
Abstract
Increased narcolepsy incidence was observed in Sweden following the 2009 influenza vaccination with Pandemrix®. A substitution of the 2009 nucleoprotein for the 1934 variant has been implicated in narcolepsy development. The aims were to determine (a) antibody levels toward wild-type A/H1N1-2009[A/California/04/2009(H1N1)] (NP-CA2009) and Pandemrix-[A/Puerto Rico/8/1934(H1N1)] (NP-PR1934) nucleoproteins in 43 patients and 64 age-matched controls; (b) antibody affinity in reciprocal competitive assays in 11 childhood narcolepsy patients compared with 21 age-matched controls; and (c) antibody levels toward wild-type A/H1N1-2009[A/California/04/2009(H1N1)] (H1N1 NS1), not a component of the Pandemrix vaccine. In vitro transcribed and translated 35S-methionine-labeled H1N1 influenza A virus proteins were used in radiobinding reciprocal competition assays to estimate antibody levels and affinity (Kd). Childhood patients had higher NP-CA2009 (p = 0.0339) and NP-PR1934 (p = 0.0246) antibody levels compared with age-matched controls. These childhood controls had lower NP-CA2009 (p = 0.0221) and NP-PR1934 (p = 0.00619) antibodies compared with controls 13 years or older. In contrast, in patients 13 years or older, the levels of NP-PR1934 (p = 0.279) and NP-CA2009 (p = 0.0644) antibodies did not differ from the older controls. Childhood antibody affinity (Kd) against NP-CA2009 was comparable between controls (68 ng/mL) and patients (74 ng/mL; p = 0.21) with NP-CA2009 and NP-PR1934 displacement (controls: 165 ng/mL; patients: 199 ng/mL; p = 0.48). In contrast, antibody affinity against NP-PR1934 was higher in controls with either NP-PR1934 (controls: 9 ng/mL; patients: 20 ng/mL; p = 0.0031) or NP-CA2009 (controls: 14 ng/mL; patients: 23 ng/mL; p = 0.0048). A/H1N1-NS1 antibodies were detected in 0/43 of the narcolepsy patients compared with 3/64 (4.7%) controls (p = 0.272). Similarly, none (0/11) of the childhood patients and 1/21 (4.8%) of the childhood controls had A/H1N1-NS1 antibodies. The higher antibody affinities against NP-PR1934 in controls suggest better protection against wild-type virus. In contrast, the reduced NP-PR1934 antibody affinities among childhood narcolepsy patients suggest poor protection from the wild-type A/H1N1 virus and possibly increased risk for viral damage.
Introduction
N
A strong genetic predisposition for narcolepsy type 1 with 75–98% of all patients associated with the human leukocyte antigen (HLA) allele DQB1*06:02 (22,33,46) suggests an autoimmune pathogenesis as an explanation for the highly specific hypocretin cell degeneration (38). A weak concordance rate of around 25% for monozygotic twins for narcolepsy type 1 indicates an important role for environmental stimulus (32). Infectious agents, including streptococcus (5,6) and H1N1 influenza A virus (15,48), have been suggested as triggers.
Increased incidence of narcolepsy was initially associated with Pandemrix® vaccination following the 2009 influenza pandemic not only in Finland (37) and Sweden (40,45) but later also reported from other European countries (8,18,34). In contrast, the incidence of narcolepsy in China was increased in conjunction with the 2009 H1N1 pandemic (14,15). It was suggested that idiopathic narcolepsy differs from vaccine-associated narcolepsy (45) and that there are differences in predisposing HLA genes before, compared with after, the 2009 influenza pandemic (13).
In a previous investigation, we observed that levels of A/H1N1 2009 HA (also present in the vaccine) antibodies were increased among the childhood patients compared with age-matched controls (27). In the present study, we wanted to test levels of antibodies against nucleoproteins and nonstructural proteins, which were either present or absent in Pandemrix.
It is hypothesized that the humoral immune response toward influenza nucleo- and nonstructural proteins differs among vaccine-associated narcolepsy patients compared with vaccinated healthy subjects. The aims were to determine (a) antibody levels toward wild-type A/H1N1 2009 nucleoprotein [A/California/04/2009(H1N1)] and Pandemrix nucleoprotein [A/Puerto Rico/8/1934(H1N1)] in 43 patients and 64 age-matched controls; (b) antibody affinity and specificity in childhood patients with narcolepsy (n = 11) compared with childhood controls (n = 21) in a reciprocal competitive assay by testing the binding of 35S-NP1934 and 35S-NP2009 with both nonlabeled NP-PR1934 and NP-CA2009 proteins; and (c) antibody levels against the nonstructural protein NS1 present in the A/H1N1 2009 virus, but not in the Pandemrix vaccine (31).
Methods
Study populations
Serum samples were obtained from 43 Swedish patients diagnosed with narcolepsy following multiple sleep latency test during the period between January 1, 2009, and December 31, 2010. The Pandemrix vaccine had been given to all 43 individuals (Table 1) and blood samples were obtained 1.5–2 years after vaccination, as previously described (27). Also included were 64 Pandemrix-vaccinated healthy controls randomly selected from the Swedish population register and asked to participate by mail. Healthy controls were matched to patients based on age and gender in a 1:2 ratio (Table 1). The identification of patients and controls, including the presence or absence of HLA DQ B1*06:02, is given elsewhere (25,27).
Ages correspond to October 2009, which was the start of the Swedish Pandemrix® vaccination campaign.
HLA, human leukocyte antigen.
Cloning A/H1N1 nucleoprotein and nonstructural protein complementary DNA
The influenza A virus [A/California/04/2009(H1N1)] segment 5′ nucleocapsid protein (NP) gene (GenBank accession FJ966083.1), influenza A virus [A/Puerto Rico/8/1934(H1N1)] NP gene (GenBank accession CY009447.1), and influenza A virus [A/California/04/2009(H1N1)] nonstructural protein 1 (GenBank accession ACP41110.1) were synthesized and cloned into the pJ204 vector by DNA2.0 (San Diego, CA).
Recombinant genes were designed with flanking restriction enzyme sites Xhol and Xbal (NP CA2009, NP PR1934) or KpnI and Xbal (H1N1 NS1) (FastDigest™; Fermentas, Helsingborg, Sweden). Kozak encoding sequence GCCACC was inserted between the start codon and the 5′ restriction site to increase initiation of translation. The protein encoding sequences were cut from the original vector, subcloned into the pTNT vector (Promega, Southampton, United Kingdom), and transformed into Escherichia coli DH5 (α) (NP PR1934 and H1N1 NS1) or Invitrogen one-shot inv110 (NP CA2009) competent cells. Complementary DNA (cDNA) plasmid extraction using the QIAprep Spin Miniprep kit (Qiagen, Solna, Sweden) was followed by sequence verification by GATC Biotech AG (Konstanz, Germany).
Generation of 35S-A/H1N1 proteins by in vitro transcription translation
Radiolabeled A/H1N1 proteins were synthesized using the TNT SP6-coupled reticulocyte lysate system (Promega) for in vitro transcription translation. cDNA pTNT-A/H1N1 protein plasmids (1 μg/μL, final concentration 0.02 μg/μL) were mixed with 35S-methionine (PerkinElmer Life and Analytical Sciences, Brussels, Belgium) and the TNT SP6-coupled reticulocyte lysate system. Incubation was carried out for 90 min at 30°C, followed by removal of the unincorporated label using Nap-5 columns (GE Healthcare Bio-Sciences, Uppsala, Sweden). The mean incorporation rate of the radioactive label was 39%, 49%, and 55% for NS1 CA2009, NP CA2009, and NP PR1934, respectively. Both nucleoproteins contained 498 amino acids with 91.6% shared homology (456/498 amino acids, Supplementary Fig. S1A; Supplementary Data are available online at
Influenza 35S A/H1N1 nucleoprotein radiobinding antibody assay
Radiolabeled A/H1N1 nucleoproteins obtained through in vitro transcription translation were used as antigens to detect antibodies in 2.5 μL serum in duplicate (12). A total of 60 μL of radiolabeled protein, corresponding to 24,000 counts per minute (cpm) in total (400 cpm/μL), was incubated overnight diluted in Tris-buffered saline (pH 7.4) supplemented with 0.15% (v/v) Tween 20, 0.1% (w/v) bovine serum albumin, and 1% (v/v) Triton X-100 (TBST). Antibody-bound proteins were separated from free 35S-labeled A/H1N1A nucleoproteins using protein A-Sepharose (Zymed Laboratories, Inc., San Francisco, CA) after transfer to filtration plates (Millipore, Solna, Sweden). The plates were washed using TBST and Super Mix scintillation cocktail (PerkinElmer) was added and the antibody-bound 35S-labeled A/H1N1 nucleoproteins were counted in a Wallac Microbeta Trilux beta counter (PerkinElmer).
A serum sample with high A/H1N1 nucleoprotein antibody level was used as an in-house standard, diluted in seven steps starting with 1:12,800, and used to express A/H1N1 nucleoprotein antibody levels in arbitrary U/mL. A total of 250 U/mL of the in-house standard serum corresponded to 2.5 μg/mL in the 35S-NP PR1934 assay and 4.6 μg/mL in the 35S-NP CA2009 assay of commercial rabbit polyclonal antibody against recombinant A/H1N1 Puerto Rico/8/34/Mount Sinai nucleoprotein (Sino Biological, Inc., Beijing, China; Supplementary Fig. S1B).
Initial analyses of all serum samples resulted in values higher than the maximum of the standard curve. All samples were therefore diluted to 1/100. The standard curve was linear up to about 200 U/mL; therefore, all samples with higher A/H1N1 nucleoprotein antibody levels than 180 U/mL at dilution 1/100 were titrated in additional steps; 1/200, 1/400, and 1/800 to fit the standard curve. Duplicate samples with a coefficient of variation (CV) ≥12% were reassayed. The intra-assay CV was 2.9% and 3.3% for NP CA2009 and NP PR1934 antibody analyses, respectively. Two control samples were included in all assays and the interassay CV was 11% and 9% for NP CA2009 and NP PR1934, respectively.
Competitive radiobinding assay using recombinant influenza A/H1N1 nucleoproteins
Using a reciprocal permutation strategy as previously described (44), serum samples from selected patients and controls (<13 years) were incubated with previously described in vitro produced 35S-radiolabeled nucleoproteins (35S-NP CA2009 or 35S-NP PR1934) in combination with either nonlabeled recombinant influenza A virus H1N1 (A/Puerto Rico/8/34/Mount Sinai) nucleoprotein (Sino Biological, Inc.) or influenza A virus H1N1 (A/California/07/09) nucleoprotein (Sino Biological, Inc.).
Antibody analyses were performed as previously described (44); 30 μL of 35S-NP (NP PR1934 or NP CA2009) diluted to 850 cpm/μL was mixed with 30 μL of commercially obtained nonlabeled nucleoproteins (final concentrations: 0, 0.0005, 0.005, 0.005, 0.5, and 5 μg/mL). The intra-assay coefficients of variation were determined as 6.4% and 5.1% for 35S-NP CA2009 displacement and as 7.3% and 3.8% for 35S-NP PR1934 displacement, respectively. 35S-NP specificity was tested in three high-titer serum samples through displacement with nonlabeled recombinant HA2009 (Sino Biological, Inc.; Supplementary Fig. S1C, D).
Influenza 35S A/H1N1 nonstructural protein (H1N1 NS1) radiobinding antibody assay
Antibodies against A/H1N1 NS1 were determined in a radiobinding assay as described for the A/H1N1 NPs. The cutoff for H1N1 NS1 antibodies was 1,525 U/mL at the 97.25th percentile in serum from 200 healthy blood donors ascertained in 2008 (21) (Supplementary Fig. S2). A serum sample with high A/H1N1 NS1 antibody titer was diluted 1:1,600–1:25 (1,000 U/mL) as an in-house standard to express A/H1N1 NS1 antibody levels in arbitrary U/mL. The intra-assay CV was 5%. The interassay CV for three in-house controls included in each assay was 14%.
HLA DQ typing
HLA DQB1*06:02 was identified with the linked SNP rs9271366 (GG-homozygous DQB1*0602, AG-heterozygous, AA-non-DQB1*06:02) using TaqMan allelic discrimination assays, as outlined elsewhere (17,28).
Ethical considerations
The study was approved by the Regional Ethics Review Board in Stockholm. All study participants or parental guardian(s) gave their written consent.
Statistical analyses
Median levels between groups were compared using Mann–Whitney's U test and Wilcoxon signed-rank test (paired comparisons). Associations between two continuous variables were measured using Spearman's ρ (rho). A four-parameter log-logistic function was fitted to the displacement data for each individual and displacement type using the drc package in R (42). The fitted function is y = D + (A − D)/(1 − (x/C)^D), where y is the binding (%) and x the concentration. The parameters A, B, C, and D were constrained to positive values during optimization. Maximal displacement percentage was computed as 100 − D, where D is the parameter that represents the minimal binding value. The affinity was calculated as half-maximal binding, Kd50, and expressed in ng/mL.
The significance threshold is set to 0.05 for the initial tests on antibody levels and antibody levels in association with age. For the following tests on displacement statistics (affinity and maximal displacement) as well as the tests of association between antibody levels and antibody levels and affinity, we use a lower significance threshold of 0.005, motivated by the multiple tests performed.
Results
A/H1N1 nucleoprotein antibodies
Binding of A/H1N1 antibodies did not differ between narcolepsy patients and controls. Median antibody binding levels toward influenza A virus [A/California/04/2009(H1N1)] nucleoprotein (NP-CA2009) in serum samples from the 43 narcolepsy patients were 12,697 U/mL compared with 13,215 U/mL in 64 matched healthy controls (p = 0.6, Fig. 1). The median antibody binding levels for influenza A virus [A/Puerto Rico/8/1934(H1N1)] nucleoprotein (NP-PR1934) were 15,709 U/mL for the patients compared with 16,753 U/mL for controls (p = 0.896, Fig. 1).
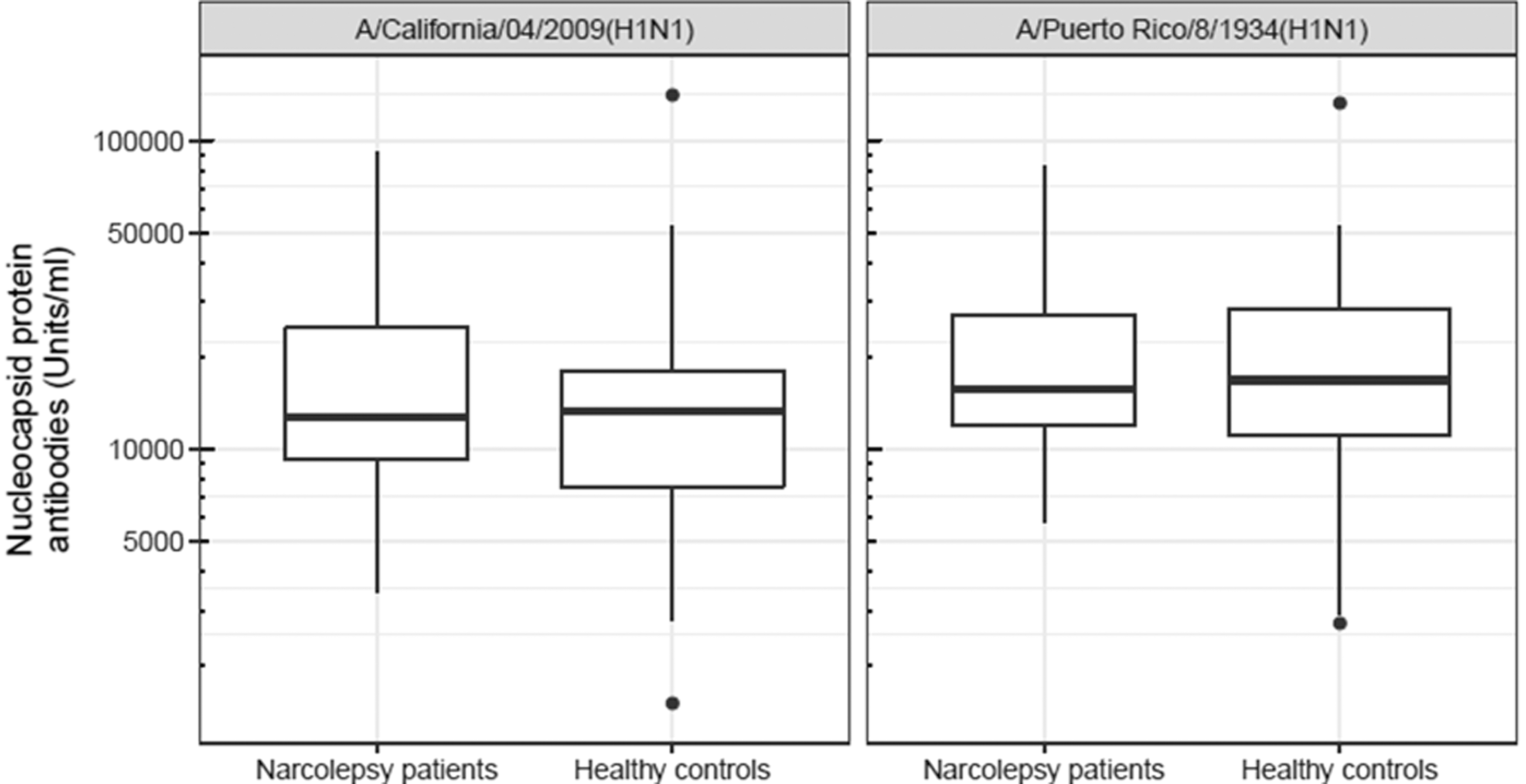
Nucleoprotein antibody levels. Median levels (
A/H1N1 nucleoprotein antibodies in relation to age
The levels of NP antibodies in the patients tended to decrease with increasing age (NP-CA2009: ρ = −0.280, p = 0.069; NP-PR1934: ρ = −0.271, p = 0.079) in contrast to the controls where the levels increased with increasing age for NP-PR1934 (ρ = 0.280, p = 0.025), but not for NP-CA2009 (ρ = 0.206, p = 0.102, Fig. 2A). A permutation test, adopted to investigate if such a large difference in ρ between the groups was to be expected by chance, showed that the opposite relationship was unlikely to occur by chance for both NP-CA2009 (p = 0.0153) and NP-PR1934 (p = 0.0058).
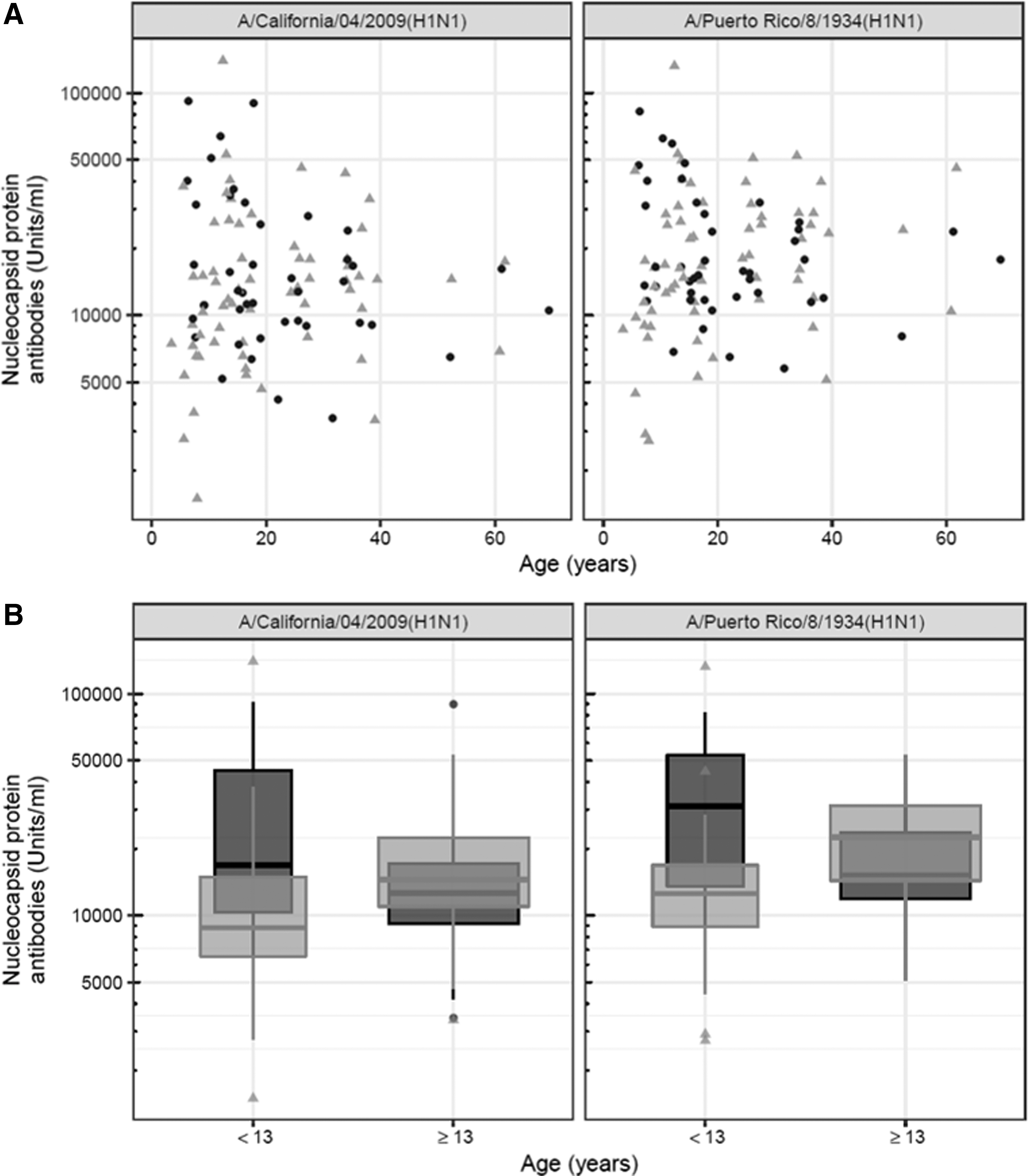
To further study the differences between younger and older individuals, both patients and controls were divided into groups based on age in October 2009 when the Swedish Pandemrix vaccination campaign was initiated: childhood patients (n = 11), childhood controls (n = 21), older patients (≥13 years: n = 32), and older controls (≥13 years: n = 43), respectively. The decision to compare subjects older and younger than 13 years was motivated by our previous study (27) in combination with the suggestion that Pandemrix vaccination might have induced younger age onset compared with idiopathic narcolepsy (45).
Median antibody levels were higher in the childhood patients compared with the childhood controls for both NP-CA2009 (p = 0.0339) and NP-PR1934 (p = 0.0246, Table 2). Median antibody levels among the older patients compared with their controls were comparable for NP-CA2009 (p = 0.279) and NP-PR1934 (p = 0.0644, Table 2 and Fig. 2B).
Median and range of antibody levels for patients and controls within the two age groups, <13 and ≥13 years of age.
The three p-value columns present (unadjusted) p-values from the following tests: aMann–Whitney's U test comparing median values between patients and controls within the age groups, bMann–Whitney's U test comparing the age groups for patients, and ccontrols.
The childhood patients did not differ from the older patients, neither in antibody levels against NP-CA2009 (p = 0.219) nor against NP-PR1934 (p = 0.0981, Table 2). In contrast, the childhood controls compared with older controls had lower antibody levels against both NP-CA2009 (p = 0.0221) and NP-PR1934 (p = 0.00619, Table 2 and Fig. 2B).
A/H1N1 nucleoprotein displacement
We next determined antibody affinity toward both NP-CA2009 and NP-PR1934 in reciprocal permutation competitive assays with serum samples from childhood narcolepsy patients (<13 years of age; age range 7–12 years, n = 11) and age-matched controls (<13 years of age, age range 3–12 years, n = 21, Fig. 3A). All samples were diluted to 50% of maximal binding, equivalent to ∼125 U/mL (Supplementary Table S1). The dilutions were, however, not exactly to 125 U/mL, although they did not differ between patients and controls for NP-CA2009 (122 U/mL range 69–188 U/mL to 98 U/mL range 14–182 U/mL, p = 0.133) or NP-PR1934 (130 U/mL range 77–186 U/mL to 114 U/mL range 21–181 U/mL, p = 0.123, Supplementary Table S1).
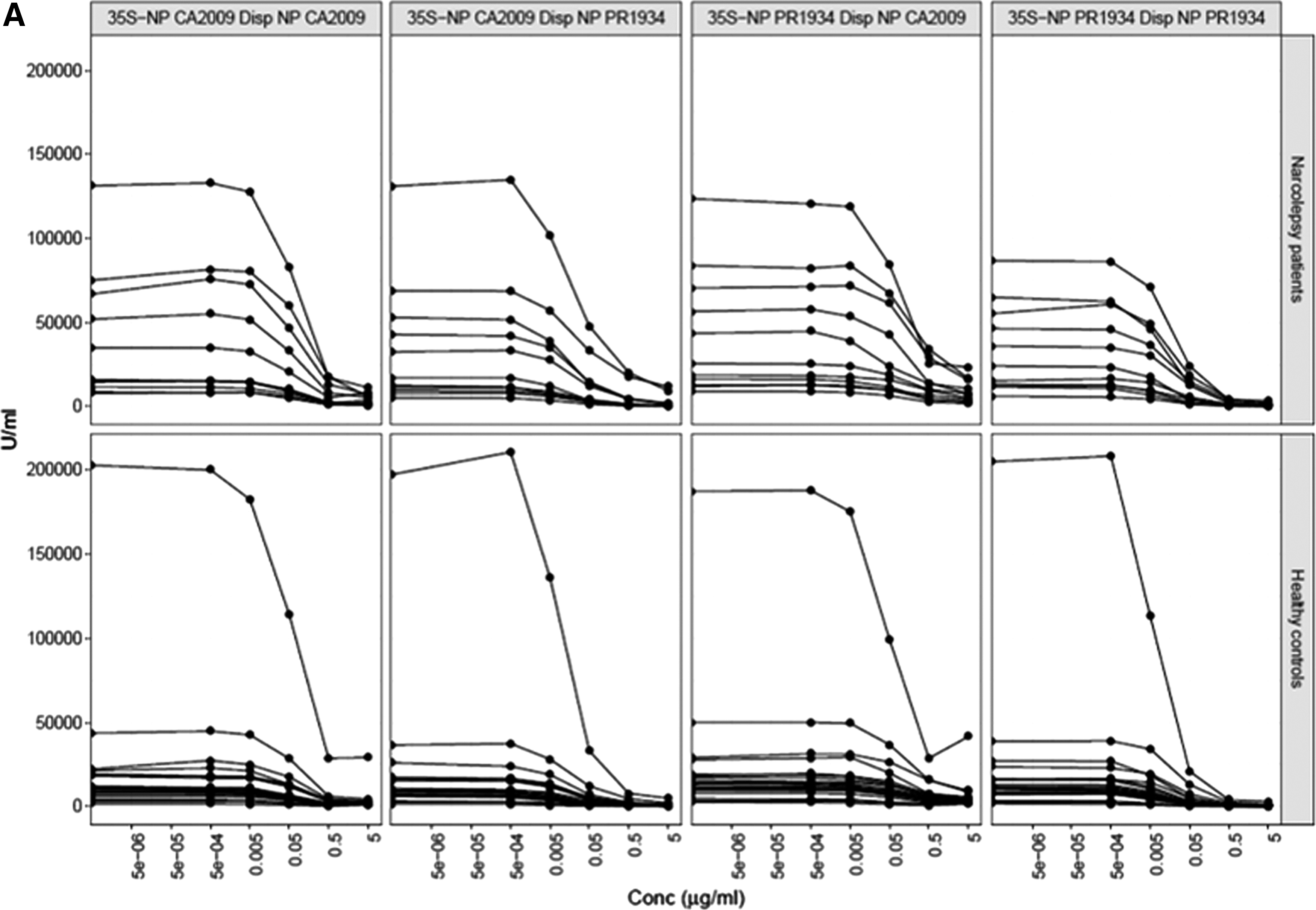
The levels (U/mL) for NP-CA2009 and NP-PR1934 correlate strongly (ρ = 0.904, p < 4.0e-08). The NP-PR1934 values were generally higher than the NP-CA2009 values (p = 1.47e-04). The coefficients of variation for the three endpoint measurements were 11.58% and 11.63% for NP-CA2009 and NP-PR1934, respectively. The variable endpoint levels within patients and controls, as well as between the two groups, are illustrated in Figure 3A after correcting the levels at 50% binding with the different degrees of dilution between the samples (Fig. 3A).
Maximal displacement
After dilution of all samples to about 50% binding, the maximal displacement (%) for NP-CA2009 did not differ when using nonlabeled NP-CA2009 (90.3%) or nonlabeled NP-PR1934 (95.0%), neither in the patient serum sample (p = 0.15) nor in the controls (nonlabeled NP-CA2009: 90.7%; nonlabeled NP-PR1934: 95.6%, p = 0.01, Table 3 and Fig. 3B). Maximal displacement (%) of 35S- NP-PR1934 in the patients was greater when using nonlabeled NP-PR1934 (98.4%) compared with nonlabeled NP-CA2009 (80.3%) (p = 0.00098) similar to what was observed in the controls as nonlabeled NP-PR1934 displaced 97.5% compared with 79.5% with nonlabeled NP-CA2009 (p = 2.9e-06, Table 3 and Fig. 3B).
Median and range of Kd50 and maximal displacement percentage (computed from the fitted four-parameter log-logistic curves) values for patient and control samples.
The three p-value columns result from tests: aWilcoxon signed-rank test comparing the displacements with NP CA2009 and NP PR1934, bMann–Whitney U test comparing median values for patients and controls in the displacements with NP CA2009, and cNP PR1934.
NP, nucleocapsid protein.
Maximal displacement (%) did not differ between patients and controls for 35S-NP-CA2009 displacement with either nonlabeled NP-CA2009 (p = 0.81) or nonlabeled NP-PR1934 (p = 0.33), or 35S-NP-PR1934 displacement with nonlabeled NP-CA2009 (p = 0.64) or nonlabeled NP-PR1934 (p = 0.97, Table 3 and Fig. 3B), respectively.
Affinity
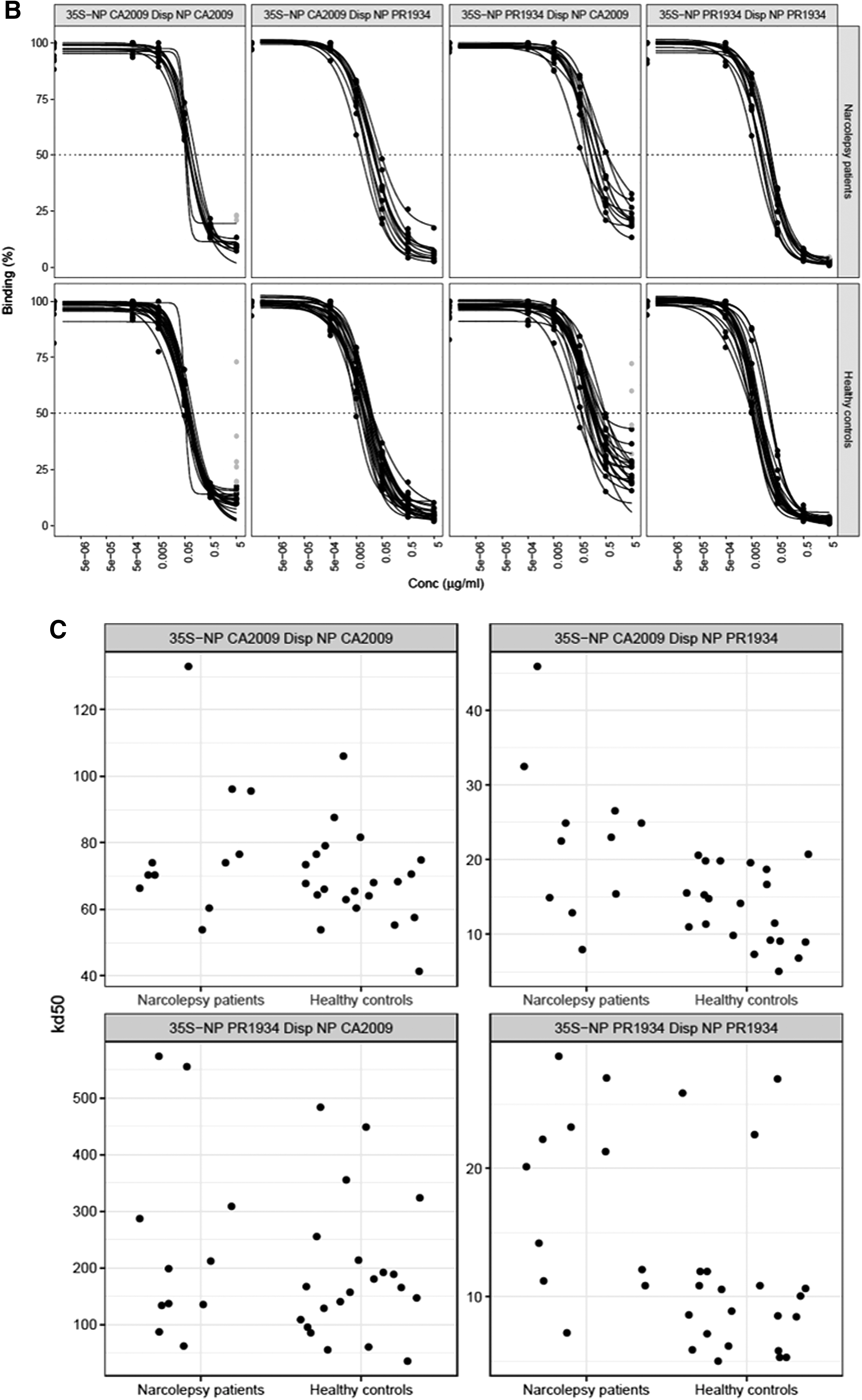
Affinity, expressed in ng/mL as the inversed measurement of half-maximum binding (Kd50), was compared between patients and controls in reciprocal permutation (Fig. 3C).
First, the affinity of NP-CA2009 antibodies against 35S-NP-CA2009 was similar in the patients (74 ng/mL) compared with the controls (68 ng/mL; p = 0.21).
Second, the affinity of NP-PR1934 antibodies against35S-NP CA2009 was lower (23 ng/mL) in the patients compared with the controls (14 ng/mL; p = 0.0048).
Third, the affinity of NP-CA2009 antibodies against 35S-NP-PR1934 did not differ between patients (199 ng/mL) and controls (165 ng/mL; p = 0.48).
However, fourth, the affinity of NP-PR1934 antibodies against 35S-NP-PR1934 was reduced in the patients (20 ng/mL) compared with the controls (9 ng/mL, p = 0.0031, Table 3 and Fig. 3C).
The affinity was different for NP-CA2009 antibodies compared with NP-PR1934 antibodies against 35S-NP-CA2009 for the patients (p = 0.00098) as well as for the controls (p = 9.5e-07). Affinity was different for NP-CA2009 antibodies compared with NP-PR1934 antibodies against 35S-NP-PR1934 for the patients (p = 0.00098) and the controls (p = 9.5e-07).
Nucleoprotein antibodies in relation to H1N1 HA antibodies and HLA DQB1*06:02 association
NP-CA2009 and NP-PR1934 antibodies correlated strongly for patients (ρ = 0.936, p < 2.2e-16) and controls (ρ = 0.962, p < 2.2e-16, Supplementary Table S2). There was no association between the levels of the two different NP antibodies and previously reported H1N1 HA antibody levels (27) (Supplementary Table S2). No association was found between the levels of either NP antibody and HLA DQB1*06:02 (Supplementary Table S3).
A/H1N1 nonstructural protein antibodies
Median antibody binding level toward influenza A virus [A/California/04/2009(H1N1)] nonstructural protein (NSI) was 23 (8–308) U/mL in the 43 narcolepsy patients compared with 23 (3–3,097) U/mL for 64 healthy controls (p = 0.399, Fig. 4A). Using the 97.25th percentile of 200 blood donors ascertained in 2008 as controls for a cutoff, it was found that 0% (0/43) in the patient group and 4.7% (3/64) in the healthy control group were considered positive for H1N1 NS1 antibodies (p = 0.272).
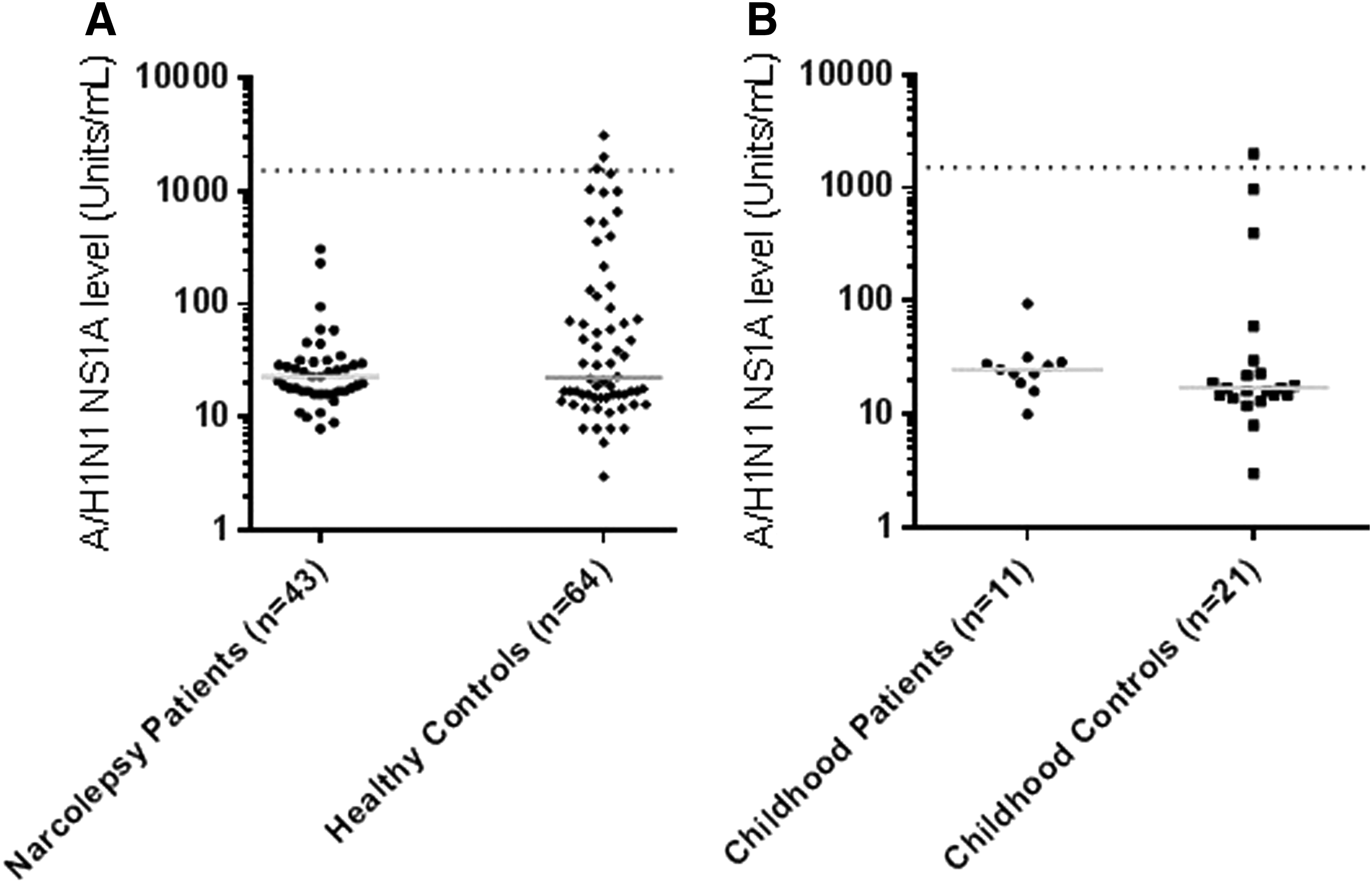
In the 11-childhood patient subgroup, the median NS1 antibody binding level was 25 (10–95) U/mL compared with 17 (3–1,999) U/mL in the subgroup of 21 healthy childhood controls (p = 0.123, Fig. 4B). Using the 97.5th percentile of the blood donors, 0% (0/12) in the patient group compared with 4.8% (1/21) among the healthy childhood controls was considered positive for H1N1 NS1 antibodies.
Discussion
In the present study, we determined antibody levels toward both wild-type A/H1N1 2009[A/California/04/2009(H1N1)] (NP-CA2009) and Pandemrix [A/Puerto Rico/8/1934(H1N1)] (NP-PR1934) nucleoproteins in narcolepsy patients and age-matched controls. The major finding was that childhood patients had a lower antibody affinity against NP-PR1934 compared with healthy, childhood Pandemrix-vaccinated controls. It was also noted that antibody affinities in childhood patients and controls were higher against NP-PR1934 than NP-CA2009. Our observation that none of the narcolepsy patients were positive for A/H1N1 NS1 antibodies indicates that they may not have been exposed to the H1N1 virus.
The affinity analysis was restricted to childhood patients not only primarily because they had higher levels of antibodies against HA2009 (27), NP-CA2009, and NP-PR1934 but also because childhood age at onset was unique to the Pandemrix-associated onset of narcolepsy (4,37,39,41,45). It is noteworthy that the association between antibody levels and age was only positive for NP-PR1934 among the controls. Previous studies of antibody levels in relation to the 2009 influenza pandemic found an age-dependent variation, with higher levels of cross-reactive antibodies among the elderly (16,19,30).
The higher antibody affinity toward the Pandemrix-associated NP-PR1934 compared with influenza pandemic NP-CA2009 is remarkable given the high similarity in the amino acid sequence between the two nucleoproteins (Supplementary Fig. S1A). A cross-reactivity of antibodies (29) has most probably occurred, as indicated by the strong correlation in antibody levels between the nucleoproteins as well as with the commercially obtained polyclonal antibody toward NP PR1934 that also reacted with NP CA2009 (Supplementary Fig. S1B).
We report a lower NP-PR1934-specific affinity for young patients despite the higher NP antibody levels compared with healthy controls. Our results suggest that the vaccine-associated nucleoprotein had a stronger ability to evoke an immune response. This reaction might have been unexpected as the primary reason for switching NP-CA2009 for NP-PR1934 was to increase the yield of the vaccine (43). It can therefore not be excluded that the resulting hybrid virus contributed to an unwanted immune response that may have triggered narcolepsy.
The question regarding whether the 2009 influenza H1N1 pandemic contributed to narcolepsy is contentious; reports of increased incidence of nonvaccine-associated narcolepsy have emerged from China (14,15). This is opposite to findings from European studies, which report no indication of wild-type A/H1N1 2009 infection as a prerequisite for narcolepsy (31). It is therefore possible that childhood narcolepsy following vaccination was related to the absence of previous influenza exposure (16).
We therefore speculate that the patients had no history of previous H1N1 exposure. The absence of prior cross-reactive H1N1 antibodies would explain the vigorous immune response against NP-PR1934 compared with NP-CA2009. However, the vigorous immune response resulted primarily in low-affinity NP antibodies, which might have been associated with the ability to generate broadly neutralizing influenza antibodies. Low-level neutralizing antibodies might allow prolonged exposure to the wild-type A/H1N1 2009[A/California/04/2009(H1N1] virus and consequent damage to the hypocretin neurons.
Narcolepsy has been associated with Pandemrix (2) used in Europe (34,37), but not with Arepanrix® (1,51) used in Canada and South America (35). Both vaccines were developed through a reassortment approach (23,24,43); coinfection in eggs of wild-type and donor strains was followed by selection of a high-growth reassortant strain with antigenic characteristics of the wild-type virus. The new A/reassortant/NYMC X-179A (NYMC X-157 × A/California/07/2009) influenza strain was selected with HA, NA, and PB1 genes from A/California/07/2009 (H1N1v) and the viral M, NP, NS, and PB2 genes from A/PR/8/34 (52).
Pandemrix and Arepanrix included both the AS03 adjuvants, but the use of detergents such as Polysorbate 80 and Triton X-100 in Pandemrix may have modified in vitro antigenic epitopes (50). Different purification and isolation protocols (Fluarix process—Pandemrix; Flulaval process—Arepanrix) were used at different manufacturing locations (Pandemrix; GSK, Dresden, Germany, and Arepanrix; GSK, Quebec, Canada) and whether these have resulted in antigenic differences are contentious (7,20,43,50).
Differences in the association of narcolepsy with Pandemrix (Europe) and Arepanrix (Canada) pandemic influenza vaccines are speculated to involve viral antigenicity (20,50). Efforts to measure the amount of NP have identified differences in the amount of NP antigen in each of these vaccines, which along with the lower prevalence of the narcolepsy risk allele HLA DQB1:0602 in Canada, in part, may explain differences in postvaccination appearance of narcolepsy with these two vaccines. In addition, the influenza nucleoproteins in the wild-type A/H1N1 2009 pandemic and Pandemrix vaccine were different (50).
Studies investigating the 1918 influenza pandemic suggest that the fatality could be due to the own immune system perhaps causing a cytokine storm (36), therefore affecting younger healthy individuals (47). The etiology of idiopathic narcolepsy is not known. Whether it can be linked to virus-induced disease in the brain as in experimental mice (48) or to alternative trigger mechanisms remains to be determined. Different trigger mechanisms may explain the epidemiological data from China suggesting an increased incidence of narcolepsy during the 2009 pandemic (14,15).
In the present study, we did not observe that NP antibody levels were related to HLA DQB1*06:02 (Supplementary Table S2). Although this would seem to be in contrast to recent studies suggesting that nucleoprotein antibody levels (3,50) were related to DQB1*06:02, it should be noted that our study of influenza A virus [A/California/04/2009/(H1N1)] HA antibody levels showed an association with DQB1*06:02 (27). A potential weakness of the present study was that samples were not available from all subjects in the original case–control study (25). It would be of interest to further analyze virus protein affinities in an extended group of narcolepsy patients and controls. It is noted that NP-PR1934 is not specific for the Pandemrix vaccine, but that it is commonly used in H1N1 vaccine production (23,43).
It is currently not explained how NP CA2009 and NP PR1934 antibody levels and affinity correlate with development of narcolepsy following Pandemrix vaccination. Further research is needed also to include analyses of antibody levels and affinities in adults. It is important to note that Pandemrix-induced narcolepsy primarily affects children, therefore this group were selected for this study.
In conclusion, an altered immune response among childhood narcolepsy patients toward influenza nucleoproteins is reported in the present study. Our data suggest increased antibody levels, but lower NP antibody affinity in the patients compared with controls. The final immunological mechanisms connecting Pandemrix to narcolepsy remain elusive, but the present study warrants further studies of antibody affinities that should be focused on the H1N1 viral protein components of the vaccine.
Footnotes
Acknowledgments
The authors thank the Lund University Medical Faculty for support and acknowledge the Swedish Medical Products Agency for the collection of blood samples. The study was supported by Filip Lundbergs stiftelse, Fredrik och Ingrid Thurings Stiftelse, Gunvor och Josef Anérs stiftelse, Kronprinsessan Lovisas Förening För Barnasjukvård/Stiftelsen Axel Tielmans Minnesfond, Linnéa och Josef Carlssons Stiftelse, Neuro Sweden, Rune Ljungdahls Stiftelse, the Segerfalk Foundation, Stiftelsen Samariten, Stiftelsen till minne av Personalföreningarna i Holmia Försäkring AB, and Stohnes Stiftelse.
Author Disclosure Statement
No competing financial interests exist
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.