) of mink plasma followed by incubation with 100 ng of biotinylated rabbit IgG to mink IgG, and finally a 1:1,000 dilution of peroxidase/streptavidin conjugate (Code No. P 0397; DakoCytomation, Denmark).
Circulating immune complexes
The final concentration of 5% polyethylene glycol solution was prepared with 1 M borate buffer of pH 8, 4, 0, mixed with the serum to be tested at a 1:30 final dilution, and incubated at 4°C for 1 h, followed by a room temperature incubation (17–20°C) for 30 min. The OD495 value was measured with a microplate reader.
Results
Isolation and determination of virulence of ADV strains
Our immunoperoxidase assay allowed ready identification of ADV-infected cells, as the parts of the cells containing virus stained brown or tan and exhibited distinct cell borders and clear structures when viewed by a microscope (Fig. 1a). Uninfected cells exhibited slight and irregular staining (Fig. 1b). Quantitation of ADV content in cell culture supernatants measured with this method was: 105.7, 105.0, 104.6, 105.2, and 104.1 TCID50/mL.

Results of immunoperoxidase assay showing
ADV infection was confirmed in all individuals that were inoculated with virus by convection immunoelectrophoresis. All infected individuals became emaciated, exhibited loss of fur glossiness, and displayed antifeedant and guzzle. Some individuals exhibited convulsions, spasms, titubation, ataxia, and other neurological symptoms. Infected individuals exhibited enlargement, hemorrhaging of organs (kidneys, liver, spleen, and lymph nodes), and gastrointestinal bleeding. Mink in the control group remained symptom free and in good health and did not develop lesions. These results indicate that the isolated ADV strain was virulent.
Indirect immunofluorescence assay results
CRFK cells were transfected with ADV DNA vaccines. After 48 h, indirect immunofluorescence was performed: serum from Aleutian disease-positive mink was used as the primary antibody, and FITC-labeled rabbit anti-mink serum was used as the secondary antibody. CRFK cells transfected with the DNA vaccine showed fluorescence signal (Fig. 2a), while CRFK cells transfected with the empty pcDNA3.1 vector showed no fluorescence (Fig. 2b). This indicated that the ADV DNA vaccine was able to infect cells and display good reactogenicity.
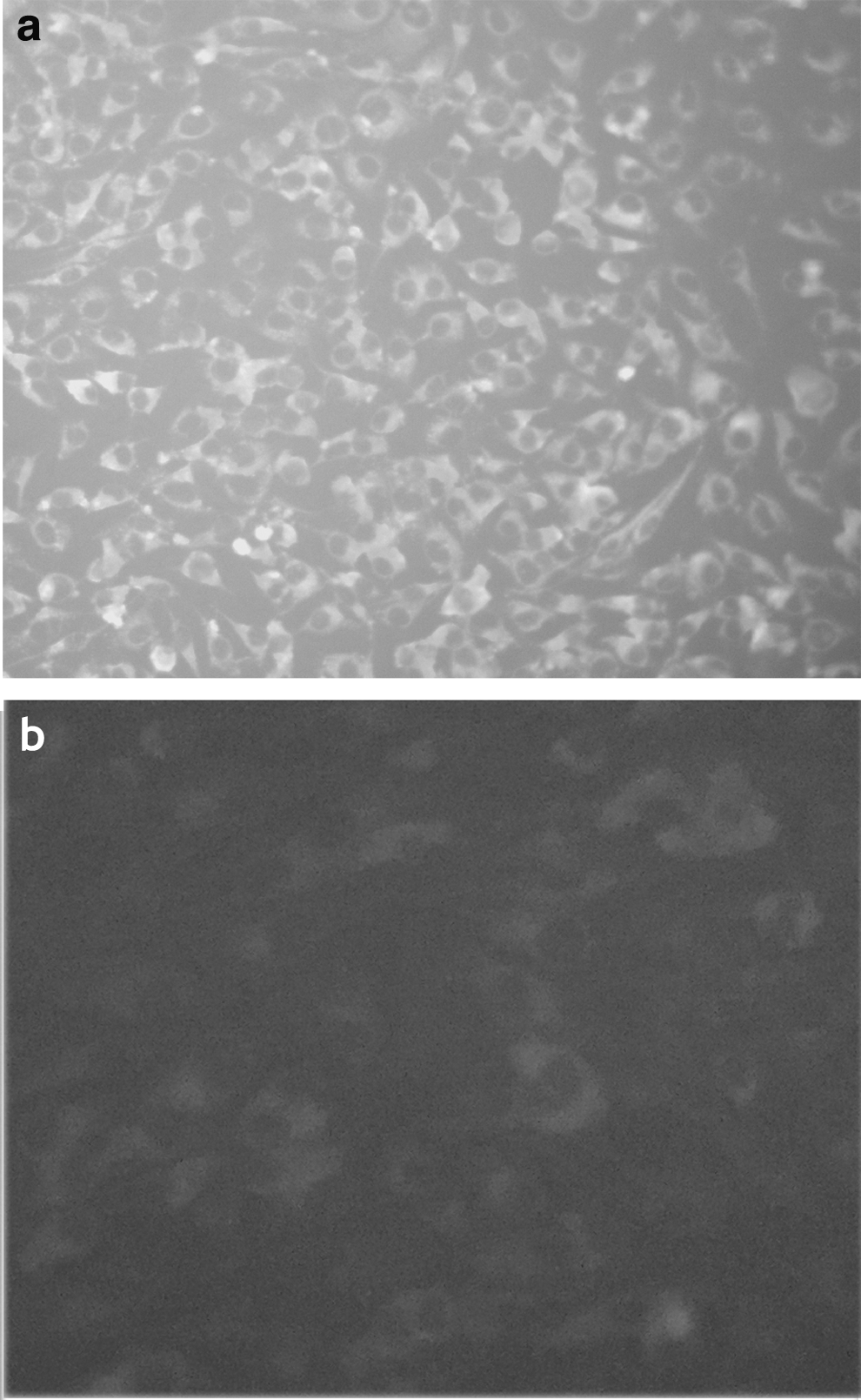
Results of indirect immunofluorescence assay showing
Detection of ADV antibody levels in mink
Following the vaccination protocol, minks were challenged with 105 TCID50 of type 4 ADV. Serum was collected every 4 weeks, and the ADV antibody level in mink serum was detected at each time point (Fig. 3).
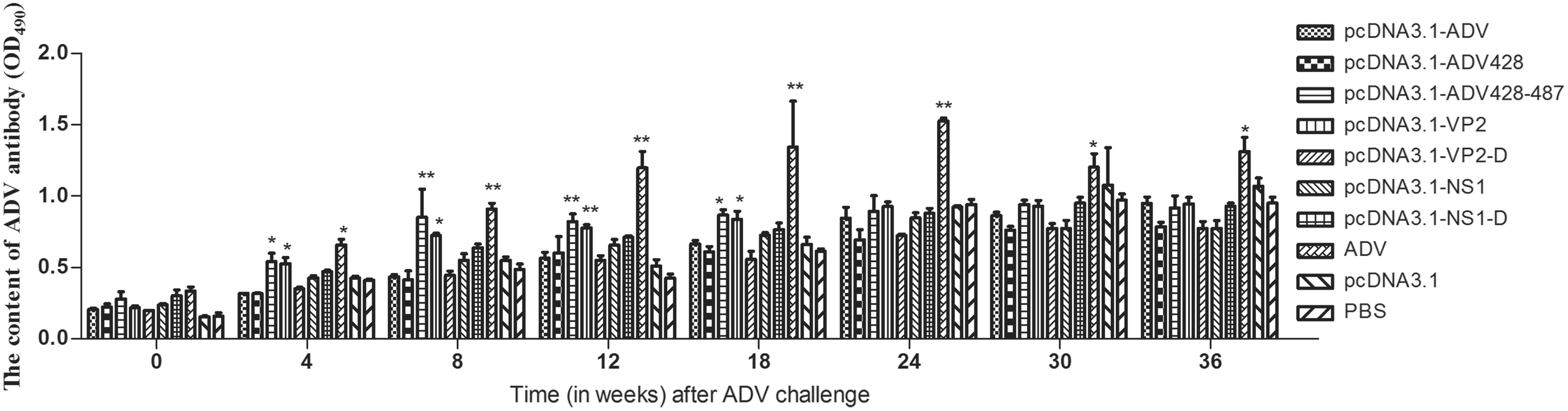
ADV antibody levels in vaccinated mink challenged with ADV. The ADV antibody levels in each vaccination group were evaluated longitudinally after ADV challenge. *Versus the PBS group (*P < 0.05, **P < 0.01). PBS, phosphate-buffered saline.
On the day of ADV challenge, the ADV value in each group was higher than that of pcDNA3.1 empty vector or PBS control groups. The antibody level of pcDNA3.1-NS1-D and ADV-inactivated virus groups was the highest. The ADV content in each group showed an increasing longitudinal trend throughout the experiment. The antibody content of each group was lower than that of ADV-inactivated virus group, and higher than the pcDNA3.1 empty vector or PBS control groups. The ADV antibody content in the mink immunized with pcDNA3.1-VP2, pcDNA3.1-ADV-428–487, and ADV-inactivated virus increased substantially from the fourth week after challenge. Twenty-four weeks after ADV challenge, antibody levels were similar in all the vaccination groups, with the exception of the group immunized with inactivated ADV, which had higher antibody levels.
Detection of circulating immune complexes
Following ADV challenge, mink serum was collected and diluted 30-fold. Serum diluted with 5% polyethylene glycol was used as the detection sample, and serum diluted with PBS was used as control. Levels of circulating immune complexes (CIC) in serum are indicated by subtracting the OD of the control samples from the OD of the test sample (Fig. 4).
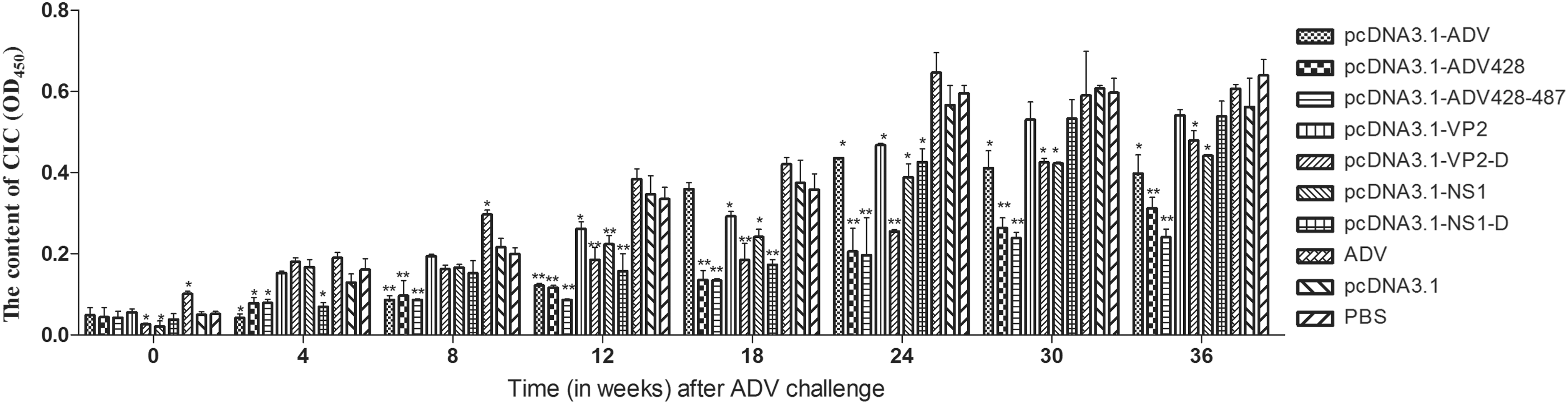
Serum CIC levels in vaccinated mink challenged with ADV. The serum CIC levels in each vaccination group were evaluated longitudinally after ADV challenge. *Versus the PBS group (*P < 0.05, **P < 0.01). CIC, circulating immune complexes.
The serum CIC content in mink serum was lowest on the day of ADV challenge. Over the course of follow-up after ADV challenge, the serum CIC content in the groups immunized with inactivated ADV, pcDNA3.1 empty vector, and PBS control groups increased rapidly; the OD values for these groups were all above 0.3 as early as 12 weeks. At the 18th week, the serum CIC level in the pcDNA3.1-ADV group was substantially higher than in the PBS control group. The serum CIC levels in the pcDNA3.1-ADV-428–487 group were lower than those in the other experimental groups (P < 0.05).
Evaluation of serum γ-globulin
Mink immunized by ADV DNA vaccines and challenged with ADV did not exhibit meaningful variation in serum γ-globulin content compared to control groups (Fig. 5). The average basic percentage of plasma γ-globulin out of total plasma protein from normal mink was found to be 10%. Mink immunized with inactivated ADV displayed increased γ-globulin content in serum over time, and serum γ-globulin in this group was higher than that of the other experimental groups. From the eighth week after ADV challenge, serum γ-globulin content in the pcDNA3.1-NS1 group was substantially increased. At the 12th week after ADV challenge, the serum γ-globulin content in the pcDNA3.1-NS1 group, the pcDNA3.1 empty vector group, and the PBS control group was nearly identical. However, the serum γ-globulin content of pcDNA3.1-ADV-428–487 group was lower by comparison (P < 0.05).
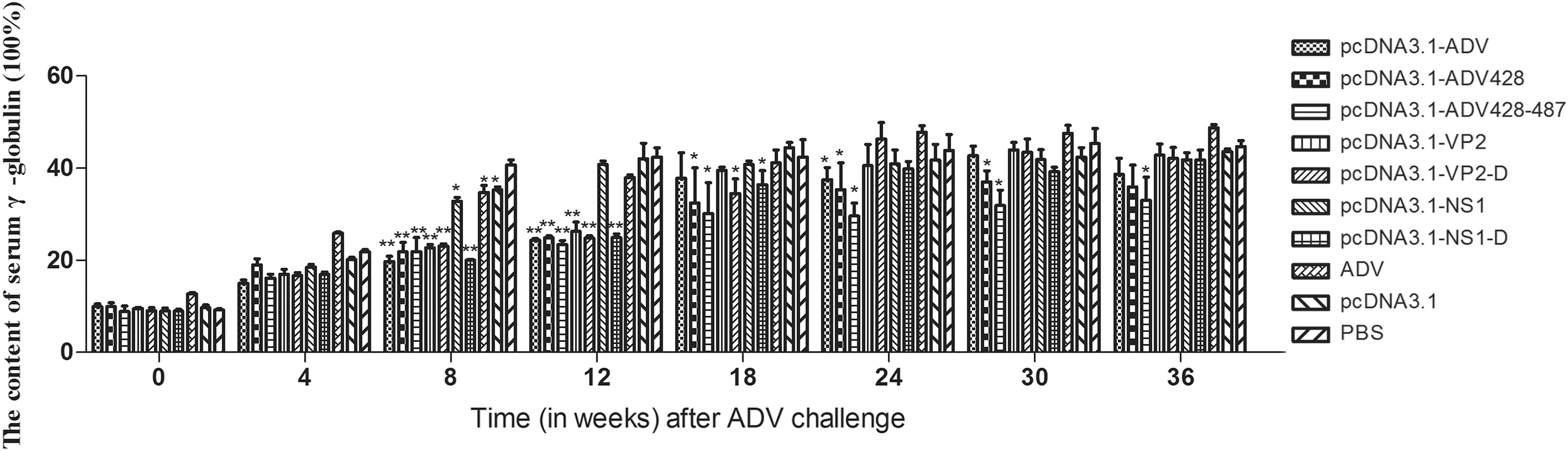
Serum γ-globulin levels in vaccinated mink challenged with ADV. The serum γ-globulin levels in total plasma protein were longitudinally measured in each vaccination group by quantitative plasma protein agarose electrophoresis after ADV challenge. *Versus the PBS group (*P < 0.05, **P < 0.01).
Quantification of CD8+ T lymphocytes
The ADV challenge by itself did not have any effect on the relative quantity of CD8+ cells in the different immunization groups, as shown by flow cytometry analysis of samples from the animals at the day of challenge (Fig. 6). On this day, the level of CD8+ cells out of the lymphocyte population was about 20%. Four weeks after immunization challenge there was a significant increase in CD8+ cells among PBLs. The percentage of CD8+ cells in the PBLs of the inactivated ADV group was substantially higher than that in the pcDNA3.1 empty vector group and the PBS control group. From the 24th week after the challenge, the CD8+ content of each group showed a descending trend.
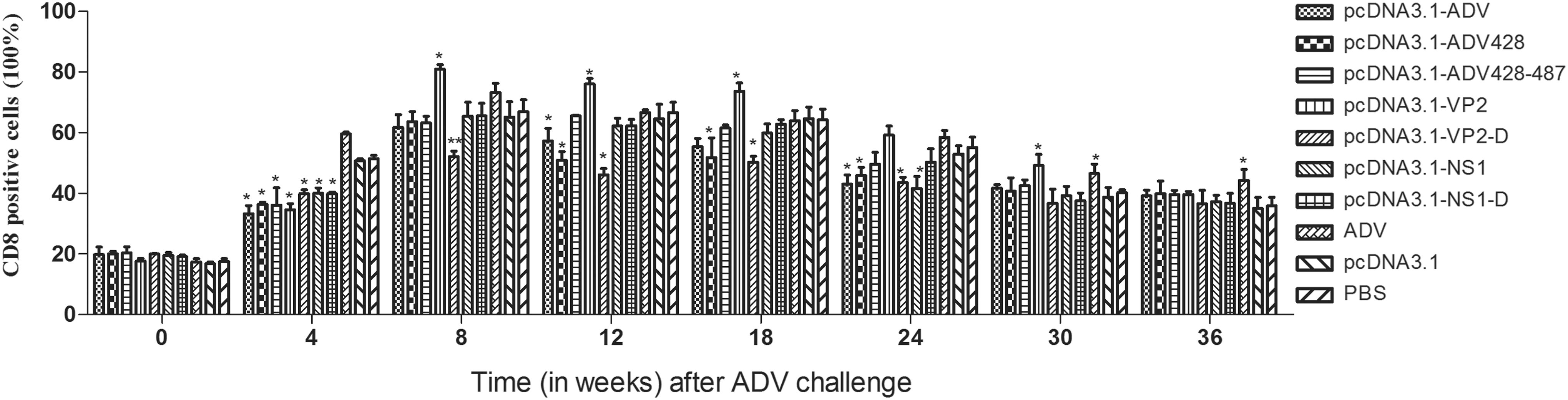
Quantification of CD8+ T lymphocytes in vaccinated mink challenged with ADV. The percentage of CD8+ lymphocytes in each vaccination group was evaluated longitudinally by flow cytometry after ADV challenge. *Versus the PBS group (*P < 0.05, **P < 0.01).
Mortality
Following ADV challenge, animals immunized with inactivated ADV began to die at eight weeks. From the 24th week, mortality was observed in animals from each vaccination group. At the end of 36 weeks, the pcDNA3.1-ADV-428–487 vaccination group showed the lowest mortality (Fig. 7).
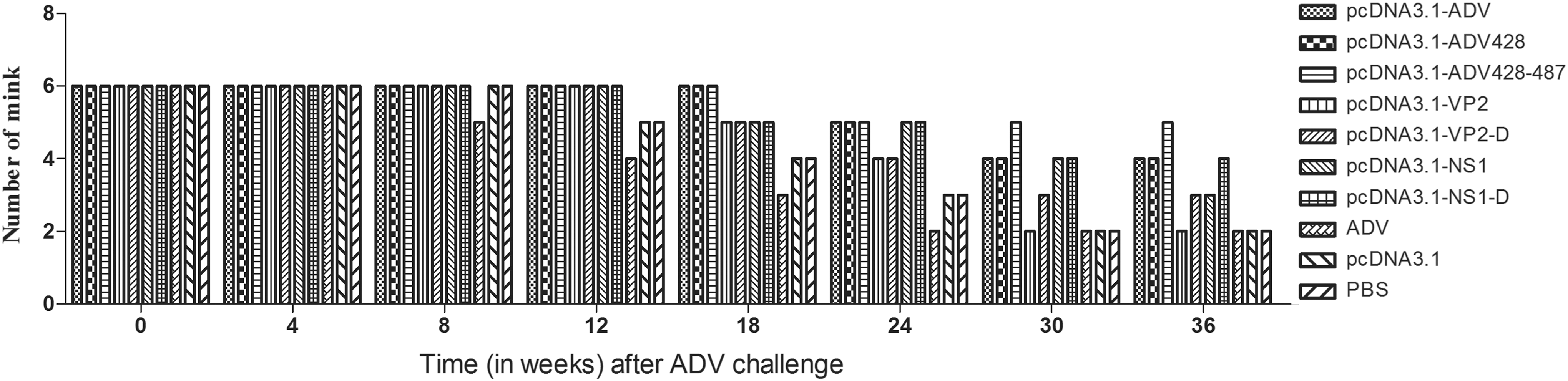
The number of minks after ADV challenge.
Discussion
Minks infected with ADV exhibit a dramatic increase in serum γ-globulin content and increased CD8+ T lymphocytes, which can secrete large amounts of IFN-γ (5). This suggests that the protective effects of an ADV vaccine can be inferred through analysis of serum γ-globulin and CD8+ T lymphocyte populations.
In this study, 4 weeks after immunization challenge, there was a significant increase in CD8+ cells among PBLs, but the experimental group was lower than the control group (P < 0.05). It maybe showed that DNA vaccine offered partial protection. Over the course of follow-up after ADV challenge, the percentage of CD8+ cells in the experimental groups was similar to the PBS group (except pcDNA3.1-ADV-VP2 group). It did not match the result of serum γ-globulin (the two results of the same group should be the same). It maybe showed that mink immunized with a DNA vaccine can exhibit an increase in CD8+ cells following ADV infection, indicating that CD8+ cell quantifications alone should not be used to infer vaccine efficacy (4).
From the result of ADV antibody levels (Fig. 3) and the content of CIC in serum (Fig. 4), it showed that the inactivated ADV vaccine could accelerate the development of ADV. It may be why the inactivated ADV vaccine was highly unlikely to induce CD8 T cell responses, but detected responses are the highest in the ADV-immunized group (Fig. 6).
Following ADV infection, minks demonstrate generation of ADV antibodies and production of CICs. Accumulation of a large amount of CICs can cause inflammatory reactions that include glomerular inflammation, arteritis, and tissue necrosis. Therefore, we evaluated both the serum γ-globulin content and CIC levels in mink after ADV challenge to achieve a more comprehensive assessment of vaccine efficacy. Eighteen weeks after ADV challenge, the serum γ-globulin content was lowest in the group immunized with the pcDNA3.1-ADV-428–487 DNA vaccine, followed by the pcDNA3.1-ADV-428 group. The CIC content in serum of pcDNA3.1-ADV-428–487 group was lower than that of pcDNA3.1-ADV-428 group, but the CIC content in the pcDNA3.1-ADV-428–487 group was still more than 0.2. These results indicate that immunization with the pcDNA3.1-ADV-428–487 DNA vaccine was more protective against ADV challenge than the other DNA vaccines.
A specific region of the VP2 gene is responsible for the replication of the ADV virus (12). In this region, there are only five amino acid differences between nonpathogenic strains and pathogenic strains. Amino acid residues 352 and 534 of the VP2 protein affect ADV replication and pathogenicity, and may be part of the pathogenic determinant (8,14). The ADV-SL3 strain is the only virulent ADV strain that is easily cultured in vitro. The amino acid sequence of the VP2 protein from the ADV-SL3 strain differs from the ADV-G strain at residue 395, and is identical to residue 395 from the ADV-U strain: VP2 AA residue 395 in the ADV-U and ADV-SL3 strains is glutamine, and in the ADV-G strain is histidine. It remains to be determined if this residue is important in determining pathogenicity of ADV. By mutating VP2 residue 352 valine to isoleucine, residue 395 glutamine to histidine, and residue 534 aspartate to histidine, we expected to obtain a more effective ADV DNA vaccine.
Twelve weeks after the ADV challenge, the serum γ-globulin content in minks immunized with the pcDNA3.1-NS1 DNA vaccine was much higher than that of the pcDNA3.1-VP2 immunization group. This suggested that mutation of these three amino acid sites in the VP2 gene may weaken the pathogenicity of ADV virus. Monoclonal and polyclonal antibodies against the 428–446 peptide of the VP2 protein can produce CIC, and epitopes of the 428–446 peptide may exist mainly in ADV-infected mink and can mediate the ADE reaction. The levels of CIC formation from the 487–501 peptide are similar to that of the 428–446 peptide. To prevent the formation of CIC, the pcDNA3.1-ADV-428 construct harbors deletion of the 428–446 peptide, and the pcDNA3.1-ADV-428–487 construct has deletion of both the 428–446 peptide and the 487–501 peptide.
Conclusions
The minks were immunized with the pcDNA3.1-ADV-428–487 nucleic acid vaccine, which resulted in the production of ADV antibodies. Following ADV challenge, the minks immunized with pcDNA3.1-ADV-428–487 nucleic acid vaccine had lower serum γ-globulin content and lower CIC levels than other immunized groups. Although the pcDNA3.1-ADV-428–487 nucleic acid vaccine did not demonstrate complete protection against ADV, it demonstrated marked efficacy and could potentially be used as a vaccine to prevent losses in mink populations due to ADV.
Footnotes
Acknowledgments
This work was supported by a grant from the National Natural Science Foundation of China (No. 31272565). The Program of Economic Animal Disease Prevention and Control in Jilin (No. 20130521023JH).
Author Disclosure Statement
No competing financial interests exist.