
Abstract
Commercial broiler and layer chickens are heavily vaccinated against economically important viral diseases with a view of preventing morbidity, mortality, and production impacts encountered during short production cycles. Hatchery vaccination is performed through in ovo embryo vaccination prehatch or spray and subcutaneous vaccinations performed at the day of hatch before the day-old chickens are being placed in barns with potentially contaminated environments. Commercially, multiple vaccines (e.g., live, live attenuated, and viral vectored vaccines) are available to administer through these routes within a short period (embryo day 18 prehatch to day 1 posthatch). Although the ability to mount immune response, especially the adaptive immune response, is not optimal around the hatch, it is possible that the efficacy of these vaccines depends partly on innate host responses elicited in response to replicating vaccine viruses. This review focuses on the current knowledge of hatchery vaccination in poultry and potential mechanisms of hatchery vaccine-mediated protective responses and limitations.
Introduction
P
Maintaining closed flocks with strict biosecurity measures are commonly implemented by the commercial poultry operations to prevent the introduction and spread of avian viruses. Such biosecurity measures include identification of all potential sources of infections and exercising steps to prevent direct and indirect transmission of the viruses into poultry flocks. Such potential sources of infections include water sources that are contaminated, animal or human movements into the flocks, live poultry markets, and contaminated vehicles and equipment (107). Although biosecurity measures, including strict regulations and quarantine practices are considered excellent ways of protecting commercial poultry operations from avian viral diseases, they are not adequate as sole means of disease prevention (114).
Vaccination, another common approach controlling avian viral diseases, is based on administration of antigenic molecules, including whole or parts of pathogens that stimulate the host's adaptive immune responses against a particular pathogen. Unlike human vaccines, poultry vaccines mostly prevent or reduce morbidity and mortality resulting from an infection rather than the infection (88). However, some vaccines could potentially reduce viral shedding and subsequent transmission (119).
Chickens are intensively vaccinated farm animals with an average of 12–20 vaccine administrations per bird done within a production cycle. This intense use of vaccines by the poultry industry is driven by the economic impacts of the diseases and the poultry vaccine administration begins prehatch at the hatchery. One potential implication of this approach plus the inherent deficiencies of the currently available poultry viral vaccines may be the vaccination failures.
In the last four decades, a number of poultry viral vaccines were made available as hatchery vaccines to administer either prehatch or immediately posthatch. Although hatchery vaccination has progressed substantially during this period and become a poultry industry practice for controlling a number of economically important poultry viral diseases globally, the reviews with a focus on this subject are scarce (82,91). Recently, a few reviews focusing on other aspects of vaccination are published for common poultry viral infections, such as infectious bronchitis virus (IBV), infectious laryngotracheitis virus (ILTV), Newcastle disease virus (NCDV), Marek's disease virus (MDV), and avian influenza virus (AIV) infections (25,37,59,89,108). Therefore, this review discuses current knowledge of hatchery vaccination-mediated mechanisms of host responses and potential limitations of this approach providing field examples.
History of Vaccination Against Poultry Diseases
The era of vaccination for disease control began in 1879, with the introduction of Louis Pasteur's live attenuated fowl cholera vaccine, first ever laboratory-developed vaccine against any pathogen in any species. Then, first licensed poultry viral vaccine was introduced against fowlpox in 1918 (30). Subsequently, the development and licensing of poultry viral vaccines increased rapidly over time. The first vaccine for infectious laryngotracheitis (ILT) was licensed in United States of America in 1933 and vaccines for Newcastle disease (ND), infectious bronchitis (IB), avian encephalomyelitis, infectious bursal disease (IBD), and Marek's disease (MD) were licensed in United States of America in 1946, 1953, 1962, 1967, and 1968, respectively (30,31).
Ultimately with further advancement in the field of vaccine development, the concept of recombinant vaccine using pox virus as a vector was introduced in 1982 (30,31). The first deoxyribonucleic acid (DNA) vaccine against a poultry disease was investigated in early 90s (92), although no DNA vaccines made into the market so far for use as hatchery vaccines against poultry diseases.
History of Hatchery Vaccination
The practice of hatchery vaccination was first introduced in the late sixties for subcutaneous immunization of birds immediately posthatch against MD using a live attenuated MDV (14). A second vaccine consisting of live herpes virus of turkeys (HVT) was introduced soon and became popular as a hatchery vaccine for MD control (84). After a decade, a third vaccine consisting of live MD virus type 2 vaccine (SB-1 strain) and a combined HVT-SB-1 vaccine have been made available for hatchery vaccination against MD (64). Almost all these vaccines are cell associated and therefore effective and efficacious, when subcutaneously administered on the day of hatch, even in the presence of maternal antibody to MDV (103), which is important since the virus is distributed ubiquitously such that most of the chickens develop antibodies early in life. Subsequently, in ovo vaccination was introduced as a mean of hatchery vaccination against MD in 1982 (102), against IBD in 1995 (40) and recently against ND, ILT, IBV and fowlpox (59,75,96,102,126).
Although, the presence of high levels of maternal antibodies in the vaccinated animals may neutralize or interfere with the replication of live vaccine strains, the real reasons for adapting in ovo vaccination as a common practice worldwide are manifold: First, inducing early immune responses leading to more optimal protection of the day-old chickens against circulating pathogens in the environment at the time of placing them in the barn than posthatch vaccination. Second, high output automated in ovo vaccine delivery equipment results in reduced labor cost at the farm level. Third, in ovo vaccination reduces stress in chickens associated with animal handling. Finally, in ovo vaccination ensures precise uniform delivery of vaccines compared with other mass vaccination techniques such as coarse spray or water vaccination.
Types of Vaccines Used in Hatcheries
Various types of vaccines, such as live (84), live attenuated (14), recombinant viral vector (23), DNA (60), and immune complex (56) vaccines have been investigated to be used in hatchery settings with various success rates. Live attenuated vaccines are prepared from the live microbes, but their pathogenicity has been weakened in the process of attenuation and capable of eliciting protective cell- and antibody-mediated immune responses in the host.
In the recombinant vaccines, the DNA sequence coding for antigen of a pathogen is inserted into a genome of an attenuated vector virus such as herpesviruses or poxviruses (106). The resultant vector is then used as vaccines resulting in the expression of target antigenic molecules stimulating both cell- and antibody-mediated immune response. The multiple genes coding for the antigenic molecules of wide range of viruses can also be inserted into a vector to produce live polyvalent vaccines. Whereas, in DNA vaccines, the naked genes coding for antigenic molecules of pathogens will be introduced into the host and some host cells will take up that constructed DNA to produce the antigenic molecules. Therefore, own host cells will function as vaccine-producing factories, stimulating antibody-mediated immune response by free-floating secreted antigen and cell-mediated immune response by displayed antigen in the host cell surface (48). The DNA vaccines are safe, inexpensive, and easy to produce than conventional vaccines. Furthermore, DNA vaccines are unable to cause diseases because they do not contain whole microbes. Additionally, in DNA vaccines, the antigenic molecules are produced from own host cells, as such it minimizes the other potential side effects.
Recently, in ovo immune complex vaccines have been developed by mixing the vaccine virus with chicken antisera (56). The formation of viral antigen complex with specific antibodies, results in modulation of antigen presentation, receptor signaling, and immune responses. In addition, immune complex vaccines are believed to play a crucial role in the generation of prolonged humoral immune responses (110). It has been shown that the immune complex antigen coated with antibodies are 100 times more efficient in inducing antibody-mediated immune response compared with native antigen (63).
The live, live attenuated, and recombinant viral vector vaccines are capable of replicating in the host for an extended period of time, thus capable of eliciting solid adaptive immune responses in addition to innate host responses (10,28,55). Currently, the majority, if not all hatchery vaccines, are either live, live attenuated, or recombinant viral vector vaccines. Although the live attenuated vaccines are closest to a natural infection and suitable for hatchery vaccination, regaining virulence of the wild viruses has been a concern (113). Although killed vaccines cannot revert to virulence (113), elicitation of weak immune responses makes them unsuitable for hatchery vaccination. Also, there are a number of studies attending to demonstrate the effectiveness of DNA vaccines for poultry since early 90s against avian influenza (AI) (92), IBD (51), IB (60), ILT (13), and MD (115). The usefulness of these vaccines as hatchery vaccines is uncertain even if they are safe and inexpensive.
Hatchery Vaccination Techniques
Hatchery vaccination is performed through in ovo embryo vaccination at embryo day (ED) 18 and coarse aerosol spray or manual subcutaneous vaccination on the day of hatch before placing the day-old chickens in the barn with potentially contaminated environment.
In ovo vaccination has been introduced for the control of MD in early 80s (102). The efficacy of the number of in ovo vaccines, such as IB vaccine (122), reo virus vaccine (45), IBD vaccine (19), AI vaccine (98), ND vaccine (4), and multivalent vaccine against MD, IBD, ND, and fowlpox, administered through in ovo route has been explored in poultry with variable efficacies. The efficacy of viral vaccines administered through this route is variable with MD vaccine, particularly the HVT has been the most efficacious and ND vaccine has been the least efficacious (4,102).
Currently in ovo vaccination has become a standard practice for hatchery vaccination against many viral diseases, such as MD, IBD, ILT, ND, and fowlpox (82,102,109,112,121). This technique facilitates the precise delivery of vaccines compared with conventional posthatch vaccination techniques, such as injection and spray vaccination. Although the optimum time for in ovo vaccination is between ED17.5 and ED19.2, it is a standard practice to perform the in ovo vaccination at ED18 in broiler eggs coinciding with transfer of eggs from setter to hatcher compartment to avoid turning in the setter compartment of the incubator (91).
In the hatcheries, IB and ND, live attenuated vaccines are administered as a coarse spray on the day of hatch, which are less time and labor consuming, efficient, and effective. Coarse spray of the vaccines produces droplets of various sizes ranging from 5 μM and above as such prevents vaccine virus penetration deep down the respiratory tract to prevent potential vaccine virus-mediated clinical reactions (59). The droplet size of 5 μM and above land largely to eyes and nares (18) and they should be confined to upper respiratory tract. Vaccine delivery as a coarse spray does not damage live attenuated virus particles as has been shown for IBV vaccines (94) and replicate to a higher titer in the upper respiratory tract (71) inducing protective immune responses (93). Coarse spray vaccine administered on the day of hatch also has been shown to induce protective immune responses against ND (39).
A subset of hatchery vaccines is also recommended to administer through subcutaneous route immediately after hatch. The subcutaneous immunization of chickens with live and live attenuated virus immediately posthatch was first introduced against MD (14,84) and still a popular hatchery vaccine for layer birds. Although handling individual chickens is labor intensive, the vaccines given through subcutaneous route are proven efficacious (85). Either HVT- or fowlpox-vectored ILT (116), ND (29), IBD (86), and AI (38) vaccines are also available as subcutaneous vaccines for hatchery vaccination.
Developmental and Functional Status of Avian Immune System at the Time of Hatchery Vaccination
Status of development of lymphoid organs at ED18 and around hatch is critical for successful hatchery vaccination outcomes. The primary lymphoid organs, such as bursa of Fabricius, is unique and it is the B cell organ of the birds. In chicken embryo, the bursa of Fabricius begins to develop as an epithelial bud at ED4 and invasion of bursa of Fabricius with B cells continuously occurs between ED8 and ED14 with establishment of around 10,000 follicles with well-defined cortex and medulla at hatching (50).
The second primary lymphoid organ, thymus, begins to develop as endodermal thymic rudiment on ED3 and clearly defined medulla and cortex are formed by ED12 (16). The secondary lymphoid organs, such as spleen, start developing around ED4 (130), and cecal tonsils by ED10 (81). It has also been recorded that the other secondary lymphoid units, such as bronchus-associated lymphoid tissues are present at the time of hatch and age-related changes are observable during early posthatch period (32).
One of the other factors that influences the success of hatchery vaccination is the functional status of the chicken immune system by the time of vaccination. Around the time when hatchery vaccination is performed, the innate immune system is more optimally developed than the adaptive arm of the immune system.
The development of the innate arm of the immune system begins during early embryogenesis and innate immune cells, such as dendritic cell (DC), macrophages, and natural killer (NK), are critical mediators of innate immune functions. The colonization of lymphoid organs by DC precursors begins between ED10 and ED15 depending on the lymphoid tissues (81). When the eggs are at 2.5–4.5 days of incubation, the macrophage progenitor cells with phagocytic activity appears in the yolk sac (21,83), which is followed by the appearance of macrophage activity in the liver and spleen at ED12 and ED16, respectively (36,105,127). The NK cells first appear at ED8 in the spleen (42) and by ED14 NK cells are functionally similar to the NK isolated from spleen of 4-week-old chickens (54).
Similarly, the innate pattern recognition receptor (i.e., toll-like receptors or TLRs) begins to develop around ED14 in the lungs and spleen (Fig. 1) (78). These innate immune receptors are capable of recognizing microbial pathogens, such as viruses, bacteria, and fungi, due to the presence of molecules that are highly conserved among them known as pathogen-associated molecular patterns (PAMPs). Furthermore, the innate effector molecules, such as type I and III interferons (IFNs), appear developing around ED14 and continue to develop simultaneously with type I and III IFN receptors (61). Altogether, by the time the hatchery vaccination is performed prehatch at ED18 or immediately following the hatch, innate immune system is equipped with all the necessary receptors and effector cells and molecules (Fig. 1).

Development of innate and adaptive arms of the immune system in chicken. Adapted from Abdul-Careem (1). Innate immune system development begins at early embryonic life with appearance of macrophage progenitor cells in the yolk sac at ED2.5, followed by the appearance of macrophage activity in the liver and spleen at ED12 and ED16, respectively. The colonization of lymphoid organs by DC precursors begins between ED10 and ED15, whereas colonization of spleen by the NK cells happens around ED8. Similarly, the innate pattern recognition receptor (i.e., TLRs) and the innate effector molecules, such as type I and III IFNs begin to develop around ED14. The adaptive immune system development takes place in the later part of embryonic life with further development of components of adaptive immune system taking place in the early posthatch period. By the time the hatchery vaccination is performed, prehatch at ED18 or immediately following the hatch, innate immune system is equipped with all the necessary receptors and effector cells and molecules. TLRs, toll-like receptors; IFNs, interferons; GALT, gut-associated lymphoid tissue; B cells, B lymphocytes; T cells, T lymphocytes; ED, embryo day; D, day; DC, dendritic cell; NK, natural killer.
Antibody-mediated immune response generated in response to vaccination is a critical function of the adaptive immune system. The development of IgM, IgA, and IgY (homologue for mammalian IgG) antibody isotypes have been described in birds (7,70). In the bursa of Fabricius, IgM synthesis appeared to begin at ED14 and IgY synthesis appeared to begin at ED21 (70). In the extrabursal tissues, IgM appears around ED17 and IgY is observable at 4 days posthatch. In the spleen, the expansion of lymphocytes containing IgM and IgY begins by day 3 and 8 posthatch, respectively (62). The functional gut-associated lymphoid tissue develops at day 4 posthatch (Fig. 1), whereas, the mature secretory IgA response develops by the second week of age in chicken gastrointestinal tissues (7).
By the time hatchery vaccination is performed, the ability of the immune system to mount antibody-mediated immune responses appears uncertain. For example, it has been shown recently that live attenuated vaccine (Arkansas serotype of IBV) administered at day 1 of age to chickens induces significantly lower antibody responses (IgY, IgA, IgM isotypes) when compared with chickens vaccinated on day 7 with less IgG avidity and less protection (120). However, maternal antibodies transferred through the yolk sac to developing embryo can persist for a few weeks posthatch, providing transient protection (124), especially when the protection correlates with the level of maternal antibody response. Maternal antibodies are found throughout the incubation period in the yolk sac and in the embryo serum after ED12. The levels of maternal antibodies rapidly decline during the posthatch period (22), but not sooner than the bird acquires the capability to mount antibody-mediated immune response following hatchery vaccination.
Recently, Gimeno et al. (41) have shown that in ovo vaccination with HVT ameliorates the limitation of potentially underperforming immune system by increasing antibody-mediated immune responses to experimental antigens. As has been shown using experimental antigens, such as sheep red blood cells and bovine serum albumin, prehatch in ovo and day 1 posthatch immunization increases the birds ability to form germinal centers in spleen earlier (4 days of age) when compared with unimmunized controls (>10 days of age) (117).
Cell-mediated immune response is also a primary function of the adaptive immune system and critical in mounting vaccine-mediated immune responses. The precursor T cells originated from bone marrow enter the thymus of the embryo in three waves; the first wave at ED6, the second wave at ED12, and the third wave at ED18 (15). However, the differentiated mature T cells will migrate to peripheral tissues continuously for many weeks posthatch and are observed in gut-associated lymphoid tissue by day 4 posthatch (101). However, the antigen-presenting molecule, major histocompatibility complex (MHC) I, is detectable between ED6 and ED7 in thymus, bursa of Fabricius, and spleen (26) and antigen-presenting cells, such as DCs and macrophages are available early during the embryo development (36,81,105,127).
Nevertheless, recently it has been shown that hatchery vaccination performed at ED18 with HVT vaccine against MD is capable of increasing T cell subsets, T cell proliferation, and antigen-presenting molecules, such as MHC I and II (41) to compensate for the lack of generation of cell-mediated immune responses around hatch. However, it remains to be seen whether other hatchery vaccines are also capable of inducing maturation of adaptive arm of the immune system as has been observed for in ovo HVT vaccination.
Mechanisms of Hatchery Vaccine-Mediated Immune Response
It is known that administration of HVT vaccine in ovo at ED18 does not induce tolerance (132) and this may be the case potentially during early posthatch period. Given the developmental stage of chicken immune system and the ability of the immune system to elicit adaptive immune responses around the time hatchery vaccination is performed, it is difficult to explain the success of hatchery vaccination in terms of protecting chickens against various diseases. The available evidence suggests potential roles of innate immune system in addition to vaccine-mediated maturation of adaptive arm of the immune system as potential mechanisms of protection mediated by hatchery vaccines.
First, given the fact that adaptive immune system is still naive to antigens when they are vaccinated prehatch and early posthatch (74,76,120), it is possible that innate arm of the immune system plays a major role in the vaccine-mediated protection (Fig. 2). When in ovo hatchery vaccination is performed at ED18 prehatch or day 1 posthatch, innate arm of the immune system plays a crucial role to protect the chicks during early posthatch period (92, 97). This protection in terms of reduction in viral replication, morbidity, and mortality following posthatch challenge with pathogenic viruses have been shown to be associated with the enhanced macrophage response, increased NK cells activity, secretion of proinflammatory mediators, such as type 1 IFNs, and increased production of an innate antiviral molecule, nitric oxide using TLR ligands delivered in ovo (43,44,49,100,104,111,112).
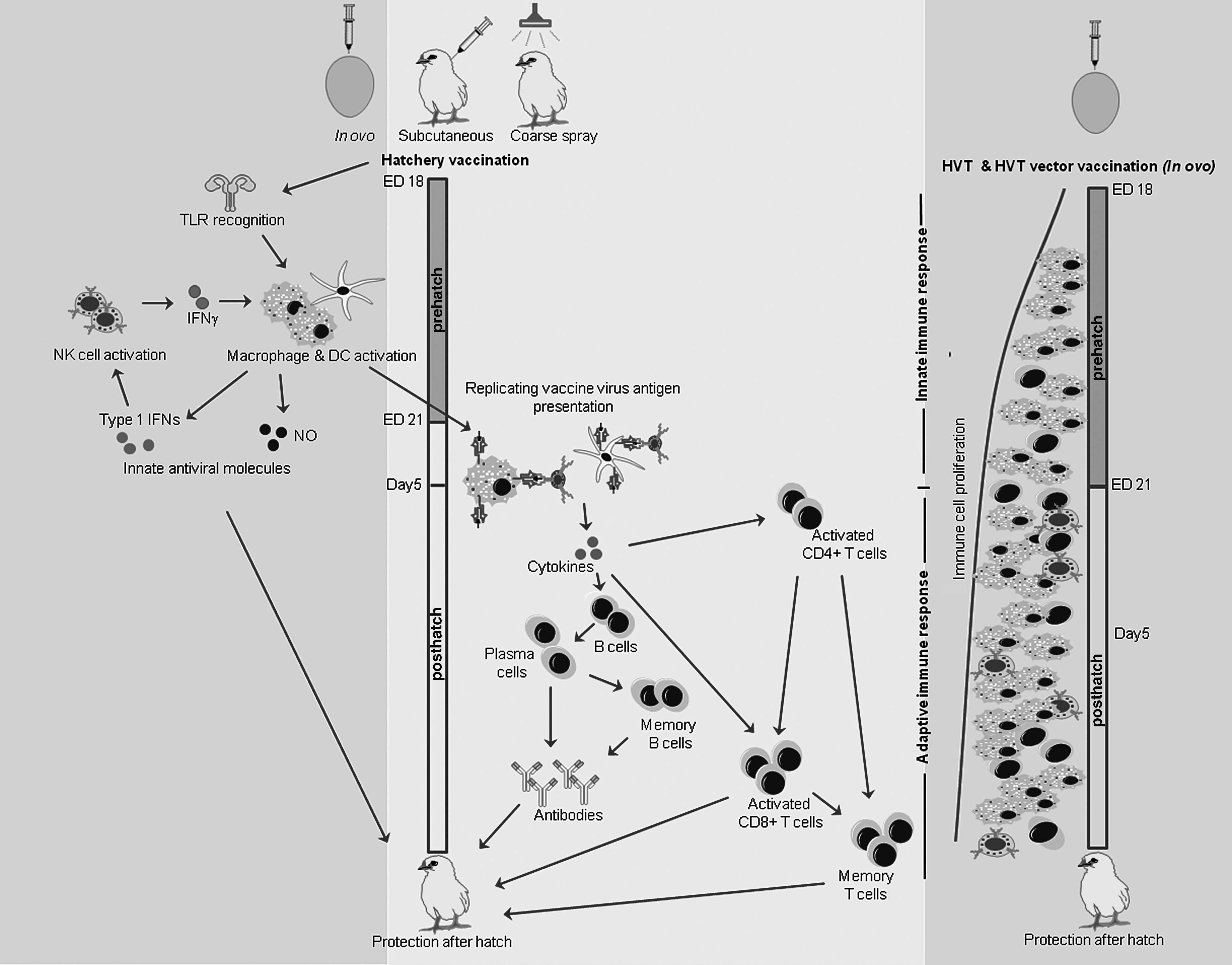
Induction of innate and adaptive host responses following hatchery vaccination. Initially, innate immune system plays a major role in the vaccine-mediated protection following hatchery vaccination. The recognition of PAMPs of vaccine virus by TLRs eventually will stimulate the immune cells, including DCs, macrophages, and NK cells for their maturation, differentiation, and expansion. Antigen presentation by DCs and macrophages along with the cytokine milieu resulting from innate host responses potentially leads to vaccine-mediated adaptive immune response during the early posthatch period. Immune response time line for HVT and HVT vector vaccination is shown separately (41). In ovo-delivered HVT vaccine is capable of increasing T cell subsets, T cell proliferation, and antigen-presenting molecules, such as MHC I and II in the spleen of chickens from day of hatch. HVT, herpesvirus of turkeys; NO, nitric oxide; PAMP, pathogen-associated molecular pattern; MHC, major histocompatibility complex.
To induce innate host responses until components of adaptive immune system is capable of eliciting adaptive immune responses, innate receptors should be engaged by the vaccines by making available PAMPs until such period. This is made possible by administration of replicating viruses as vaccines or delivering vaccine antigens using slow release delivery mechanisms. Binding of PAMPs results in conformational changes in the cytoplasmic signaling domain of TLRs and consequently recruitment of adaptor molecules takes place to initiate downstream signaling (65,67).
The initiated signaling from TLRs associated myeloid differentiation primary response-88 molecules, will pass through the activation of many cytoplasmic mediators, such as interleukin 1 (IL-1) receptor-associated kinase 4, IFN regulatory factor 5, and tumor necrosis factor receptor-associated factor 6 and ultimately leading to the activation of nuclear factor kappa (NF-K) B or activator protein (AP) 1 (5,125). NF-KB or AP1 in turn upregulates the gene transcription of proinflammatory mediators in the nucleus (6,125). The PAMPs-TLRs immunostimulatory pathways eventually stimulate the immune cells, including DCs, macrophages, and NK cells for their maturation, differentiation, and expansion. Although NK cell activity has not been observed in the context of hatchery vaccination performed prehatch or day 1 posthatch, increase of NK cell cytotoxicity 4–14 postvaccination following day 6 vaccination with HVT has been documented (49,99).
The literature on macrophage recruitment pre- and posthatch following hatchery vaccination are scarce, although increase in macrophage numbers in lungs has been observed following induction of innate immune response using in ovo-delivered TLR ligands (47,111,112). There are no data to support that DCs are recruited following hatchery vaccination; however, the fact that DC precursor cells colonize lymphoid organs before being colonized by lymphoid precursor cells (81) suggest a potential role of DCs in hatchery vaccination. Chicken DCs express TLRs as such are capable of recognizing PAMPs derived from replication of vaccine viruses (128). Chicken DCs treated with IBDV antigens or TLR ligands have been shown to increase CD40 and CD86 costimulatory molecules increasing splenocyte proliferation (72,73). Avian DCs are also known to increase B cell proliferation increasing antibody production (24).
Since DCs and macrophages are the antigen-presenting cells, it is possible that these two cell types along with the cytokine milieu (129) resulting from innate host responses potentially mediate adaptive immune response posthatch as primary and secondary antibody response following hatchery vaccination in chicken has been observed (104,123).
Second, it is possible that vaccination process may increase the ability of the immune system to respond to vaccine antigens. Abdul-Careem et al. compared in ovo-delivered and D14-injected HVT vaccine replication and found that in ovo-delivered HVT replicates better (3). This may be because of the fact that early splenocytes are more permissive to vaccine viral replication compared with adult chicken splenocytes and age-related differences in host responses may also contribute to different patterns of vaccine viral replication observed (3). The same study demonstrated that in ovo immunization of HVT increases IFNγ and IL-10, although constitutive levels of cytokine gene expressions are lower prehatch (2). Recently, it has been shown that in ovo-delivered HVT vaccine is capable of increasing T cell subsets, T cell proliferation, and antigen-presenting molecules, such as MHC I and II in the spleen of chickens (Fig. 2) (41). This response mediated by HVT hatchery vaccination also has been shown to increase antibody-mediated immune response to experimental antigens (41). The formation of germinal centers in secondary lymphoid organs is critical in the elicitation of antibody-mediated immune responses. When compared with unimmunized controls, it has been shown that administration of experimental antigens in ovo or day 1 posthatch leads to the formation of germinal centers 10 days earlier around 4 days of age (117).
However, the persistence of vaccine antigens until adaptive immune system is ready to process antigens is important and that may be a potential reason why only replicating viruses are commonly used as hatchery vaccines. Induction of innate immune responses mounted immediately following hatchery vaccination may be capable of dampening the vaccine viral replication rather than elimination, potentially allowing the persistence of vaccine antigens longer. Previously, it has been shown that the induction of innate host response through in ovo delivery reduces the viral replication and not the complete elimination (111,112).
Limitations of Hatchery Vaccination
Variable efficacies
Of the vaccines currently licensed for hatchery use, only a portion are efficacious. In ovo vaccination has been practiced for the control of MD, ILT, ND, AI, and IBD using either live virus, live attenuated virus, or recombinant viral vectors. The live and live attenuated vaccines used against MD are very efficacious when used pre- or posthatch at the hatchery. Before the introduction of vaccines in the early 70s, the poultry industry experienced great losses due to MD-associated paralysis, high morbidity, and mortality. The use of MD virus vaccine has significantly reduced the number of culls, condemnations, and mortality in vaccinated flocks (58). The introduction of in ovo vaccination against MD has increased the economic benefit to the producers when compared with the conventional day-old administration of the vaccine (96).
Currently, there are three licensed recombinant vaccines for ILTV, two using a HVT and one using a fowlpox virus as vectors. These vaccines are more commonly used in broiler chickens than the live-modified ILTV vaccines. Interestingly, the immunity observed in broiler chickens vaccinated with recombinant HVT-ILT vaccine confers protection against respiratory clinical signs, but not against viral shedding (57,116). The immunity conferred by this vaccine requires more time to build when compared with recombinant fowlpox-ILT vaccine and with live-modified ILTV; therefore, young birds may not be fully protected in the event of an ILTV challenge (57,116). Although recombinant fowlpox-ILT vaccine generates ILTV immunity earlier than recombinant HVT-ILT vaccines, recombinant HVT-ILT vaccines are able to have a statistically significant reduction of clinical signs in terms of depression, respiratory difficulties, or conjunctivitis in older broiler birds when vaccinated by the in ovo route (57,116). Both these vector ILT vaccines elicit partial protection when compared with live attenuated ILT vaccines (116).
Currently there are two HVT and fowlpox-vectored AI (H5) vaccines and there are no efficacy data available for these two hatchery vaccines (108). It has also been shown that HVT-vectored ND vaccine when delivered in ovo or day 1 posthatch requires 4 weeks to elicit full protection and 3 weeks to elicit partial protection (29) indicating that birds are not protected by the time they are placed in the barn environment. In ovo delivery of HVT-vectored IBDV vaccine or injection of live attenuated IBD vaccine are unable to prevent disease caused by a variant IBD isolated in Western Canada infected at day 6 posthatch (66). Contrastingly, a Korean study shows that in ovo-delivered HVT-vectored IBD vaccine is protective when challenged at day 1 posthatch with minimal bursa of Fabricus damage (95) suggesting that the efficacy may be dependent on the time and virulence of the challenge. Altogether, live and live attenuated vaccines are more efficacious as hatchery vaccines when compared with viral vectored vaccines.
Maternal antibody interference
The presence of high level of maternal antibodies in the vaccinated host may neutralize or interfere with the replication of live vaccine strains, therefore, vaccine-mediated immunity could potentially be reduced. There are few observations that suggest that some hatchery vaccines could be neutralized by maternal antibodies in the host. For example, it has been shown that although a single dose of live ND vaccine is sufficient in providing adequate protection in antibody-negative chickens, multiple vaccinations are required to provide a similar level of protection in chickens with potential maternal antibody interference (8). Although, information are lacking on maternal antibody interference of IB vaccines, the fact that maternal antibody against IBV could protect against virulent challenge encountered day 1 and not day 7 (79) suggests that maternal antibodies potentially neutralize IB coarse spray administered day 1 posthatch.
Commercial IBD live vaccines are available as mild, intermediate, and intermediate plus strains of serotype 1 IBDV. Mild or intermediate vaccines are used in parent stocks to prime the immune system before boosting with an inactivated vaccine (17) with a view of generating maternal antibody levels in the progeny. However, the same vaccines are not effective if administered to progeny with high levels of maternally derived antibodies, suggesting maternal antibody interference with the effectiveness of the vaccine (9,80,118). Nevertheless, this can be avoided by administering the vaccine after maternal antibodies have declined to a level that does not interfere with neither vaccination nor protection mediated by maternal antibodies (9,17) or choosing appropriate vaccine strains based on maternal antibody levels (97). The use of highly pathogenic vaccine strains (intermediate or intermediate plus strains) to combat interference with maternally derived antibody had been described, although it may increase the risk of development of subclinical disease and subsequent immunosuppression (46,77,80,82,118).
HVT- and fowlpox-vectored ILT, ND, AI, and IBD vaccines are less vulnerable to maternal antibody interference since HVT-vectored vaccines are cell associated (27) and the clearance of fowlpox-vectored vaccines is dependent on cell-mediated rather than antibody-mediated immune responses (34). However, it has been shown that maternally derived antibodies against AIV (90) and ILTV (131) could potentially interfere with fowl poxvirus-vectored vaccines. HVT-vectored IBD vaccines appear to be less affected by the presence of maternally derived antibodies (68). Recently, it has been shown that day 1 hatchery vaccination with NCD-vectored ILT vaccines does not interfere with maternal antibodies present against NCDV and ILTV (131).
Immunosuppression
During immunosuppression, antibody- and cell-mediated immune responses will be affected; therefore, vaccine-mediated adaptive immune response will not be adequate to protect the birds when a disease outbreak occurs. Many factors may lead to immunosuppression in birds, including stress due to poor nutrition or poor environmental conditions, concurrent viral infection, such as MD and IBD, or consumption of feeds with mycotoxin contamination.
Hatchery vaccination with some vaccines also have been shown to be immunosuppressive. For example, certain vaccines such as IBD and reovirus vaccines can negatively influence the developing immune system in late embryonic stage as has been shown by in ovo vaccination trials conducted using reovirus (45) and IBD vaccines (20,56). The in ovo delivery of an immune complex vaccine against IBD leads to transient B cell depletion and bursa of Fabricius atrophy in birds, which are reversible with or without maternal antibody interferences (53). Immunosuppression as observed by decreased B cell functions, such as proliferating ability, antibody production, and increased susceptibility to secondary bacterial infections, has been demonstrated following hatchery vaccination at the day of hatch using bivalent HVT + SB1 or Rispens vaccines against MD (35).
Of the vaccines administered at the hatchery, live HVT vaccine and HVT-vectored vaccines are commonly used for the control of MD, ILT, ND, IBD, and AI. HVT vaccine itself does not induce immunosuppression (33,52), except in rare occasions (35). HVT-vectored IBD vaccine is not immunosuppressive when delivered on the day of hatch (87). However, recently, it has been shown that HVT-vectored IBD vaccine delivered in ovo could increase the pathogenesis of IBD characterized by immunosuppression when compared with the live attenuated IBD vaccines (66).
Conclusions
Currently, hatchery vaccination in poultry depends on live, live attenuated, and viral vectored vaccines. Hatchery vaccines are available against major economically important viral diseases in chickens (MD, ILT, IB, IBD, ND, and AI) to be administered as either in ovo, coarse spray, or subcutaneous vaccines around the hatch within a short window of time before the day-old chickens are being placed in potentially contaminated barn environments.
There is a dearth of information in the area of potential mechanisms of protection mediated by these hatchery vaccines (3,41). The comparison of the efficacies of these hatchery vaccines show that the viral vectored vaccines are less efficacious and research is needed to identify ways of improving the efficacy of viral vectored vaccines as useful hatchery vaccines. It is also important to investigate alternatives that ameliorate potential maternal antibody interference and immunosuppression observed with some hatchery vaccines. When multiple vaccines are administered around hatch and within a short time period, it is possible that there may be positive (69) or negative interactions, and investigations are necessary to identify such interactions to improve hatchery vaccination practices for the benefit of poultry industry.
Footnotes
Acknowledgments
The graduate studies of M.S.A. and U.D.S.S. are supported by the Canadian Poultry Research Council, Alberta Agriculture and Forestry and Natural Sciences and Engineering Council (NSERC) of Canada grants awarded to M.F.A. The graduate study of V.P. is supported by grants from MITACS and Institute of Poultry Technologies, Alberta, Canada. The studies of A.A. are funded by Saskatchewan Agriculture, Canada. H.A. received a scholarship from the Egyptian government to visit University of Calgary to conduct research.
Author Disclosure Statement
No competing financial interests exist.