
Abstract
Vaccines are essential tools for the prevention and control of infectious diseases in animals. One of the most important steps in vaccine development is the selection of a suitable adjuvant. The focus of this review is the adjuvants used in vaccines for animals. We will discuss current commercial adjuvants and experimental formulations with attention to mineral salts, emulsions, bacterial-derived components, saponins, and several other immunoactive compounds. In addition, we will also examine the mechanisms of action for different adjuvants, examples of adjuvant combinations in one vaccine formulation, and challenges in the research and development of veterinary vaccine adjuvants.
Introduction
I
To date, numerous compounds of organic, inorganic, synthetic, and natural origin demonstrate an ability to stimulate immune responses and potent adjuvant properties (138). These substances can be broadly divided in two different groups—immunostimulants [saponins, Toll-like receptor (TLR) agonists, cytokines] and delivery agents (emulsions, microparticles, mineral salts) (17,97). Immunostimulants stimulate the antigen-presenting cells (APCs) and promote the secretion of various cytokines. On the other hand, delivery agents help to preserve the conformation of antigens (Ag) for proper presentation to APCs and provide a slow release for continuing immune stimulation. For instance, TLR agonists and other immunostimulatory substances can enhance immune cell recruitment and cytokine secretion, whereas emulsions and mineral salts can produce a depot effect at the injection site with a prolonged release of the antigen and continued stimulation of immune cells.
Vaccine development for food animals differs from that for human and companion animals. In humans, the health and well-being of the individual is of paramount importance, whereas, disease control measures for farm animals must be cost effective. While human vaccines can be marketed at more than a $100 per dose, the price for vaccines used in livestock industry is usually no more than a couple dollars per dose (64,83). Therefore, less financial resources are available for research and development of animal vaccines compared with that for human vaccines. On the other hand, livestock vaccine technology has fewer regulatory restrictions compared with the development of human vaccines (83). Thus, due to safety restrictions, some compounds are prohibited for use in human vaccines (Quil-A®, mineral oil-based emulsions), but are successfully employed in vaccines for livestock species. For more information about differences between human and veterinary vaccine research and development, the readers are referred to the review by Knight-Jones et al. (64).
In this article, the attention will be given to commercially available livestock vaccine adjuvants and adjuvants that are currently under investigation, including mineral salts, emulsions, TLR agonists, saponins, and other immunoactive compounds. The mechanisms behind their activity and synergistic effects in multiadjuvanted formulations will be considered.
Mineral Salts
Aluminum compounds were among the first adjuvants used in vaccines. It is well known that aluminum salt can induce high-titer IgG with a relatively long-lasting immunity, ease of formulation, and a long record of general safety. The early experiments with potassium alum (KAl(SO4)2:12H2O) administered with toxoid demonstrated that enhanced immune response in rabbits against the toxoid was due to the depot effect provided by the adjuvant (35). Since then aluminum compounds have been widely used in many commercially available vaccines for humans and animals (10,33,37). Examples in livestock applications include multivalent inactivated vaccines administered with aluminum compounds as adjuvants for calves and pregnant cattle against bovine herpes virus 1 (IBRV), bovine viral diarrhea virus type 1 (BVDV), parainfluenza virus type 3 (PI3V), and Mannheimia (Pasteurella) haemolytica (RESPIRAVAX™; MSD Animal Health, United Kingdom) (87).
There are several reports on the use of aluminum adjuvants in vaccines against economically important viral infections, including Newcastle disease and foot-and-mouth disease (FMD) (82,104,118), although their effectiveness against intracellular pathogens is extremely limited (73). Aluminum adjuvants promote the secretion of interleukin (IL)-4 cytokine, which stimulates T helper 2 (Th2) immune response. This induces the production of IgG1 and IgE immunoglobulins, and eosinophils (44,135), which makes these adjuvants good candidates for antibacterial and antiparasitic vaccines (37). To improve the efficacy of aluminum compounds against viral diseases, the strategy of its coadministration with other adjuvants to stimulate Th1 cell-mediated response has been employed in many research and clinical trials (44,102,146). The advantages and challenges of multiadjuvanted vaccines will be considered in detail in the last chapter of this review.
Calcium phosphate is another commercially available mineral salt adjuvant. It has been studied and used in vaccines against various toxoids and viral pathogens (40,41,113,114). Several studies have demonstrated that calcium phosphate induces less local tissue irritation when compared with aluminum adjuvants, which can be attributed to the reduced IgE antibody production (40,136). However, calcium phosphate and aluminum hydroxide have comparable potency in stimulating antigen-specific humoral immune responses in a side-by-side study (1). DNA vaccine administrated with calcium phosphate nanoparticles induced sufficient humoral and cell-mediated immune responses to protect animals against lethal challenge with FMD in mouse and guinea pig models (57). Inactivated Newcastle disease vaccine with calcium phosphate as adjuvant was less effective in producing protective immunity than vaccine administered with the natural polymer, chitosan (139).
Nonetheless, mineral salts are relatively safe and cost-effective adjuvants with a long history in human and animal vaccinology (36). They demonstrate good adjuvanticity in vaccines against extracellular pathogens; however, their application in vaccines targeting intracellular bacterial infections is limited.
Emulsions
As with mineral salts, emulsions have been employed as adjuvant systems in animal vaccines for a long time. Emulsions are formed when two immiscible liquids are brought together; one of them can organize into small droplets, dispersed within the other, and stabilized by an interfacial surfactant layer. Emulsions are also a good choice for animal vaccines because they are relatively simple to produce, cost effective, and demonstrate good efficacy in the production of antibody responses (17).
Different types of emulsions have different mechanisms of action in vaccine formulations (Fig. 1). Water-in-oil (W/O) emulsion is a dispersion of water droplets within continuous oil phase (Fig. 1a). Antigen is entrapped in the water phase surrounded by a continuous oil phase and slowly released upon breakdown of oil after injection (43). The depot effect at the injection site preserves the antigen from fast clearance by phagocytosis and the liver and, therefore, extends the time available for immune cell recruitment and antigen processing (17). The most well-known example of W/O emulsion adjuvants are Freund's adjuvants (30). Based on the paraffin oil and heat-killed and dried mycobacteria (Freund's complete adjuvant, FCA) or without mycobacteria (Freund's incomplete adjuvant, FIA), these adjuvants are very efficient in inducing high-titer antibody responses. However, Freund's adjuvants produce strong adverse reactions, such as local inflammatory lesions, pain, and distress, which have prevented their use in animal and human vaccines (127). Successfully commercialized W/O emulsions are available under the brand name Montanide™ Incomplete SEPPIC Adjuvants (ISA) (SEPPIC, France). These adjuvants show effectiveness on the level of FIA, but have fewer side effects (55,71) and are utilized in veterinary vaccines. ISA contain purified mineral or squalene oil and mannitol esters as emulsifiers. W/O Montanide adjuvants have been used in vaccines for Newcastle disease in chickens (3), subunit vaccine for Mycoplasma hyopneumoniae in pigs (56), inactivated vaccine for FMD in swine (50) and cattle (52,63), and farmed fish vaccines for various viral and bacterial infections (132,137).
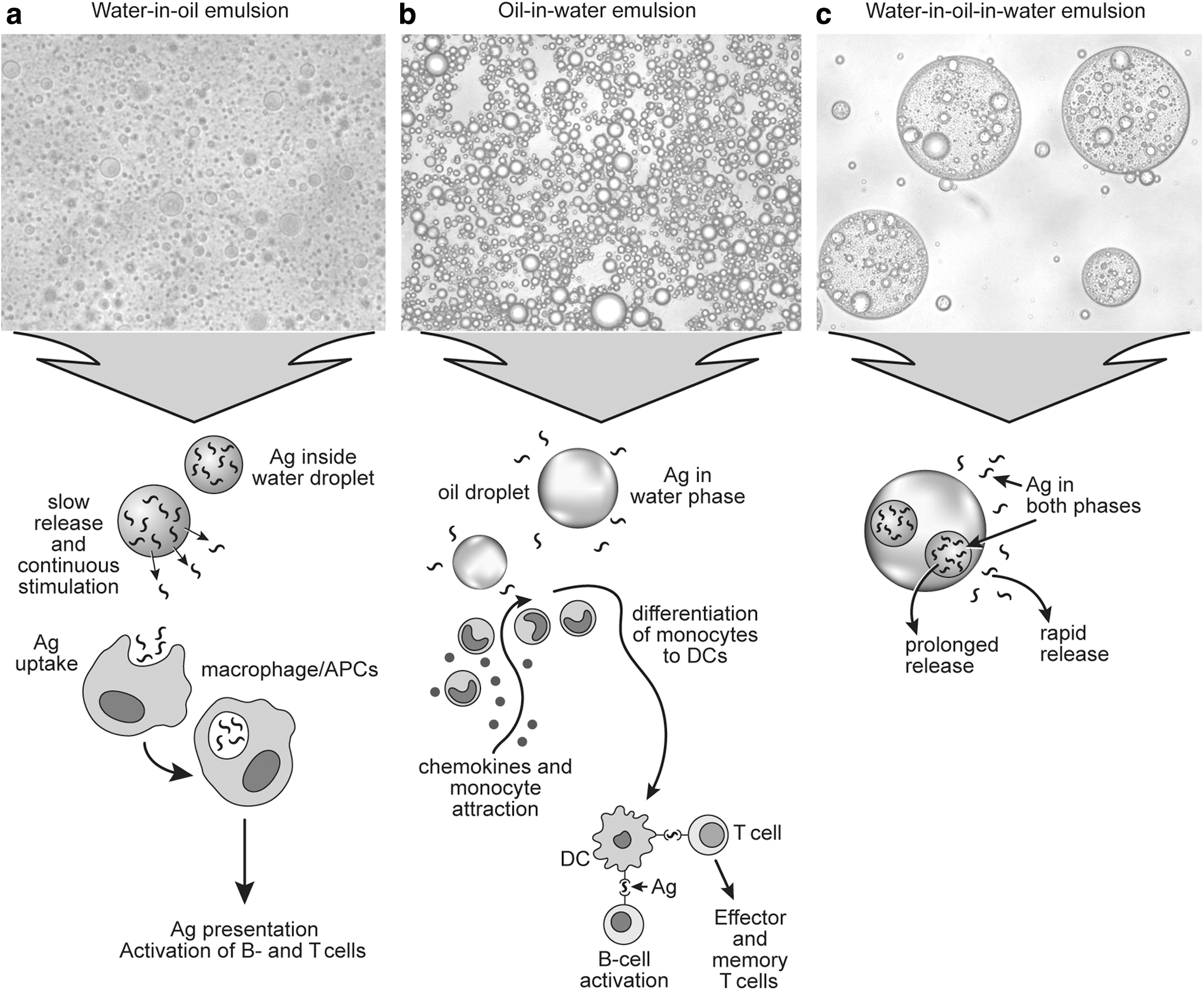
Microscopic images of different types of emulsions and their mechanisms of action.
Another type of emulsion utilized in vaccines is oil-in-water (O/W) emulsions, formed by the dispersion of oil droplets in the aqueous phase (Fig. 1b). Studies on MF59 (a squalene O/W vaccine adjuvant) has shed some light on how O/W emulsions induce immune responses (95). Unlike W/O emulsions, O/W emulsion-based adjuvant does not form an antigen depot at the injection site. Instead, the oil droplets facilitate the chemokine-driven immune cell recruitment and the differentiation of macrophages and dendritic cells (DCs) (23,119). MF59 demonstrated better adjuvanticity in stimulating cell-mediated immune response against influenza virus than aluminum hydroxide or calcium phosphate (140).
For veterinary applications, several commercially available O/W adjuvants exist under the brands of Montanide ISA, Emulsigen® (MVP Technologies, USA), and MetaStim® (Fort Dodge Laboratories, USA). These adjuvants are used in livestock vaccines against various economically important bacterial and viral antigens (4,8,45,68,141). For example, MetaStim coformulated with inactivated equine influenza virus stimulated higher expression of IL-12 and interferon-γ (IFN-γ) than the vaccine with aluminum phosphate as adjuvant (45). Recently, a new O/W emulsion adjuvant based on mineral oil and an inexpensive food-grade plant-based emulsifier was shown to be effective in vaccine formulations against swine influenza virus, M. hyopneumoniae, and classical swine fever (CSF) virus (32,75). These research findings suggest that O/W emulsions are more suitable adjuvants for vaccines against viral pathogens than mineral salts.
In attempts to overcome the issues with local reactions and high viscosity associated with W/O emulsions, research efforts have been devoted to develop multiphasic water-in-oil-in-water (W/O/W) emulsions (Fig. 1c) as vaccine adjuvants (11,31,70). In W/O/W emulsions, oil droplets containing internal water droplets are dispersed into continuous water phase. It has been speculated that W/OW emulsions may provide both fast release of antigens from the external water phase and prolonged release from the internal water droplets. Thus, this type of vaccine may quickly and continuously stimulate immune cells (11). Multiple emulsions have a very fragile structure and their formulations present great challenges. Currently, only few W/O/W emulsion adjuvants are available on the market. Oil-based formulations, Montanide ISA 201 and 206, produce multiple emulsions after gentle mixing with antigenic aqueous phase. These adjuvants demonstrate effectiveness and provide protection for different livestock species against several economically important pathogens, including influenza and FMD viruses (FMDVs) (9,48,80).
The design of emulsions for adjuvant application involves several parameters. The nature and the amount of emulsion components have a significant impact, not only on the physical and chemical stability of the final products, but also on the level of immune response and adverse reactions after vaccination (54,96). The choice of oil for the emulsion should also be considered. Several studies demonstrate that poorly metabolized mineral oil has superior efficacy in vaccine formulation compared with plant-derived oils (54,128). In addition, the proper concentration of surfactants should be determined to produce a stable emulsion, avoiding excessive amounts. Adverse reactions have been reported after vaccination with high concentrations of surfactants in emulsions (96).
The insufficient knowledge in the relationship between the size of emulsion droplets and the quality and quantity of immune responses is a major limiting factor for rational design of emulsion-based adjuvants. Recently, one study has demonstrated that emulsions with droplet size of 160 nm produce significantly higher antibody responses in mice than emulsions with 20 nm droplet diameter (120). However, another report speculates that particles with a size range of 10–200 nm can move faster and enter the lymphatic system and target the immune cells more efficiently (77).
Emulsion-based adjuvants are a very attractive selection for livestock vaccine formulations. These adjuvants can be formulated using inexpensive, readily available components such as mineral oil and food-grade emulsifiers. Further investigation is needed to fully understand the relationship between physical characteristics of emulsions and their ability to induce immune responses in animals.
Toll-Like Receptor Agonists
When Freund and his coworkers observed that the addition of killed mycobacteria to mineral oil emulsion improves the antibody production in rabbits and guinea pigs (30), the mechanism behind this phenomenon was unknown. Modern science has made a significant progress in understanding why certain bacterial-derived components can induce specific immune responses. TLRs are membrane-anchored receptors expressed in macrophages and DCs that recognize specific components of microbes and initiate innate host defense mechanisms (133). Activation of TLRs can contribute to the induction of antigen-specific immunity by inducing the expression of certain cytokines involved in T cell differentiation (Fig. 2). For example, lipopolysaccharides (LPS), a structural component of Gram-negative bacteria surface membrane, is a TLR4 agonist that can stimulate APCs to secrete IL-1β, IL-12, IL-18, IL-23, and IL-27. These cytokines are responsible for the activation of Th1 cells, which mediates cell-mediated and humoral immunity against viruses and other intracellular pathogens (69). Monophosphoryl lipid A (MPLA), a low-toxic component of LPS, is used in licensed human vaccine for Hepatitis B (Supervax™; Dynavax Technologies, USA) (126). MPLA combined with aluminum hydroxide (AS04; GlaxoSmithKline, United Kingdom) improves cell-mediated immune responses to hepatitis B antigen compared with the vaccine formulated with aluminum hydroxide only (94). Studies on MPLA as an adjuvant for commercial animal vaccines have yet to be reported. Other examples of TLR agonists include bacterial lipoprotein and flagellin that are recognized by TLR2 and TLR5, respectively. In addition, TLR9 can identify cytosine phosphate guanine (CpG) motifs in bacterial DNA and induce T cell-mediated immunity (134).
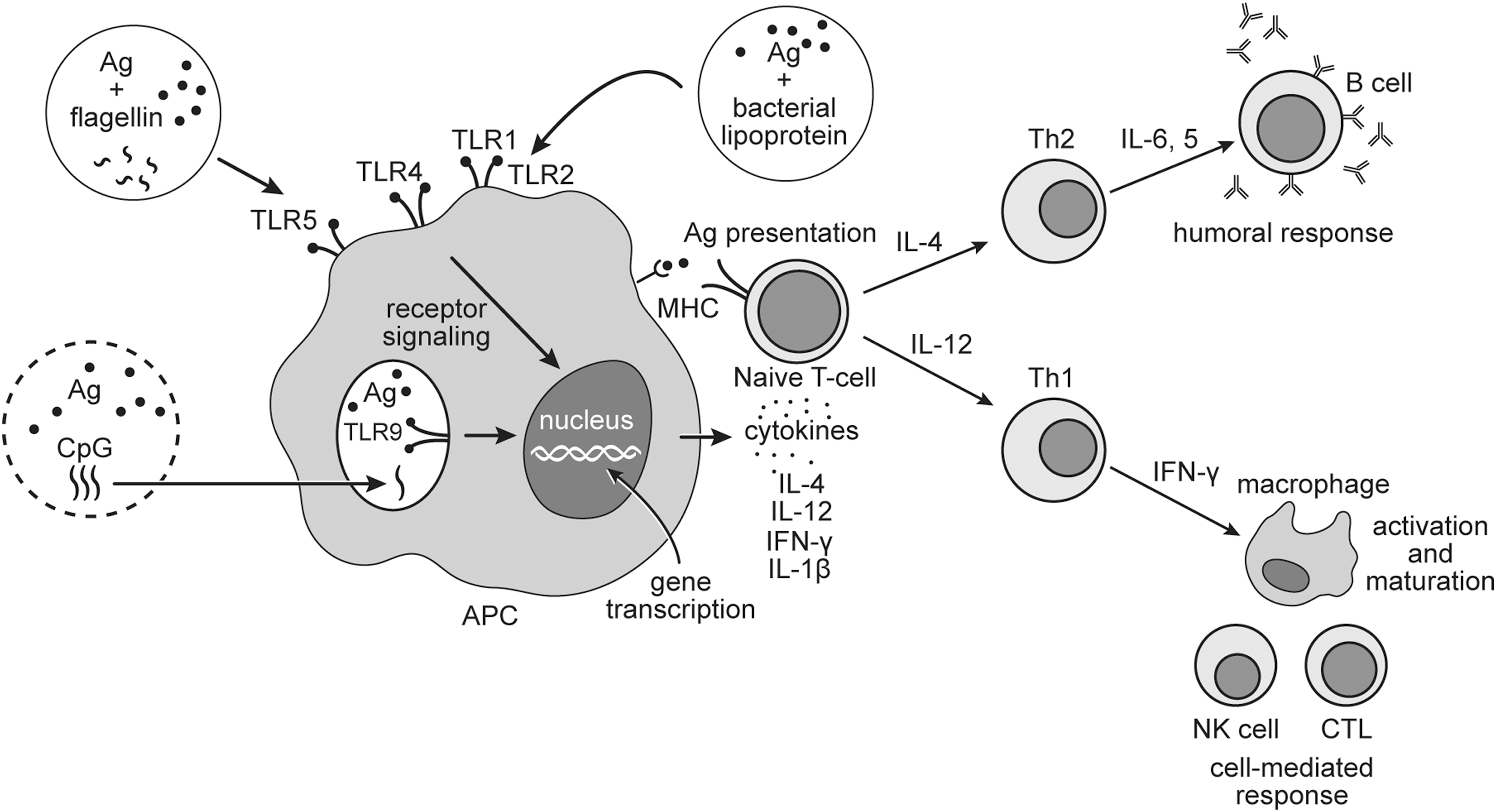
Mechanism of activation of the Ag-specific cell-mediated immunity by TLR agonists. Microbial components interact with TLRs that lead to APC maturation and activation of the signaling pathways resulting in immune responses. APC presents Ag through MHC class II molecules and activates naive T cells. APC secretes Th1-polarizing cytokines (IL-12) or IL-4, driving toward Th2-cell response. APC produces IFN-γ and IL-1β, which promote the activation and maturation of leukocytes. Th1 cells promote maturation and activation of NK cells and CTL through secreting IFN-γ, whereas Th2 cells produce IL-4, -5, -6, and other cytokines and activate B cells. CTL, cytotoxic lymphocytes; IFN, interferon; IL, interleukin; MHC, major histocompatibility complex; NK, natural killer; Th, T helper; TLR, Toll-like receptor.
Recent examples of TLR agonist utilization in livestock vaccines include the use of Pseudomonas aeruginosa membrane lipoprotein OprI in a CSF subunit vaccine (110). OprI promotes T cell and humoral responses through TLR2-mediated activation and maturation of APCs. The bacterial protein flagellin, a TLR5 agonist, has also demonstrated adjuvant properties in vaccine formulations. Guinea pigs that received flagellin-adjuvanted vaccine developed higher levels of anti-FMDV neutralizing antibodies than that from animals received the vaccine without flagellin (39). When formulated with rabies virus-like particles, both flagellin and heat-labile bacterial enterotoxin, can promote the production of virus-specific neutralizing antibodies and CD4+ and CD8+ T cells in dogs (107). Escherichia coli heat-labile enterotoxin was also employed as an adjuvant in subunit swine influenza virus vaccine (131). CpG oligodeoxynucleotides (ODNs) interact with TLR9 and stimulate Th1 response through the induction of IL-12 and IFN-γ (134). CpG ODNs administered with inactivated avian influenza virus in intranasal vaccination of chickens induced stronger immune responses than antigen alone (145). Fusion of CpG motifs with chicken anemia virus proteins helped to improve the efficacy of this subunit vaccine for poultry (47).
Another interesting approach in the development of TLR agonist-based adjuvants is the application of bacterial ghosts (BG) (81). These nonliving cells prepared from Gram-negative bacteria are free from cytoplasmic contents with preserved cellular morphology and surface antigens, such as LPS, peptidoglycans, and MPLA (53). Pigs treated with BG-adjuvanted chimeric vector-based vaccine rAdV-SFV-E2 were completely protected from CSF virus challenge, in contrast to the limited protection in pigs treated with the vaccine without BG (143). However, temporal allergic reactions were also observed in pigs receiving the BG-adjuvanted vaccines.
Although TLR agonists have attracted considerable attention in veterinary vaccine technology, their applications possess challenges as well. Overactivation of the innate immune system through pathogen-recognition receptors (PRRs) can lead to septic shock (59). The presence of heat-killed mycobacteria in the FCA promoted higher antibody responses compared with FIA (30,127). However, high toxicity and delayed-type hypersensitivity reactions were also associated with this adjuvant (108). In a recent study, recombinant bacterial flagellin administered in combination with an O/W emulsion in a vaccine against malignant catarrhal fever induced an immunosuppressive mechanism in cattle (68). As a result, higher fatalities were observed in comparison with the group receiving the vaccine without flagellin. Hence, it is important to find the right balance between safety and efficacy as well as to develop a better understanding of the relationship between the structure of these adjuvants and the immune response they induce in animals.
Immunoactive Compounds
Cytokines
Cytokines are key players of TLR agonist-mediated immune responses. The use of cytokines as vaccine adjuvants is being investigated intensively in animal vaccines (72). These small proteins play crucial roles in cell signaling, inducing, and directing cellular and humoral immune responses. For instance, IL-2, IL-12, and IFN-γ facilitate the stimulation of Th1 cells responsible for cell-mediated immune responses against intracellular pathogens. On the other hand, cytokines IL-4, IL-5, and IL-10 promote the development of Th2 cells and antibody production, which are important in host defense against extracellular pathogens. In addition to promoting Th1-cell activity, IFN-γ also enhances the uptake and presentation of antigens by inducing the expression of major histocompatibility complex (MHC)-II molecules on APCs (42). In a recent study, pigs immunized with attenuated CSF vaccine with IFN-γ as an adjuvant developed significantly higher levels of anti-CSF virus IgG antibodies and increased the expression of MHC-I and MHC-II molecules compared with groups that received the attenuated vaccine alone (26). Coadministration of porcine IFN-α with subunit FMD vaccine promoted the production of antigen-specific antibodies and protected pigs from FMDV challenge (15).
Cytokines are also used in vaccines for various viral diseases of poultry (5), bovine (7), and fish species (132). Recent examples include the incorporation of IL-18 in the vaccine for Newcastle disease (46) and IL-7 in DNA vaccine for infectious bursal disease (49). Bovine IL-18 was also used as an adjuvant in a DNA vaccine against FMD that induced cell-mediated and antibody responses in cattle. However, not all animals were protected from a challenge with the FMD virus (65). Atlantic salmon immunized with a DNA vaccine consisting of salmon anemia virus protein and fish type I interferon induced higher levels of antibodies and influx of B cells and CD8+ T cells compared with that immunized with a DNA vaccine without the cytokine (14).
Saponins
Saponins are amphipathic compounds composed of one or more hydrophilic glycoside moieties attached to a lipophilic triterpene derivative. These naturally occurring substances are used in many pharmaceutical applications. The ability to stimulate immune responses has promoted saponins as adjuvants in animal vaccines (61).
The most prominent saponin-based adjuvant is Quil-A (Brenntag Biosector A/S, Denmark). Quil-A is a heterogeneous mixture of water-soluble saponins extracted from Quillaja saponaria, a tree indigenous to South America. Due to its toxicity, Quil-A is not suitable for human vaccines (129); it is widely used for veterinary applications (18,19,116,144). It has been demonstrated that addition of Quil-A to vaccines for FMD and porcine reproductive and respiratory syndrome (PRRS) had improved their immunogenicity in comparison with vaccine groups that received no adjuvant. QS-21, the purified fraction of Q. saponaria, is currently in many clinical trials for human cancer and infectious vaccines (147). Moreover, QS-21 is used as an adjuvant in a commercially available vaccine for feline leukemia (78).
The combination of cholesterol, phospholipids, and purified fractions of Quil-A in immune-stimulating complexes (ISCOMs) helps to improve the stability and reduce the toxicity of saponins (92,130). ISCOMs have cage-like structure that assists in preserving and delivering the antigen to APCs. In addition, the particulate nature and size of the complexes efficiently promote antigen phagocytosis by the APCs (112). ISCOMs can effectively stimulate both CD4+ and CD8+ T cell responses. Thus, ISCOMs have been employed in licensed vaccines for horses against influenza virus in several European countries (100). These complexes have also been utilized in a subunit vaccine against herpes equine virus 2 (EHV-2). This vaccine helped to protect young horses against EHV-2 challenge (93). Several studies also demonstrate that it can efficiently induce the production of neutralizing antibodies and enhance protective immunity against BVDV in sheep and calves (13,58).
Studies on the immunoregulatory activities of Quil-A and QS-21 have demonstrated that they can elicit cell-mediated immune responses with the stimulation of both Th1 and cytotoxic lymphocytes (61,62). Q. saponins can induce the production of IgG1 and IgG2a antibodies in mice (60). Substantial efforts have been made to understand the interconnection between the structural components of saponin and their immunological functions (Fig. 3). Saponins are currently not recognized as a TLR ligand (105). However, the sugar groups of saponins can bind to the lectins on APCs. This promotes antigen phagocytosis and stimulates APCs to secrete cytokines responsible for the activation of humoral and cell-mediated responses (79). It was hypothesized that the aldehyde group on the triterpene domain of the saponin molecule interacts with T cell surface receptors and facilitates its costimulation (79,124). This knowledge is useful for the development of synthetic analogs of naturally derived saponins and the design of saponin molecules with preserved immune activity and reduced toxicity (27). For livestock applications, the approach of searching in nature for Q. saponaria analogues with less toxic saponins would be more economically feasible. For instance, the extract from Q. brasiliensis leaves has been shown to be less toxic, but it demonstrates similar cellular immune response in a murine model compared with saponins from Q. saponaria (123).

Structural representation of saponin molecule. Saccharide moieties interact with APC receptors and promote Ag phagocytosis, and the aldehyde group on the triterpene domain provides costimulation of T cell receptors.
Polymers
Polymeric substances are often utilized in vaccination as delivery vehicles to preserve the antigen and provide its controlled release after injection (28,117). In addition, some polymers may also act as immunostimulants. In this study, several examples of the immunoactive natural and synthetic polymeric substances as veterinary vaccine adjuvants will be reviewed.
The natural polymer chitosan is employed not only in vaccine technology, but also in drug delivery and antibacterial applications (66). This linear polysaccharide is a product of chitin deacetylation, composed of randomly distributed β-(1 → 4)-linked
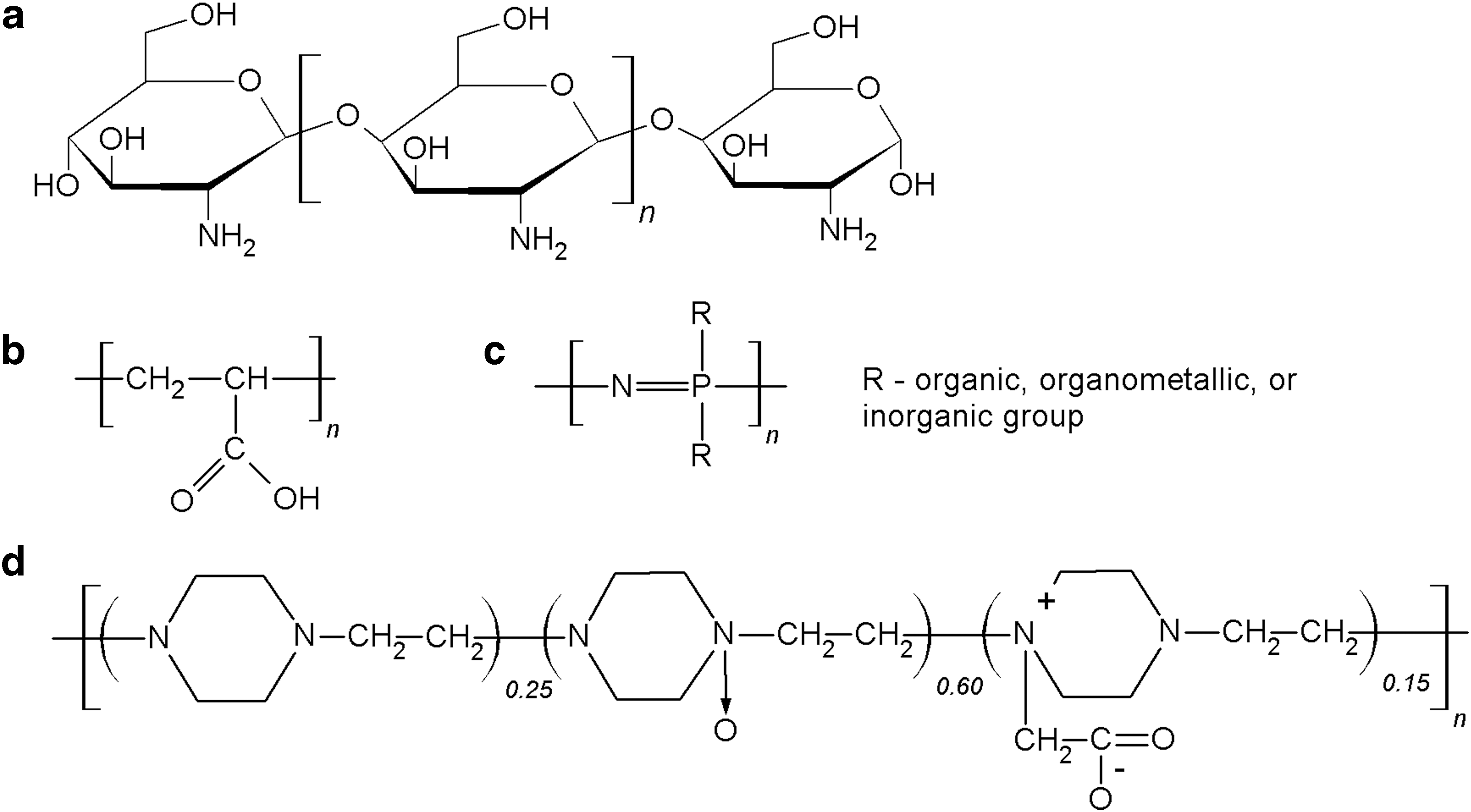
Chemical structures of polymeric adjuvants:
Several synthetic polymers have also been tested as vaccine adjuvants. Polyacrylic acid polymers (carbomers) (Fig. 4b) such as Carbopol® (Lubrizol Advanced Materials, Inc., USA), have been utilized in veterinary medicine since 1970s. The adjuvant properties of carbomers were first recognized in a murine vaccination experiment (22). These polymers are currently employed as adjuvants in inactivated and vector-based licensed vaccines against influenza virus for horses (88,100), in swine vaccines (ImpranFLEX®; Boehringer Ingelheim, Germany) against porcine circovirus type 2 (25) and M. hyopneumoniae (98). Recently, Carbopol has been shown to effectively promote IFN-γ production and T cell differentiation in pigs after vaccination with modified live PRRS virus vaccine (76) and to improve the efficacy of DNA vaccine against equine herpes virus (84). The mechanism of carbomer-mediated adjuvanticity has yet to be elucidated. Previous studies proposed the ability of these crosslinked polymers to entrap and slowly release antigenic molecules stimulating immune cells (67). Recent research findings suggest that polyacrylic acid polymer can promote Th1 cell responses through improved uptake of antigens by APCs without the activation of PRRs (34).
In contrast to carbomers, polyphosphazene polymers have been studied mostly in laboratory settings and yet to reach commercialization (89,103). Polyphosphazenes are a diverse group of hybrid inorganic–organic polymers with alternating phosphorus and nitrogen atoms in the backbone (Fig. 4c). Instead of the depot effect (103), polyphosphazene polymers tend to form noncovalent bonds with antigen molecules, therefore, assist with the delivery of antigens to APCs (2). Increased secretion of proinflammatory cytokines and chemokines at the injection site is also associated with this adjuvant (6). Polyphosphazenes have been reported to increase the secretion of IgG and IgA antibodies in the mucosa compared with vaccine alone and vaccine with CpG oligonucleotides as adjuvant in a mouse model (122). Although polyphosphazenes demonstrate promising results in laboratory animals, there are limited data on using them in farm animals. It was reported that polyphosphazene-adjuvanted Actinobacillus pleuropneumoniae vaccine can increase IgG antibody titers and IFN-γ production in pigs compared with O/W Emulsigen-adjuvanted vaccine (20). In contrast to the emulsion-based adjuvant, polyphosphazene did not induce inflammation at the injection site in pigs.
Another example of synthetic polymeric adjuvant is Polyoxidonium® (Petrovax Pharm, Russia). Polyoxidonium is a copolymer of 1,4-ethylenepiperazine, 1,4-ethylenepiperazine-N-oxide, and (N-carboxymethylene)-1,4-ethylenepiperazinium bromide (Fig. 4D) (106). This polymeric compound induces the production of IL-6, tumor necrosis factor-alpha (TNF-α), and IL-1β cytokines by mononuclear blood cells in a dose-dependent manner in vitro (24). Live vaccine administered with polyoxidonium as adjuvant elicits high protective immunity in guinea pigs against Brucella abortus (21). This adjuvant is also licensed in human vaccine for influenza in Russia and CIS (106).
Polymeric substances are very attractive adjuvant candidates for vaccination against viral infections, including veterinary applications. Polymers are known to have a better safety profile than emulsion-based adjuvants, and are capable of inducing cell-mediated immune responses that are important for host defense against viral pathogens.
Combination of Adjuvants
The combination of two or more adjuvants in one vaccine formulation may lead to improved efficacy. A “perfect” adjuvant can deliver the antigen in its native conformation to APCs, stimulate the secretion of the right cytokines, and activate effective humoral and cell-mediated immune responses. Due to the difficulty in finding one substance that can perform all these functions, incorporating different adjuvants in one vaccine formulation and their potential synergistic effects are being intensively examined (85,86). Several adjuvant combinations are already in different stages of clinical trials in human vaccines against infectious diseases and cancer (38,86). Results from these studies may be used to design adjuvants for animal vaccines.
The success of FCA in the production of high antibody titers encouraged many researchers to combine emulsions and bacterial-derived components in one vaccine formulation. Several research groups have evaluated the combination of bacterial CpG motifs with O/W or W/O/W emulsions as adjuvant systems in vaccines for avian influenza and FMD (51,74,91,115). The mixture of CpG ODNs and emulsions can induce higher levels of neutralizing antibodies than these adjuvants alone. Emulsions provide sustainable release of the antigen and CpG ODNs, whereas the later can activate IFN-γ-secreting lymphocytes (142). The combination of inactivated spores of Bacillus atrophaeus and Q. saponins enhanced the antibody responses against rabies virus in a murine model in comparison with vaccine groups formulated with spores or saponins alone (99). The combination of saponins and emulsions was also shown to improve the efficacy of FMD vaccines (16,144). QS-21 accelerated the generation of anti-FMDV neutralizing antibodies in cows immunized with the vaccine with W/O/W emulsion adjuvant. In an experiment with mice vaccinated against FMD, the combination of saponins and emulsion induced higher titers of IgG1, IgG2a, IgG2b, and IgG3 antibodies (125).
Although several promising adjuvant combinations have been tested in laboratory settings, further investigations are needed to confirm their safety and efficacy in large animals. Moreover, the impact of combining different adjuvants on manufacturing costs has to be evaluated thoroughly before it can be applied to make commercial animal vaccines.
Conclusion
Numerous natural and synthetic substances can be used as adjuvants to improve the efficacy of animal vaccines. Some of them, such as aluminum compounds, emulsions, saponins, and carbomers, have already been used in licensed products; whereas others (cytokines and polyphosphazenes) are still evaluated experimentally. Several criteria should be considered in selecting/developing adjuvants for animal vaccines: effectiveness in the target animal species, induction of a rapid and long-lasting protective immunity, safety for animals, compliance of food safety regulation, feasibility for scale-up production, and last but not the least, cost effectiveness. Finding the appropriate adjuvant or their combinations to meet the above criteria is one of the major challenges in animal vaccine development.
Footnotes
Acknowledgments
This research was supported by an award from the National Bio and Agro-Defense Facility Transition Fund and a USDA ARS Specific Cooperative Agreement 59-5430-001-23S, NP-103. The authors would like to thank Mal Hoover, medical illustrator of the College of Veterinary Medicine, Kansas State University, for creating the illustrations for this article.
Author Disclosure Statement
No competing financial interests exist.