
Abstract
Sow immunity plays an important role in preventing viral infection and disease in newborn piglets. Vertical transmission of porcine circovirus type 2 (PCV2) may perpetuate porcine circovirus associated disease (PCVAD) in newborn and growing pigs. Hence, the immunological effects of maternal immunoglobulin transfer of PCV2-specific antibodies on PCV2 viremia and immune response in piglets in commercial swine herds were evaluated. Sow vaccination has been shown to reduce viral shedding and viremia, and increases the neutralizing antibody (NA) titers. Since NAs are important for control of PCVAD and mammary secretions may contain high anti-PCV2 NA levels, we examined the PCV2 NA levels in colostrum, milk, sow serum, and piglet serum over time to investigate an association between NA levels and protection against infection. NA titers were remarkably high (up to 10−6 50% neutralizing titer) in sow serum and colostrum on all farms regardless of viremia levels. In piglets vaccinated at 3 weeks of age, NA titers peaked at 10 weeks of age and continued to maintain high viral neutralizing titers to slaughter. The impact of maternally derived neutralizing activity was most evident during the suckling period. Although PCV2 was transmitted from sows to piglets in colostrum, piglets were largely nonviremic at weaning. Thus, NAs appear to control or suppress initial infection even though they are unable to clear or prevent infection later in life.
Introduction
P
PCV2 NAs are important for control of PCVAD, since negative or low titers of NAs correlate with disease and high levels of viral replication (7,21). Also, the appearance of NAs and interferon (IFN)γ-secreting cells has been correlated with a reduction in viremia (16,22,34). The immune response to vaccination and natural infection differs in that vaccination induces NAs more so than infection, which induces high levels of non-NAs, suggesting that the PCV2-specific antibody response does not predict the protective immune response (36,38). Vaccination of sows or piglets, or both sows and piglets, significantly reduced viremia and induced higher NA titers and a higher proportion of CD4+CD8+IFNγ + cells compared to nonvaccinated animals (6,22).
Both colostrum and milk may have high NA titers, as well as infectious virus; thus both virus and NA are passed onto piglets during nursing, but remain protected against disease (11,12). High levels of maternal NAs and PCV2-specific antibodies are observed in piglet serum in the first few months of life and they decrease over time (11,12). In vaccinated animals, PCV2 NAs were observed in 25% of pigs at 2 weeks postvaccination and by 4 weeks, 100% of animals were positive for NAs, which were maintained for at least 8 weeks (15).
In this study, we developed a sensitive NA assay to identify the level of PCV2-specific NAs present in sow serum, piglet serum, colostrum, and milk samples to determine levels and temporal kinetics of NA transfer to piglets and the effect on viremia. The highest NA titers were observed in colostrum and titers decreased over time in both milk and piglet serum. NA titers peaked at 10 weeks of age, decreasing out to 24 weeks, but continue to maintain high titers. Overall, NAs are present in high titers in both mammary secretions and serum.
Materials and Methods
Virus, cells, and samples
An isolate of PCV2b, viral strain 321 (Genbank ID JF290418), was used for all NA assays. VR1BL cells, kindly provided by Dr. Suresh Tikoo (VIDO, Saskatchewan, Canada), were grown in minimal essential medium (MEM) media supplemented with 1 × non-essential amino acids, 10 mM HEPES, 50 μg/mL gentamicin, 50 μg/mL hygromycin B, and 10% fetal bovine serum (4). Serum, colostrum, and milk samples were obtained from commercial swine farms from archived samples that were provided for various studies.
PCV2 viral copy number and antibody titer
Porcine umbilical cord (PUC) blood, PUC swabs, piglet serum, and colostrum from PCV2 stable and unstable sow herds were analyzed for PCV2 virus presence by PCR at the Health Management Center (Boehringer Ingelheim, Ames, IA). Prefarrow sow serum samples from PCV2-stable and PCV2-unstable farms were analyzed by enzyme linked immunosorbent assay (ELISA) at the Iowa State Veterinary Diagnostic Laboratory (Ames, IA). Stable and unstable are terms used to classify sow herds as previously exposed (i.e., show widespread presence of antibodies in serum) in combination with absence or near absence of viremia (stable) or presence of viremia (unstable) as determined by PCR. The terminology was first applied to porcine reproductive and respiratory syndrome virus status of sow herds and has been widely adopted for other swine diseases (14). In this study, PCV2-stable farms were identified as having 1% or less PCV2-positive sows, and PCV2-unstable farms showed ∼50% of sows as PCV2 positive by PUC testing.
Samples of colostrum, sow serum, and piglet serum were analyzed for PCV2 antigen by isolating total DNA from serum and colostrum using the QIAamp DNA blood mini kit (Qiagen, Valencia, CA), following the manufacturer's protocols as described previously (4). Viral copy number was determined using a quantitative PCV2 PCR assay as described previously (4). PCR was a more reliable estimator of viremia than was classical growth in cell culture, since cell culture permissiveness was highly variable (4). PCV2 capsid-specific antibodies were determined using an ELISA as described previously (3).
PCV2-neutralizing antibodies
VR1BL cells were seeded on polyethylinimine-coated 96-well cell culture plates at 3 × 104 cells per well and incubated for 1.5 days until semiconfluent monolayers were formed (60–90% confluent). Colostrum and milk samples were prepared by centrifugation at 13,000 g for 15 min, diluting the aqueous fraction of colostrum samples 1:32 in MEM media, and filtering both sample types through a 0.22 μm filter (Millex-GV; EMD Millipore Corp., Billerica, MA). Serum, colostrum, and milk samples were heat inactivated for 30 min at 56°C and serially diluted fourfold starting at 1:2 or 1:8 for serum and milk, and 1:32 for colostrum. Colostrum samples with out-of-range titers were reanalyzed with a 1:5 serial dilution, starting at 1:50. Diluted samples were incubated with an equal volume of PCV2b virus (∼5 × 104 tissue culture infectious dose 50% to give a multiplicity of infection [MOI] of 0.5 per well) and incubated for 1 h at 37°C, 5% CO2. The sample-virus mixture (100 μL) was then transferred to the semiconfluent VR1BL cell monolayers and incubated for 2 h at 37°C, 5% CO2. The inoculum was removed, cells were washed twice with MEM media, 200 μL of VR1BL complete media was added, and cells were incubated for 48 h at 37°C, 5% CO2. Media were then removed, cells were washed with phosphate-buffered saline, and fixed for 10 min at −20°C using ice-cold methanol. All samples were analyzed in duplicate or multiple replicates. Controls on each plate included cells only, cells and virus only, cells with virus and positive control serum, and cells with no virus plus test sample.
Plates were stored at −20°C until ready for staining and analysis. Infected cells were visualized by immunofluorescence analysis using a PCV2 capsid-specific monoclonal antibody (Rural Technologies, Brookings, SD) and an AlexaFluor 488 goat anti-mouse immunoglobulin G (IgG) secondary antibody (Invitrogen, Carlsbad, CA). Nuclei were stained with bisbenzimide (Sigma-Aldrich, St. Louis, MO). Stained monolayers were imaged on either an Olympus IX70 inverted fluorescence microscope (Olympus America, Inc., Center Valley, PA) or a Nikon A1 confocal microscope at the University of Minnesota CBS Imaging Center using SPOT basic imaging software (SPOT Imaging Solutions, Sterling Heights, MI) or NIS-elements AR software (V 4.30.02; Nikon Instruments, Inc., Melville, NY). Infected cells in the images were counted using ImageJ software. Data analysis was based on one image per well, containing ∼1,500 cells, of which ∼30% were infected in the absence of NAs (33). Fifty percent NA titers were calculated by plotting the curve corresponding to the number of infected cells at each dilution and calculating the dilution that corresponded to a 50% reduction in infected cells in the test wells containing virus only. Ninety percent NA titers were also determined during test validation, but 50% titers were routinely calculated to provide information about virus–antibody interactions under equilibrium binding conditions, as described previously (30,35). The 50% titer values also appeared to be less variable, with sample differences being more pronounced, as was observed previously (7).
Results
Optimization and validation of NA assay
The neutralizing assay was based on previously developed assays for porcine delta coronavirus and porcine reproductive and respiratory syndrome virus neutralization, and optimized for PCV2 and its permissive VR1BL cell line, which is 40 times more permissive than PK-15 cells for viral growth, to allow for increased sensitivity and dynamic range (4,30,35). The assay was optimized for input virus (MOI 0.5), culture duration (48 h), and sample dilution using multiple serum, milk, and colostrum samples. Optimal sample dilution curves were determined to consist of fourfold serial dilutions for serum and milk starting at 1:2 or 1:8, and at 1:32 for colostrum. Further dilutions of colostrum starting at 1:50 with fivefold serial dilutions were necessary for samples with exceptionally high titers. NA titers were observed from <1:4 to >1:106, showing a much greater dynamic range than had been observed in previous NA assays (6,7,26).
Neutralization of differing amounts of virus with dilutions of NAs was examined to determine if PCV2 neutralization followed the percentage law as well as to determine the optimal MOI for routine testing (1,2). The percentage law basically states that a given amount of antibody neutralizes virus at a constant percentage, regardless of the amount of virus present (1,2). We examined the percent of inhibition of viral infection at various MOI's due to the addition of NAs from various samples at various dilutions (Fig. 1, colostrum sample results). We determined that the antibodies neutralized virus infection at all MOIs and the 50% neutralizing titers decreased with decreasing amounts of input virus (Fig. 1). The percentage law was valid in the range of 0.1 to 1.0 MOI. At an MOI >1.0, the virus/antibody ratio, rather than antibody concentration alone, determined the 50% neutralizing titer. At lower dilutions, the 50% neutralizing titer was highly variable due to the low number of positive infected cells in a well, even in the absence of NAs. Thus, an MOI of 0.5 was used routinely for titer determinations.
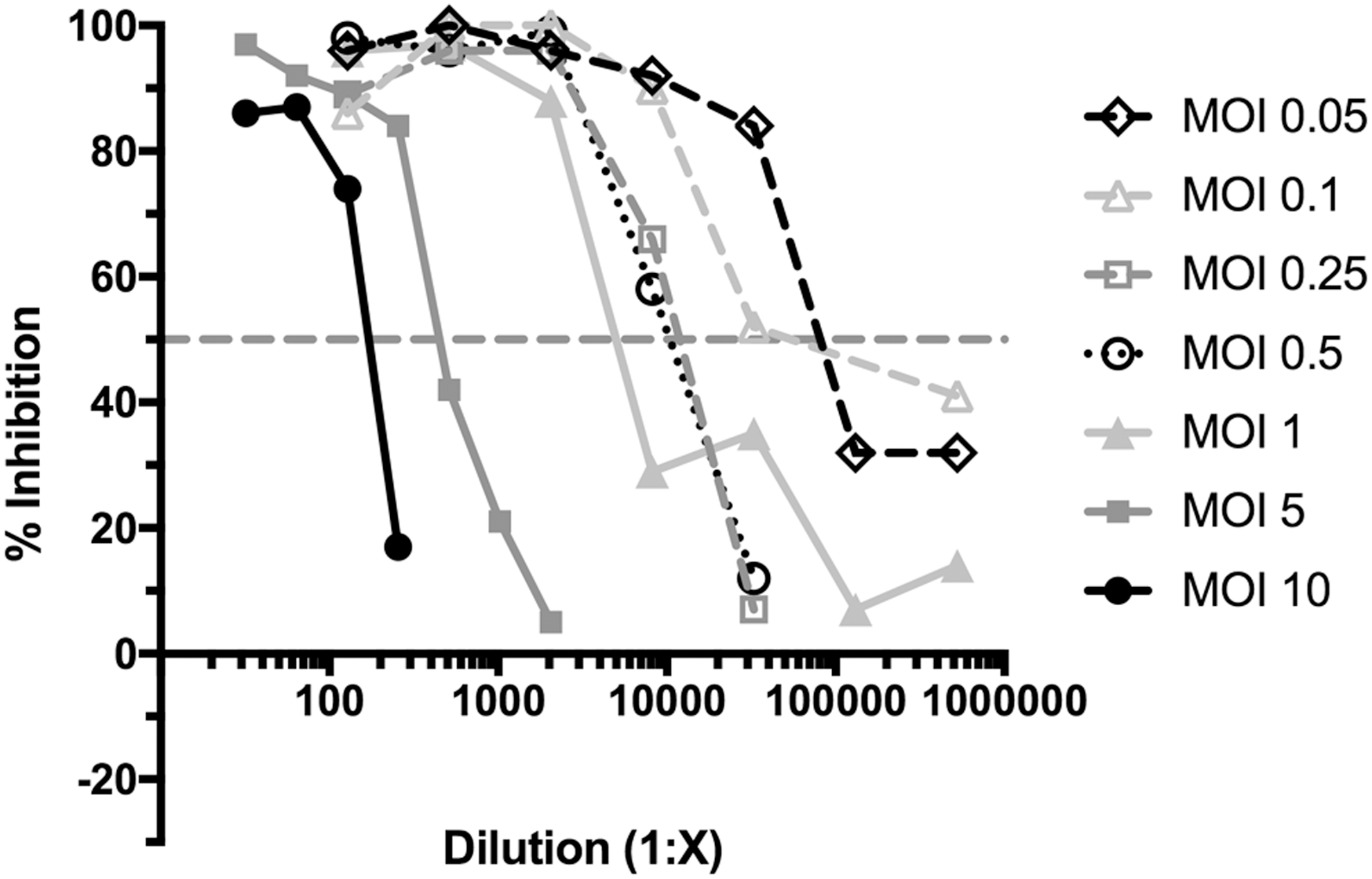
Validation of the NA assay. The NA assay was run on multiple samples of serum, colostrum, and milk. Data from a single colostrum sample that contained a high titer of NAs are shown in this study as an example of results. The colostrum sample was serially diluted twofold for a final dilution ranging from 1:32 to 1:2,048 for MOIs of 5 and 10, and fourfold serially diluted from 1:128 to 1:8,388,608 for MOIs 0.05 through 1. The colostrum dilutions were mixed with differing amounts of virus before adding to VR1BL cells for 2 days and then processed to enumerate infection. The percent inhibition measured the percentage of virus that was neutralized by the antibodies present in the colostrum sample over the dilution series. NA, neutralizing antibody; MOI, multiplicity of infection.
PCV2 status and NA levels
In this experiment, four commercial pig farms were examined for the presence of PCV2 antigen, antibodies to PCV2, and NA levels to determine if NA titer correlated with the PCV2 status of the farm. Two farms (Farm 1 and 2) were reported to have few or no sows that were PCV2 virus positive and were considered PCV2 stable (Fig. 2A). By contrast, on the other two farms (Farms 3 and 4), ∼50% of sows were PCV2 positive, although the virus was present at low levels, and so the farms were considered unstable (Fig. 2A). Results were recorded as positive or negative since positive tests were consistently near the limit of detection. Analysis of total anti-PCV2 antibodies by capsid ELISA showed that antibody levels varied between farms, but did not correlate with viral presence (Fig. 2B). Examination of the PCV2 NA titers in sow colostrum from these farms showed higher PCV2 NA titers in the PCV2-stable farms with an average colostral 50% NA titer of 1:74,000 ± 13,000 (mean ± standard error of the mean) in animals from PCV2-stable farms and a significantly lower level (1:27,000 ± 5,700) in animals from PCV2-unstable farms (p < 0.01) (Fig. 2C). The 50% NA titers in colostrum ranged from 1:86 to 1.1 × 106. The findings showed that sows on stable farms with minimal PCV2 presence in serum have higher neutralizing titers in serum than sows on unstable farms.
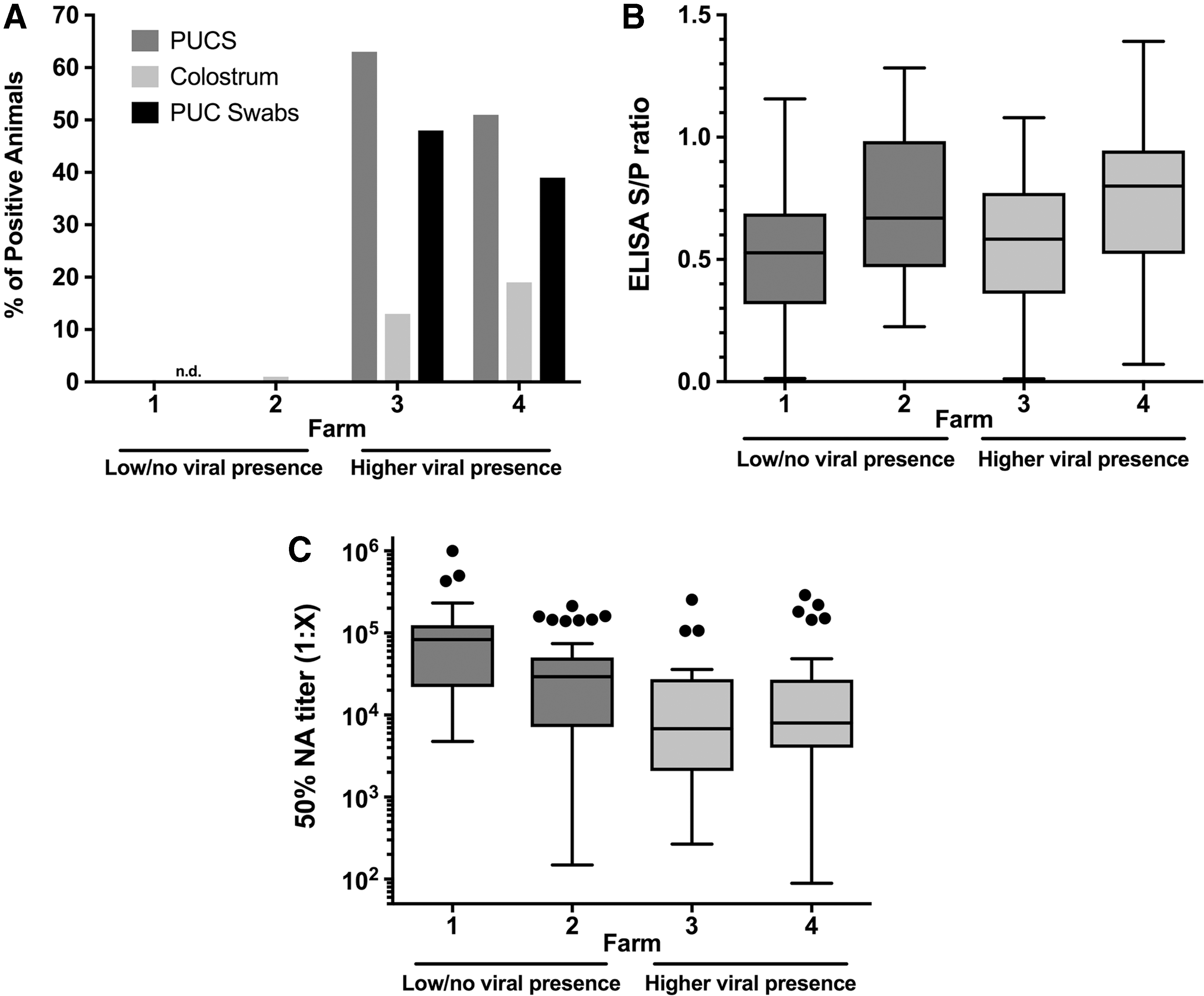
NA titers from PCV2-stable and PCV2-unstable farms.
Comparison of NAs in colostrum, milk, and serum
A second experiment was carried out to compare NA levels in serum, colostrum, and milk of sows compared to levels in the serum of their piglets. Sow serum was collected before farrowing and on the day of farrowing, and piglet serum was collected on days 1 or 2, 5, 12, and between days 17 and 19. Colostrum was obtained on the day of farrowing (day 1), and milk was collected on days 3, 12, and 19. In this experiment, colostrum gave the highest NA titers with an average 50% NA titer of 1:33,000, with the highest sample having a titer of 1:180,000 (Fig. 3). The mean colostrum titer was 2.7-fold higher than in sow serum before farrowing. Farrowing and the onset of nursing were associated with a large decline in serum antibody titers, presumably due to transfer into colostrum (32). Neutralizing titers in mammary secretions decreased over time, consistent with depletion of initial immunoglobulin stores, dilution due to increased milk output, and isotype change from primarily IgG in colostrum to immunoglobulin A (IgA) in milk (35). Neutralizing titers in piglet serum were highest following colostrum ingestion through day 5 and then declined (Fig. 3). Interestingly, the large range of NA titers of colostrum and early milk (day 3) appeared to partially translate into a broad titer range early in piglet life, but the variation in piglet serum was limited at all later times. A decline in piglet serum neutralizing titers over time was expected since passive transfer of maternal antibodies is limited to the period of colostrum production, about 36 h. Without further input before development of active antibody production, titers will decline as a function of increase in blood volume as piglets grow, and biochemical decay of immunoglobulins.
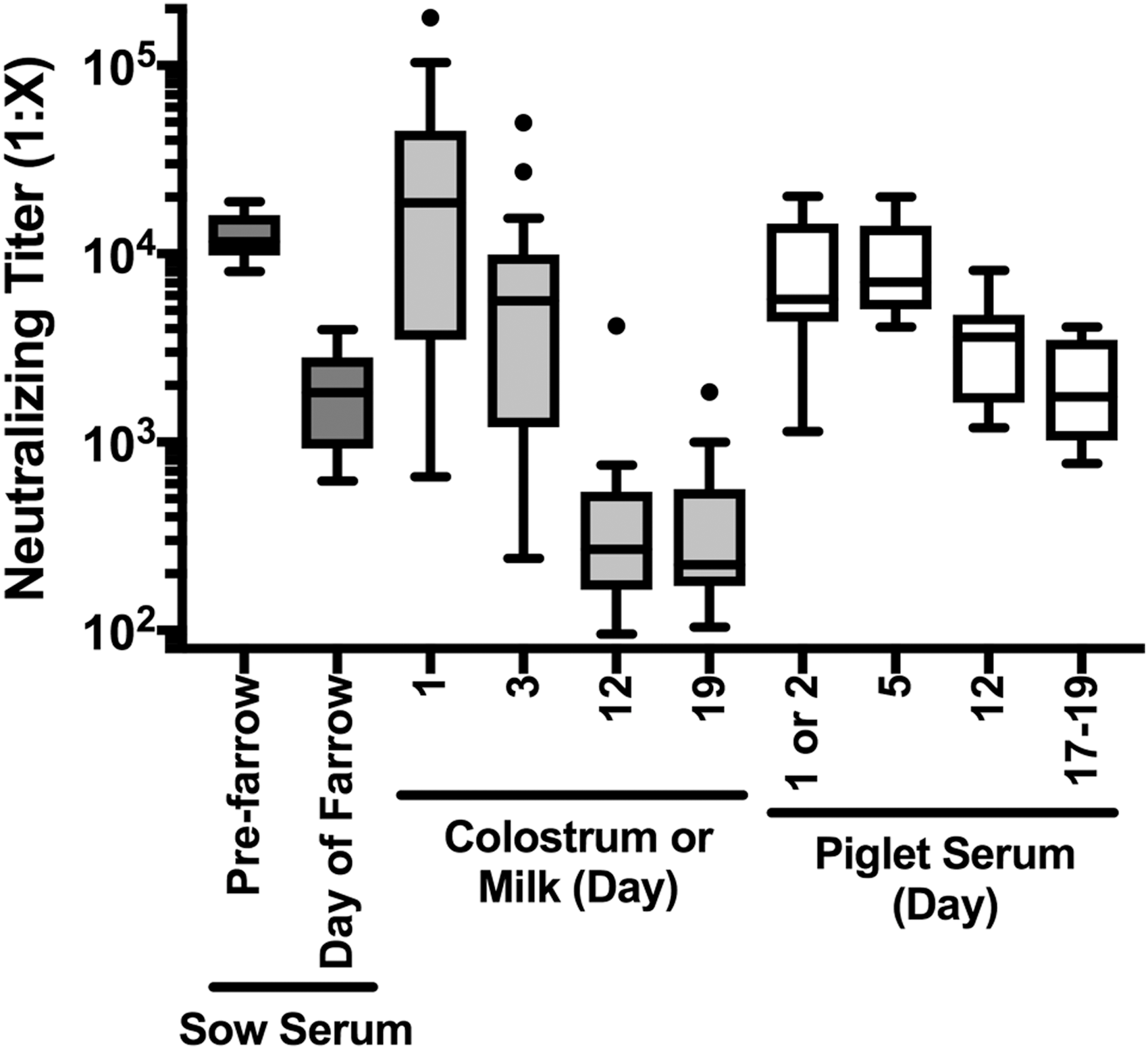
NA titers from sow and piglet serum, colostrum, and milk. Sow serum (n = 10), colostrum (n = 28), and milk (n = 20–28 per time point), and serum from their piglets (n = 6–10 per time point) were obtained from a commercial farm. NA titers were determined for all samples and are shown in a box-whisker plot using the Tukey determination of whiskers to show outliers. Day of farrowing is day 1.
Time course of NA titer development in growing pigs
To examine the induction of active antibody production in vaccinated, growing pigs, the time course of NA development from birth to slaughter was followed in pigs on a commercial farm that was positive for PCV2, but with no clinical signs of PCVAD in the herd. The findings were compared to total antibody levels determined by anti-capsid ELISA and to PCV2 viremia in the pigs over time and in the parental sows. As shown in Figure 4A, two of seven sows were viremic and six of seven shed PCV2 in colostrum. Conversely, the large majority (8 of 10) of piglets was PCR negative at birth and remained negative at weaning (3 weeks of age). By 10 weeks of age, all piglets were viremic with an average viremia level of 13,000 DNA copies per milliliter, which increased to about 50,000 copies per milliliter at 15 weeks. At 24 weeks of age, there was a wide range of viremia levels, both higher and lower than at 15 weeks. Clinical signs of PCVAD were not observed even though pigs were viremic.
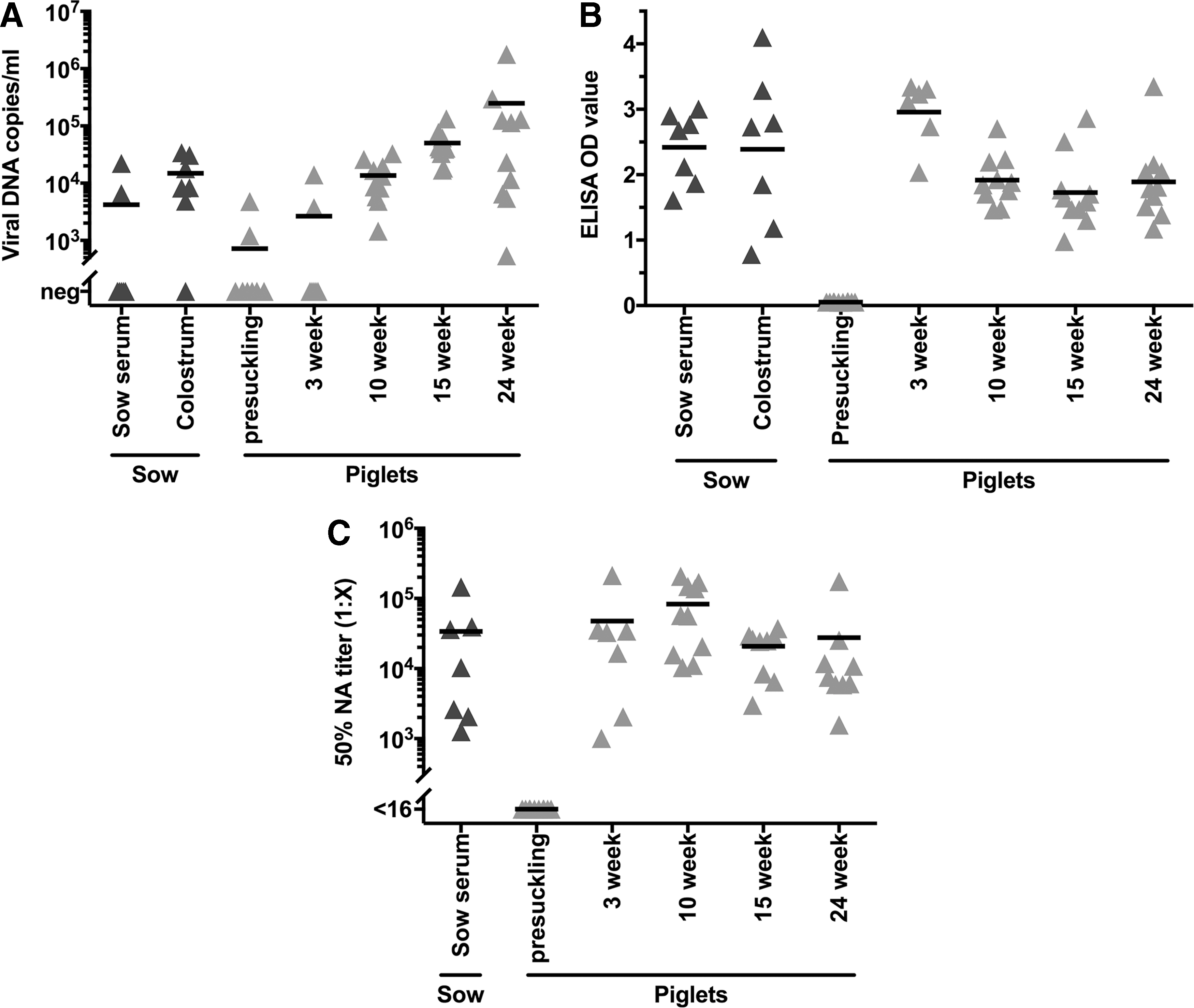
Sow and piglet viral and antibody levels over time. Sow serum, colostrum, and piglet serum samples were examined for viral DNA levels
All sows were seropositive for PCV2 capsid IgG antibodies and transferred antibodies to colostrum (Fig. 4B). Piglets, since they do not acquire maternal antibodies before birth, lacked anti-PCV2 antibodies before suckling, but had high levels at 3 weeks of age, and maintained high levels at later time points due to active antibody production. Anti-PCV2 antibodies were highest at 3 weeks of age, whereas slightly lower levels were maintained from 10 to 24 weeks of age. Specific NAs were present in all sows at an average titer >104 (Fig. 4C). Nursing piglet titers were equivalent to titers in parental sow serum. In contrast to ELISA-positive antibodies, neutralizing titers were consistently high throughout the growth phase to 24 weeks.
Average 50% NA titers from sow serum were 1:34,000 and average titers for piglet serum at 3, 10, 15, and 24 weeks were 1:48,000, 1:83,000, 1:21,000, and 1:28,000, respectively. NA titers in sow and 3-week-old piglet serum were correlated (r s = 0.81, p < 0.05). Viral DNA levels and 50% NA titers in all growing pigs, summed across all time points, were widely scattered and did not correlate.
Discussion
Maternal immunization increases the amount of antibodies available for passive transfer of antibodies from the sow to the piglets, increasing the protection against neonatal infections (32). Since piglets are born agammaglobulinemic, they are reliant on ingestion of colostrum for humoral protection. Even though pigs are immunocompetent at birth, it takes a period of weeks to mount a protective immune response to infectious challenge (32). Colostrum contains antibodies, cytokines, immune cells, and antigens, which are transferred to the newborn piglet during the first 24–36 h of suckling to protect against infection (32,35). Thus, if the sow is immune to infection, immunity will be transferred to the piglets, in addition to colostral nutrients and energy. However, passive transfer of specific antibodies may also interfere with immunization during the nursing period and delay development of an active immune response (32). For PCV2, the development of protective immunity has been shown with both sow and piglet vaccination, and maternal antibodies have not been shown to interfere with vaccine-based protection of piglets (10,23,24).
The NA assay developed in this study used a high infection dose, and highly permissive cells to PCV2 infection, which allowed for the optimization of key parameters of the assay and analysis of the percentage law. Analysis of the antibody reactivity independent of antigen concentration can be accomplished under equilibrium conditions that exist at 50% NA titer, but not at 90% NA titer (30). Neutralized and nonneutralized virus are in equilibrium at a 50% NA titer, thus allowing for mathematical analysis of the functional qualities of antibodies in the antisera following the law of mass action, as has been examined for other NA assays (17,18,29,30,37). More stringent neutralization results may be useful for diagnostic purposes or in cases of background cross-reactivity such as with dengue virus (31). However, interfering responses have not been observed in PCV2 neutralization assays (7,26).
Comparing PCV2-stable to PCV2-unstable farms showed that high NA titers were present on all farms, regardless of the frequency of PCV2-positive animals, and at much higher levels than had been observed previously (7,12,16,26). All farms were free of PCVAD despite a higher prevalence of PCV2-positive animals on the PCV2-unstable farms. Higher NA titers were observed in PCV2-stable farms, that is, with low prevalence of PCV2, suggesting that NAs may be involved in control or prevention of PCV2 infection at the herd level. All PCV2-positive animals had low viral loads and viral levels were not correlated with NA titers.
The relationship between NAs and viremia is not simple. Conflicting experimental results showed an inverse correlation or no correlation between viral load and NA titers, as we observed in this study (12,34). Similarly, the relationship between functional NA titers and total anticapsid antibody levels was not significantly associated in this study and elsewhere (34). It is well established that PCV2 infection occurs readily in the presence of robust humoral immunity in farmed pigs (3,5,12,27,28). The simultaneous presence of circulating antibodies, neutralizing activity, and PCV2 virus is perplexing and has not been satisfactorily answered. However, we show that sow colostrum provides a high level of NAs to piglet blood and piglets are largely nonviremic up to weaning. These observations indicate that maternally derived NAs may be preventing viremic infection by PCV2 that is present in colostrum.
Anti-PCV2 NA titers in sow serum were 6.4-fold higher before than on the day of farrowing. The marked decrease in serum activity is consistent with immunoglobulin translocation to mammary glands into colostrum (32). It has been determined that 100% of IgG, 85% of immunoglobulin M, and 40% of IgA in colostrum are obtained from serum, whereas milk antibodies are mainly synthesized locally in the mammary gland and are predominately IgA (∼85–90%) (32,35). NAs were highest in colostrum and lower in milk, especially at later time points. Piglet serum also had high levels of NA for the first few days and then decreased by fourfold to the end of the nursing period. Thus, piglets obtain a large dose of PCV2 NAs from colostrum that is absorbed into the bloodstream, and smaller amounts from milk, primarily in the form of IgA that is retained in the gut. The kinetics of antibody decline in the nursing period is variable among sows, although on average the decline is consistent with previous observations (35).
Interestingly, we observed that PCV2 viremia levels increased over time even though NAs were maintained at high levels in growing pigs. PCV2 capsid-specific antibodies peaked at 3 weeks of age, suggesting that vaccination at 3 weeks of age does not boost antibody levels. However, vaccination appears to be involved in increasing NA titers, as they increase after vaccination at 3 weeks until peaking at 10 weeks of age. It was previously shown that vaccination favored development of a neutralizing response, whereas infection elicited primarily non-NAs (36).
In conclusion, mammary secretions of PCV2-immune sows, both viremic and nonviremic, contained high levels of PCV2 NAs that were transferred to piglets during suckling and provide protection against viremic infection. Higher NA titers were observed in sows on stable farms with low PCV2 prevalence. NA titers increase after piglet vaccination and persisted in pigs throughout the first 6 months of life even though total anti-PCV2 antibodies declined. PCV2 NAs appear to be increased by vaccination of piglets and play a role in protection of growing pigs from PCVAD even though viral loads increase.
Footnotes
Acknowledgments
This work was carried out, in part, using software and resources available at the University Imaging Centers at the University of Minnesota. VR1BL cells were a kind gift from S.K. Tikoo (Vaccine and Infectious Disease Organization, University of Saskatchewan, Saskatoon, Canada). The authors would like to thank Zeynep Akkutay-Yoldar and Matilda Wagner for their help in performing neutralization assays for the serum, colostrum, and milk samples.
Author Disclosure Statement
The work was funded, in part, by support from Boehringer Ingelheim Vetmedica (BIV) and the USDA National Institute of Food and Agriculture, multistate project MIN-63-112. M.P.M. receives consulting income from BIV. The relationship has been reviewed and managed by the University of Minnesota in accordance with its conflict of interest policies.