
Abstract
The sudden emergence of infectious pathogens such as Zika virus (ZIKV) holds global health concerns. Recent dissemination of ZIKV from Pacific to Americas with an upsurge of congenital anomalies and Guillain Barre Syndrome (GBS) in adults has created an alarming situation. High-throughput studies are in progress to understand ZIKV's mode of pathogenesis and mechanism of immune escape, yet the pathogenesis remains obscure. Mainly ZIKV's envelope (E) protein and nonstructural proteins (mainly NS1 and NS5) manipulate host cell to support viral immune escape by modulation of the interferon pathway and complement antagonism. The development of direct therapeutics for ZIKV infection is required to overcome the rapidly evolving viral threat. Currently, the existing strategies for ZIKV treatment are only supportive. Although, there is no prophylactic or therapeutic vaccine presently available, however, recent efforts have brought up ZIKV vaccines into clinical trial phase 1. This review presents the highlights of recent advances in understanding immune evasion strategies adapted by ZIKV and existing therapies against the virus.
Introduction
Z
ZIKV infection and its strong association with neurologic syndromes, especially fetal microcephaly, have brought together the scientific community for ZIKV research. ZIKV is broadly categorized in Asian and African strains and recent Brazilian outbreak was instigated by the Asian strain. Due to the 95% sequence similarity, African and Asian/Brazilian strains represent a single serotype (36,45). Nevertheless, neurotropic disorders are mainly inclined toward Asian strain outbreaks rather than African strain. Since there is only sporadic epidemiology of ZIKV in Africa, the neurotropic effect of African strain lacks the evidence. In addition, the evolving pathogenicity of Asian strain has increased its infectivity in host (human) and vector (Aedes aegypti), making its transmission and survival relatively favorable (70).
ZIKV shares sequence similarities and pathogenicity with other human flaviviruses such as West Nile Virus (WNV), Dengue Virus (DENV), Japanese Encephalitis Virus (JEV), and Yellow Fever Virus (YFV). However, ZIKV proteins bear a few differences to significantly enhance neurotropic pathology, its harmful effect on fetal brain development and autoimmunity, where antigens of ZIKV invoke the production of autoantibodies (71). As described earlier, flavivirus nonstructural proteins (NS1 and NS5) contribute toward host immune evasion (85). Lately, the evidence of ZIKV E protein sharing amino acid glycosylation sequence with WNV E protein indicates its inclination toward neurotropic pathogenesis (77).
Due to the absence of antiviral therapies against ZIKV, a new quest for therapeutics has begun. Recent in vitro studies have put forward 20 previously approved compounds that may have the potential to reduce ZIKV viral load (11). Currently, there is no approved anti-ZIKV drug and the scientific community is promptly engaged in developing it.
Recent ZIKV Outbreaks
Historically, only sporadic human cases of ZIKV infection were reported (98). ZIKV attained minor clinical attention before its epidemic in Yap Island, Micronesia, during 2007. The Micronesian epidemic infected about 73% of the population from which only 19% patients showed mild and short-lived symptoms (38,58). In 2013, ZIKV outbreak covered 66% population of French Polynesia in correlation to increased cases of Guillain Barre Syndrome (GBS) (23). GBS is an autoimmune neurologic disorder involving polyradiculitis and demyelinization, leading to inflammation of nerve cells, which further induces muscle weakness (51). Subsequently, ZIKV epidemic dispersed from South Pacific to South America and caused a severe outbreak in late 2015. Brazil was a major victim of this ZIKV outbreak infecting 0.5–1.5 million population with exceeded reports of GBS and neonatal microcephaly (20). Phylogenetic analysis of ZIKV strains in Brazil links it with that of Asian strain involved in French Polynesian outbreak (5,39).
These severe outbreaks enforced World Health Organization (WHO) to declare a Public Health Emergency of International Concern (PHEIC) in February 2016 (48). Centers for Disease Control (CDC) officially confirmed the link of ZIKV infection to neurologic abnormalities in newborns and adults in April 2016 (79,94).
Structure and Pathogenesis of ZIKV
Similar to other flaviviruses, ZIKV possesses nonsegmented single-stranded (+ss) RNA genome closely related to DENV, WNV, JEV, and YFV (52). ZIKV genome consists of 10.794 kilobases flanked by 5′ and 3′ noncoding regions (NCR) encoding for a single polyprotein of ∼3500 amino acids. The polyprotein comprises three structural proteins: capsid (C), precursor of membrane (prM), envelope (E), and seven nonstructural proteins: NS1, NS2A, NS2B, NS3, NS4A, NS4B, and NS5 (68). Figure 1 illustrates the ZIKV structure and genome arrangement (57).
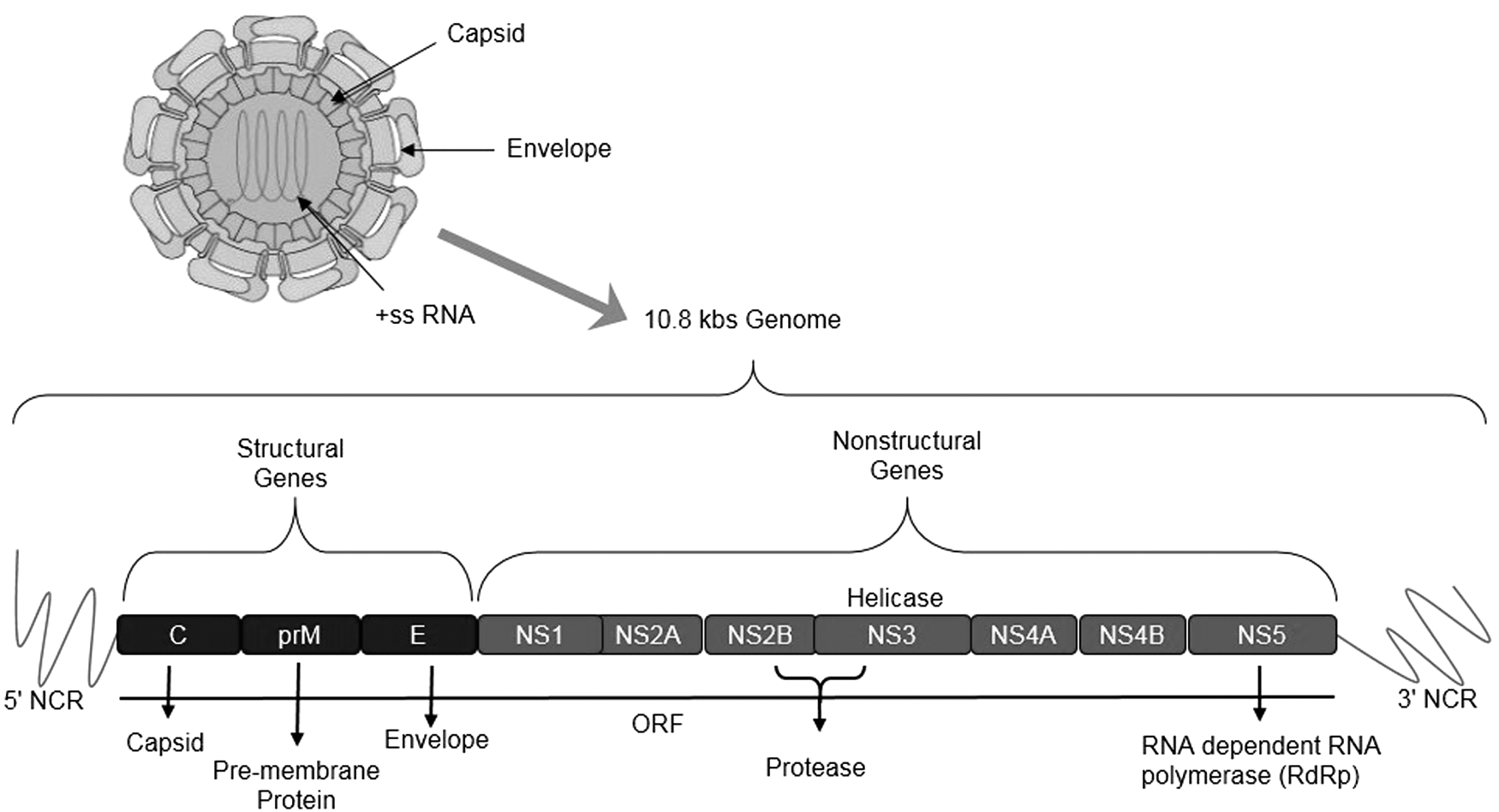
Genome of Zika virus consists of 10.8 kilo bases, which code for three structural proteins, that is, capsid (C), precursor of membrane (prM), envelope (E), and seven nonstructural proteins, that is, NS1, NS2A, NS2B, NS3, NS4A, NS4B, and NS5.
ZIKV infections remain asymptomatic in approximately 80% patients and only 20% express symptoms (38). Generally, symptoms appear from 6 to 11 days after transmission and the virus is cleared from blood within 24 days in 99% of the patients. However, the virus may persist in body secretions for a longer duration, thus avoiding early detection of ZIKV. In the case of pregnancy, the infection extends for a longer period of time due to its replication in the fetal brain for months, and the mother might remain asymptomatic (33,102).
With the previous history of rashes, mild cold-like symptoms such as fever and myalgia, ZIKV was considered a minor threat. Later evidence indicated its linkage with congenital microcephaly and GBS owing to the fact of cross-reactivity of antiviral immune responses against ZIKV with human proteins (24,96,101). The coevolution of Asian strain with respect to its host is one of the factors for neurologic diseases correlating ZIKV infection. An NS1 protein mutation has increased the survival and viremia of ZIKV in A. aegypti and subsequently enhanced pathogenesis in the primary host (human) as well (70).
ZIKV infection during the initial stage of pregnancy caused fetal microcephaly, congenital defects, and miscarriages (105). Immune-compromised mice model of ZIKV infection elucidated the role of ZIKV in neural progenitor cells (NPCs). ZIKV accomplishes NPC apoptosis by inhibiting cell cycle and differentiation leading to cortical reduction and microcephaly (32,64,78).
Immune Evasion by ZIKV
ZIKV RNA genome enters the host cell and starts translation process to encode the single polyprotein molecule. The transcribed polypeptide is then catalytically cleaved into ten distinct proteins, including three structural and seven nonstructural. Flaviviruses have evolved numerous strategies to escape host immunity by inhibiting interferon, complement system, NK cells, and T cell and B cell responses (111). Notably, ZIKV proteins have great similarity to those of other flaviviruses; WNV, DENV, YFV, and JEV. Precursor of membrane (prM), envelope (E), and nonstructural proteins; NS1, NS3, and NS5 play essential roles in ZIKV infectivity and pathogenesis. ZIKV's NS3 and NS5 have been found sequentially and structurally similar to other flaviviruses, but the sequence of prM, E, and NS1 diverges and makes the pathogenesis of ZIKV relatively different (71). Recurrent insights of ZIKV pathogenesis with context to immune evasion mechanisms are limited to NS1 and NS5 role in immune escape, which shows the necessity of further research.
Lately, the comparative computational analysis of E protein from several ZIKV strains of African and Asian lineages gives an indication that amino acid glycosylation upon E protein has a variance among them. The glycosylation of asparagine (ASN) 154 is a significant feature of Asian/Brazilian strains and may potentially contribute toward different pathogenicity (77). ASN 154 N-linked glycosylation is further confirmed by the electron microscopic structural analysis of ZIKV (103). The N-linked glycosylation is a significant feature of WNV association with neurotropism (15). Moreover, the genetic flexibility of E protein to tolerate minor mutations may also play a crucial role in immune evasion (42).
Generally, the multifunctional role of flavivirus NS1 protein is critical in hijacking the host replication system. The cleaved NS1 protein rapidly dimerizes in the intracellular environment and exists as a dimer associated to endoplasmic reticulum (ER) membrane (mNS1). Conversely, in the extracellular environment, it exists as a secreted hexamer molecule (sNS1) (73,74).
Similar to WNV, ZIKV NS1 dimer associates upon the luminal face of ER to lay the base for replication complex, in association with NS4B (a transmembrane protein) and NS4A proteins (93,110). Both NS4A and NS4B interact with hydrophobic β-roll of NS1 as a stable foundation of vesicle packet (VP), a structure induced by nonstructural proteins upon ER membrane, which carries the replication of viral RNA. This phenomenon is yet to be confirmed for ZIKV (112). Within the VP, NS1 also interacts with prM and E proteins for their joining with replication complex in virion development, Figure 2a portrays VP and virion formation (93,100).
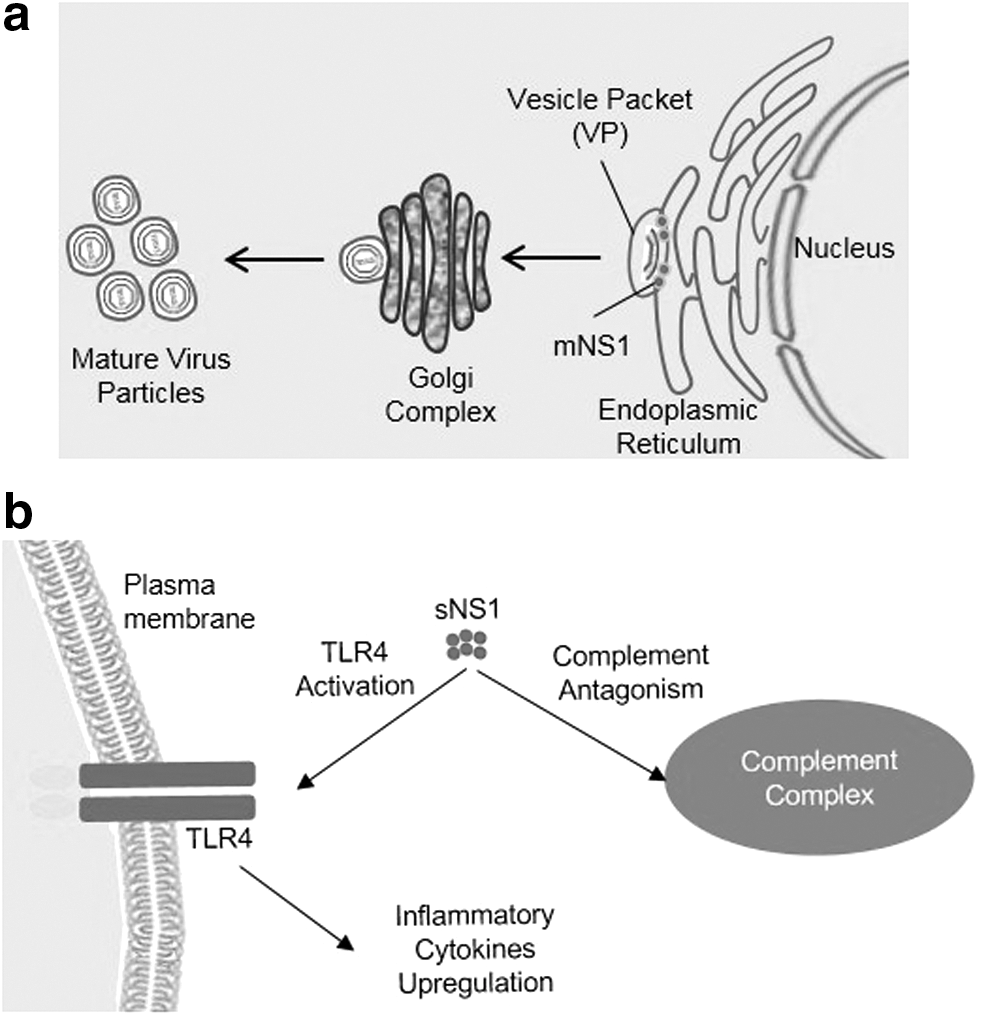
NS1, the only flavivirus protein to be secreted from the cell, plays a variety of roles in immune escape, mainly complement antagonism (28). For the survival of secreted virus progeny, sNS1 interacts with the complement system, which is an antibody-mediated immune response to neutralize the infection (29). sNS1 activates the Toll-like receptor 4 (TLR4) as well, to initiate the inflammation process, which further induces the peripheral blood mononuclear cells and macrophages to express inflammatory cytokines (Fig. 2b) (16). Due to the continuous accumulation of inflammatory cytokines, endothelial cell plasma membrane may lead to vascular leakage (7,80). Furthermore, a single point mutation—alanine to valine—at residue 188 of NS1 protein has enhanced its infectivity and adaptability by increasing the NS1 concentration in host sera (70).
ZIKV proteins intervene with cellular pathway during the developmental stage of the fetus and induce congenital impairment. The Akt-mTOR signaling plays a crucial role in the regulation of cellular development, proliferation, and inhibition of autophagy in NPCs (41). Recent evidence suggests that ZIKV NS4A and NS4B proteins cooperate to inhibit the activation of Akt. Thus, inhibition of Akt-mTOR pathway in NPCs causes an imperfect development of brain leading to microcephaly (67). Preliminary studies indicate that NS4A and NS4B complex inhibit the P13K enzyme, which further terminates the Akt-mTOR signaling cycle; however, this phenomenon requires further explorations.
Type 1 interferon (IFN) pathway is an innate immune response, which establishes an antiviral state in a host cell by induction of JAK-STAT pathway to upregulate interferon-stimulated genes (ISGs) (75,107). ZIKV's NS5 protein elicits a key role in the disruption of ISG stimulation pathway (43). ZIKV attacks STAT2 protein to inhibit the IFN pathway for completion of viral life cycle.
Previously, the nuclear presence of NS5 in DENV and YFV was reported (47,60). Recently, NS5 presence in the nucleus of host cell was indicated, despite its main role for RNA replication in host cell cytosol (43), although the significance of nuclear localization is yet to be discovered. As the consequence of overexpression of human STAT2, ZIKV NS5 reverts to the cytoplasm where it interacts with STAT2 and restrains it from entering the nucleus, as shown in Figure 3 (43). Immune evasion from type I IFN in ZIKV infection is likely to be by STAT2 degradation. Similar is the case of DENV NS5 that degrades STAT2; however, ZIKV NS5-induced degradation is independent of UBR4, which is necessary for DENV (83). Conversely, recent reports indicate the species restriction of ZIKV NS5 from interferon antagonism. In the case of mouse, the type 1 IFN-pathway is required to be completely removed for the propagation of ZIKV infection (6,61,95). In addition, the full-length structure of the NS5 protein of ZIKV suggests a structural inclination toward JEV, which also falls in the neurotropic category (106).

JAK-STAT pathway, an interferon-stimulated pathway for the upregulation of ISGs that are responsible for establishing an antiviral state in the cell; it starts with the stimulation of interferon-alpha receptor 1 and 2 (IFNAR1 and IFNAR2), which activate Janus Kinase 1 (JAK1) and it further activates Signal transducer and activator of transcription 1 (STAT1 and STAT2) for the upregulation of ISGs. Upon successful infection of ZIKV, NS5 protein of ZIKV accumulates in the nucleus of host cell and as the STAT2 concentration increases, it relocates to cytoplasm, where it interacts with STAT2 and thus inhibits the JAK-STAT pathway, leading to immune evasion of ZIKV. ISG, interferon-stimulated gene; ZIKV, Zika virus.
Therapeutic Regimens Against ZIKV
Currently, ZIKV-specific treatments are unavailable, and only supportive drugs are administrated during ZIKV infection to enhance host-immune response (8). The lack of diversity in ZIKV strains makes it easier to find targets for vaccines and antivirals (109). ZIKV was previously overshadowed by DENV and WNV outbreaks. However, the scientific community is in the quest of developing broad-spectrum antivirals and vaccines to control the reemergence of such viral diseases.
Interferon Therapy
IFN administration upregulates ISGs for immune modulation to develop an antiviral state. Emerging evidence suggests that inducible transmembrane proteins (IFITMs) may play a role of restriction factor to inhibit the ZIKV infection. These restriction factors have previously been reported to inhibit a wide range of viruses, including influenza virus, DENV, WNV, Filovirus, and SARS coronavirus (9,21,27,50,104). IFITM3 inhibited ZIKV replication at an early stage assuming that it halts the fusion pore formation by altering the plasma membrane properties (65,92). IFITM3 elicited an intense decrease of ZIKV RNA at early intracellular level. Studies suggest that IFITM3 may provide early defense by viral replication obstruction (99).
IFN therapy for ZIKV infection with context to polyamine depletion causing impairment in viral replication and translation has also been examined. Spermine/spermidine N1- acetyltransferase (SAT1) is a catabolic enzyme for polyamines, which either converts them into putrescine or exports them from cell (91). Escalation in SAT1 concentration increases spermidine and spermine acetylation, making them inaccessible for viral transcription and translation (76). Transcriptomic analysis suggests that type-I IFN induces SAT1 (97) similar to HIV-1 infection of immature dendritic cells (53). IFN-β induced a significant surge in the SAT1 expression, indicating that SAT1 is necessary for IFN signaling. SAT1 knockouts have confirmed an increased viral titer in model studies. A remarkable reduction in ZIKV replication subsequent to SAT1 expression may support its role as a restriction factor. In vitro studies approve that polyamines are prerequisites for virus transcription and translation (84). Thus, SAT1 is a drug candidate to stop the viral replication.
Lately, a mouse model study of an ISG encoding cholesterol-25-hydroxylase (CH25H) enzyme indicates antiviral activity against ZIKV (63). IFN activation by TLR stimulation leads to CH25H production (12,49,90). Mainly, CH25H converts cholesterol into 25-hydroxycholesterol (25HC), which regulates sterol biosynthesis (49,54). 25HC mediates retinoic acid-inducible gene-I (RIG-I) induction in macrophages to upregulate the interleukin-8 (IL-8) pathway, which mediates atherosclerotic inflammation (108). 25HC acts against a number of the virus, including ZIKV, by suppressing viral fusion with host membrane, thus inhibiting the entry of virus particle (19,63,69).
Furthermore, an IFN-mediated protective role against ZIKV was affirmed by IFN-γ release of placental cells to inhibit virus particles in placental barrier (13). IFN is an expensive treatment with severe side effects due to which, it is not recommended during pregnancy in females. Right now, ISG involvement in case of ZIKV inhibition remains unclear.
Nucleoside Analogs
Nucleoside analogs are safer and tolerable in targeting only viral proteins to cause their premature termination. Nucleoside analogs are comparatively efficient drug candidates for the treatment of RNA viruses, for example, sofosbuvir, a drug against HCV infection. C-methylated nucleosides such as ribavirin and favipiravir (T-705) are being tested as possible antivirals for ZIKV (26). C-methylated nucleosides include 7-deaza-2′-C-methyladenosine (7DMA) and 2′-C-methylcytidine (2CMC), which were previously developed against HCV infection.
Murine model testing showed a moderate anti-ZIKV effect of 7DMA, while already approved drugs like Ribavirin and favipiravir were comparatively less effective (40,113). Anti-HCV drug sofosbuvir (approved) has been successfully tested for ZIKV therapy in neuronal stem cells, human tumor cell lines and mice model, with negligible toxic effects (22). The nucleoside analogs BCX4430 and GS-5734 are broad-spectrum antivirals against RNA viruses and are currently in the clinical trials for phase 1 and 2 (86). Broad-spectrum drugs, especially the nucleoside analogs, are possible anti-ZIKV drugs, but it requires more research and evidence to be licensed. These compounds have the potential to be structurally and functionally optimized for their usage as prodrugs. Conclusively, the in vitro ZIKV inhibition activity of 2′-C-methylated nucleosides has shown promising effect.
Anti-ZIKV drugs hold a challenge to cross the blood–brain and placental barrier for their targeting capacity (11). For instance, ribavirin administration during pregnancy is not recommended due to its adverse effects on fetal development. Furthermore, antiviral strategy such as monotherapy against RNA virus diseases may give rise to rapid reemergence of that disease as the virus evolves after a short period of time, thereby indicating the importance of the development of drugs with multitarget approach and its efficient use for a longer time period, as well as reducing the possibility of selecting out the newly emerged strains. Outbreaks like ZIKV infection indicate the importance of broad-spectrum antivirals, for example, ribavirin (31), which can be efficiently taken in the phase III clinical trials to boost the countermeasures against a viral disease (4).
Vaccines
Flaviviruses, including YFV, Tick-borne Encephalitis Virus (TBEV), JEV, and DENV, have been studied comprehensively for successful vaccine development (14,72,82,88). Also, a purified inactivated vaccine (PIV) against YFV with satisfactory immunogenicity results has been developed (81). Current flavivirus vaccine results make it plausible to develop live attenuated and inactivated vaccines for ZIKV. Other vaccine approaches for ZIKV include DNA vaccines, subunit vaccine, and viral vector vaccine platforms expressing viral structural proteins.
Emerging evidence in mice and R. macaque model reveals that PIV, DNA-based platform, mRNA, and virus-like particle (VLP)-based platform elicit reasonable tolerability and immunogenicity against ZIKV (3,37,55,59). Eventually, five vaccines candidates, including DNA, mRNA, and VLP, were promoted to clinical trial phase 1 in late 2016 (10).
The novel DNA vaccine comprising the combination of ZIKV prM and Envelope gene segment (prME) was a breakthrough vaccine, which later progressed to the clinical trials phase 2 (2,87). In R. macaque, the immunogenicity ratio of full-length ZIKV prME DNA vaccine demonstrated higher response compared to the Env DNA vaccine (only the envelope protein segment) and PIV, indicating the significance of using two antigens simultaneously, as well as reduced the need for booster shots of vaccine (59). DNA-based vaccines have been found effective in mice and monkey; unfortunately, the level of immunogenicity tends to decline in human clinical trials (66).
Furthermore, murine model studies administrating a subunit vaccine involved two delivery methods, that is, MNA-ZIKV-rEfl and Ad5.ZIKV-Efl. First is a transcutaneous delivery technique, which involves carboxymethyl cellulose microneedle arrays (MNAs) to translate E protein of ZIKV in host cell, whereas, the second delivery system is an adenovirus-based delivery of E protein (Ad5.ZIKV-Efl). Both approaches induced immunogenic responses; MNA-based vaccines have a higher efficacy as there is no particular immune cross-reaction and the codelivery with an adjuvant induces a substantial increase in immunogenicity when compared with adenovirus. Adenovirus-based delivery provides a potent immunogenic response; however, the drawback of this delivery is the expression of an anti-adenovirus antibody, reducing its efficacy (Table 1) (55). mRNA-based vaccine approaches have also been tested, implementing two delivery methods for fused prM and E protein encapsulated in lipid nanoparticles (prME mRNA–LNP) (89) and delivered by Vesicular Stomatitis Virus (rVSV-ZprME) (18), inducing a higher titer of neutral antibodies against ZIKV E proteins.
A live attenuated vaccine may require only one dose to induce rapid immunity and establish a long-term protection. Conversely, a subunit vaccine requires multiple doses to induce prophylactic immunity, although it is safer and requires less time for development. Nevertheless, all the possible vaccination approaches should be examined to boost the vaccine development. Previously, the viral vector-based vaccines for relatively closer viruses, such as YFV, JEV, and DENV, have shown a greater success ratio compared to others. The clinical phase 1 trials are in progress for a measles-vectored vaccine–MV-ZIKA-101–which induces immunity against ZIKV prM and E proteins (1). To date, the pipeline of ZIKV vaccines denotes a rapid progress. Aforementioned phase 1 vaccine candidates are potentially promising; however, advancement to further phases is exclusively dependent upon their incessant efficacy.
Conclusion
ZIKV's emergence in association with congenital neuronal anomalies and GBS carries overall health concerns. The molecular structure of ZIKV proteins and their interaction with host proteins require further investigation. A rapid research response to reveal the mechanisms underlying ZIKV pathogenesis and treatment strategies is adding to the continuously evolving pool of knowledge. Mechanism adapted by ZIKV to escape from immune system somehow mimics related flaviviruses such as DENV, JEV, and WNV, while the minor differences account for the severity of ZIKV infection. Especially, ZIKV's E and NS1 proteins require comprehensive investigation to elucidate its pathogenesis. The neurotropic and teratogenic nature of ZIKV arise due to an extraordinary capacity to settle in immune-privileged sites by crossing blood–brain barrier and placental barrier. In addition, the recent findings suggest that ZIKV NS1 protein is evolving to induce higher infectivity and a complex of NS4A and NS4B proteins suppresses the development of NPCs. This may provide future insights to overcome the ZIKV pathogenesis. ZIKV research is at the initial stage; it requires time to overcome the therapeutic and diagnostic challenges. A better understanding of ZIKV's molecular mechanisms through in silico and in vivo studies may pave way for diagnostics, therapeutics, and vaccine development to overcome ZIKV epidemic threat.
Compliance with Ethical Standards
Research involving human participants and/or animals
This is a review study and it has been compiled by extensive review of current literature. The study does not involve any human participant and/or animal models.
Informed consent of patients was not required for this study as it is a literature review.
Footnotes
Author Disclosure Statement
The authors, who contributed for this review article, do not hold any kind of conflicts of interest.