
Abstract
Fowl aviadenoviruses (FAdVs) are distributed worldwide in poultry farms. Some FAdVs are the causative agents of inclusion body hepatitis and hydropericardium syndrome that cause significant economic losses to the poultry industry. In contrast with human adenovirus, the study of the molecular biology of FAdV is still far behind. We previously showed that FAdV-9 open reading frame 1 (ORF1) is a dUTPase enzyme that contributes to the upregulation of type I interferons and is not required for virus replication in vitro. In the present study, we compared virus replication in vivo and the host immune response in chickens orally inoculated with a dUTPase knockout virus (ORF1stop), the rescued version of ORF1stop (resORF1), and wtFAdV-9. Our data showed that replication of ORF1stop was delayed on days 1 and 3 postinoculation compared with wtFAdV-9, as evidenced by significantly less virus shedding in feces and lower viral loads in tissues. Moreover, we found that there was a significant difference in the induction of cytokine gene mRNA expression in tissues and IgG antibody responses in ORF1stop versus wtFAdV-9-infected chickens, suggesting that ORF1 plays some roles in modulating the host immune response. Our study provides useful data on the mechanism of the host immune response against FAdV infection.
Introduction
F
Compared with human adenoviruses, the molecular biology of FAdVs is less well understood. We recently identified open reading frame 1 (ORF1) of FAdV-9 as a functional dUTPase enzyme (8). dUTPase is a ubiquitous enzyme that catalyzes the cleavage of dUTP to dUMP and pyrophosphate, thereby leading to low dUTP/dTTP ratios in cells and preventing the misincorporation of deoxyuridine into DNA. To date, the dUTPase gene has been characterized in many viruses, such as adenovirus (36), herpesvirus (12), retrovirus (27), African swine fever virus (26), and poxvirus (6).
A dUTPase knockout FAdV-9 virus (ORF1stop) and its rescued version (resORF1) were generated in our laboratory for functional studies. Our data showed that FAdV-9 dUTPase was not essential for virus replication in vitro as the knockout of ORF1 did not affect viral DNA replication and the production of progeny virus in chicken hepatoma cells (CH-SAH) and primary chicken embryo liver (Celi) cells. For herpes simplex virus (HSV), the deletion of dUTPase did not affect virus replication in vitro, but caused reduced viral replication in the central nervous system of mice (29). Leang et al. demonstrated that dUTPase of Murid herpesvirus 68 (MHV-68) is necessary for efficient virus replication in the lungs of infected mice (18).
Therefore, the aim of this study was to explore the effects of FAdV-9 dUTPase in virus replication and in modulating the host immune response against viral infection in orally inoculated chickens. Virus shedding, viral load in liver and cecal tonsil, cytokine gene expression in spleen, liver, and cecal tonsil, and antibody levels were studied.
Materials and Methods
Viruses and cells
ORF1stop, the dUTPase knockout virus, and resORF1, the ORF1-rescued virus, were both generated through site-directed mutagenesis and homologous recombination, based on a FAdmid clone pPacFAdV-9 that contains the whole FAdV-9 genome (9,24). After rescue, both viruses were passaged three times in CH-SAH followed by sequencing their genomes. The confirmed mutant viruses, together with the wild-type virus, wtFAdV-9, were propagated and titrated in CH-SAH cells as described previously (1). CH-SAH cells were maintained in Dulbecco's modified Eagle's medium/nutrient mixture F-12 Ham (DMEM-F12), supplemented with 10% nonheat-inactivated fetal bovine serum, 2 mM L-glutamine, 100 U/mL penicillin, and 100 μg/mL streptomycin.
Experimental animals
One-day-old specific pathogen-free white Leghorn chickens were obtained from the Canadian Food Inspection Agency (Ottawa, ON, Canada) and housed in the Isolation Unit of the University of Guelph throughout the study. All animal experiments were reviewed and approved by the Animal Care Committee of the University of Guelph according to the Guide to the Care and Use of Experimental Animals of the Canadian Council on Animal Care.
There were 164 chickens in the experiment. At 10 days of age, four birds were euthanized and tissues were collected to serve as negative controls. The remaining 160 chickens were divided into four groups (each with n = 40) and orally inoculated with (1) phosphate-buffered saline (PBS), which served as a mock control; (2) 2 × 107 plaque-forming units (pfu)/chick ORF1stop virus; (3) 2 × 107 pfu/chick resORF1 virus; and (4) 2 × 107 pfu/chick wtFAdV-9, respectively.
At 0, 1, 3, 5, 7, 10, 14, 21, 28, and 35 days postinoculation (d.p.i.), cloacal swabs were collected from all birds (n = 40, 35, 30, 25, 20, 15, 15, 10, 10, and 10, for each time point, respectively) in 1 mL PBS supplemented with 100 U/mL penicillin and 100 μg/mL streptomycin to determine the virus titer in feces. At 10 h.p.i., 1, 3, 5, 7, 14, and 35 d.p.i., liver and cecal tonsil were collected from five birds in each group to determine the viral load in tissues. Liver, spleen, and cecal tonsil were collected from the same five birds in each group to investigate cytokine gene expression, including interferon (IFN)-α, IFN-β, IFN-γ, interleukin (IL)-10, and IL-8. In addition, serum samples were collected from all birds at 0 (n = 40), 7 (n = 20), 14 (n = 15), 21 (n = 10), 28 (n = 10), and 35 (n = 10) d.p.i. to test for FAdV-specific IgG antibodies.
DNA and RNA extraction
Viral DNA was extracted from tissues stored at −80°C with the QIAamp DNA Mini Kit (Qiagen, Inc.) following the manufacturer's instructions. The DNA concentration was measured using a NanoDrop 2000 spectrophotometer. RNA was extracted with TRIzol (Invitrogen) from tissues stored in RNAlater as described previously (9), before digestion with DNase I (Fermentas) and quantification with a NanoDrop 2000 spectrophotometer.
Quantitative polymerase chain reaction and quantitative real-time polymerase chain reaction
To determine the viral load in tissues, viral DNA was quantified by quantitative polymerase chain reaction (qPCR) with the LightCycler® 480 SYBR Green I Master Kit (Roche Diagnostics) and FAdV-9-specific primers as previously described (32). To determine the cytokine gene expression in tissues, cDNA was first synthesized from total RNA with SuperScript™ II Reverse Transcriptase (Invitrogen) and random primers. The cDNA was diluted 1:10 and used for quantitative real-time polymerase chain reaction with the LightCycler 480 SYBR Green I Master Kit (Roche Diagnostics) and gene-specific primers as previously described (8,9,13).
Enzyme-linked immunosorbent assay
Heat-inactivated serum samples were diluted 1:25 in wash buffer (0.05% Tween 20 in PBS). The IgG levels were detected by enzyme-linked immunosorbent assay (ELISA) and presented as sample to positive (S/P) ratios (25).
Statistical analyses
Statistical analyses were performed using GraphPad Prism 6.0 software (San Diego, CA). A one-way analysis of variance was used to determine significant differences between any two groups. The critical level for significance was set at a p value of <0.05. Data were expressed as mean ± standard error of the mean.
Results
Virus shedding
To compare virus shedding in chickens among ORF1stop, resORF1, and wtFAdV-9 groups, cloacal swabs were collected for the determination of virus titer in feces by plaque assay. All samples from chickens before inoculation and the mock-infected group were negative (data not shown), and the virus titers for other groups are summarized in Table 1. Virus was detected in chickens of all three virus-infected groups from day 1 to 14 p.i., but no virus was detected at days 21, 28, and 35 p.i. for any group. The peak of virus shedding in all three groups occurred at day 5 p.i., with an approximate titer of 5.0 × 104 pfu/mL. However, at days 1 and 3 p.i., the virus titer in the samples of ORF1stop group was significantly lower (∼4 and 14-fold at 1 and 3 d.p.i., respectively) than that of the wtFAdV-9 group, whereas no significant difference was found at later days. The virus titers of samples from the resORF1 group were similar to those of wtFAdV-9 throughout the experiment.
Values of p < 0.05 for titers are statistically significant with respect to wtFAdV-9.
Percentage of chickens shedding virus.
N/A, not applicable; ND, not detected; pfu, plaque-forming units; p.i., postinoculation.
Viral loads in tissues
To determine the viral loads in tissues, the viral genome copy numbers in livers and cecal tonsils were quantified by real-time PCR with specific FAdV-9 primers. No virus was detected in samples from chickens before inoculation and mock-infected groups (data not shown). The results of the other groups are summarized in Table 2. In livers and cecal tonsils, the viral genome copy numbers were 4.3 and 3.3-fold lower (p < 0.05) in the ORF1stop group than that in wtFAdV-9 groups at day 1 p.i., and were 3.8 and 2.5-fold lower (p < 0.05) in the ORF1stop group than that in the wtFAdV-9 groups at day 3 p.i., respectively. There was no significant difference between the ORF1stop group and the wtFAdV-9 group at other time points. The resORF1 group showed very similar results to wtFAdV-9, and there was no significant difference between these two groups at any time point.
CT, cecal tonsil; L, liver.
Antibody response
Antibody levels against FAdV-9 were determined to investigate the role of ORF1 on the humoral immune response. Anti-FAdV antibodies were not detected in samples from any group before inoculation. The weekly ELISA results from days 7 to 35, based on the S/P ratio, are shown in Figure 1. Throughout the experiment, no anti FAdV antibodies were detected in the mock group, whereas antibody levels increased in all virus-infected groups from 1 week p.i. until the end of the experiment. The antibody level (S/P ratio) in the ORF1stop group was significantly lower than that of wtFAdV-9, except at 1 week p.i., whereas the resORF1 group had a similar antibody response to that in wtFAdV-9 over the course of the experiment.
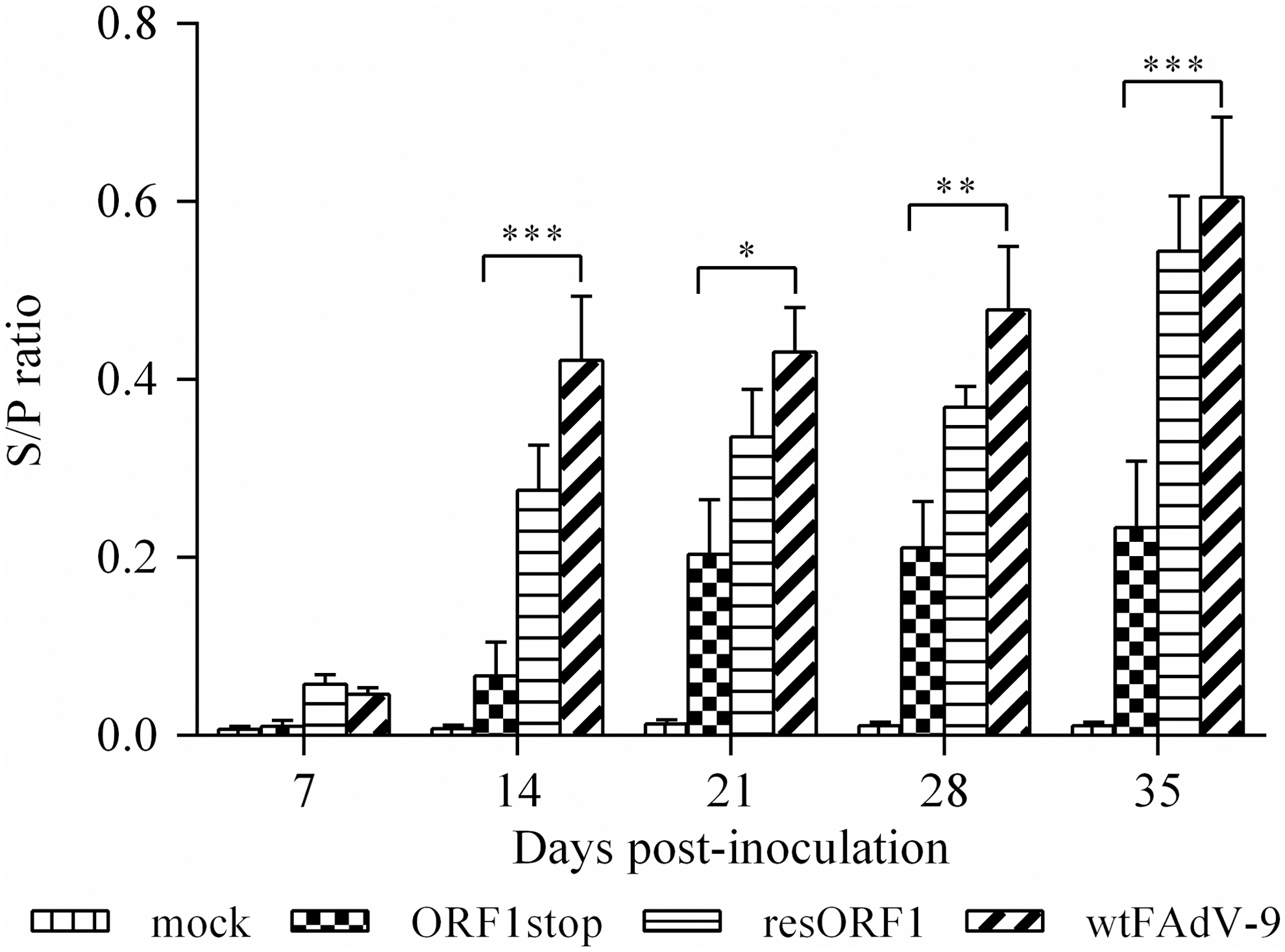
FAdV-specific IgG antibody response in chickens. Chickens were orally inoculated with ORF1stop, or resORF1, or wtFAdV-9, or mock infected. Heat-inactivated serum samples were diluted 1:25 in wash buffer (0.05% Tween 20 in PBS) and used for ELISA. The absorbance value was read in a BioTek ELISA microplate reader at 405 nm. The S/P ratio was used to indicate the antibody level. Statistical significance was indicated by *p < 0.05, **0.001 < p < 0.05, and ***p < 0.001. Brackets above the bars indicate comparison between the ORF1stop group and wtFAdV-9 group. ELISA, enzyme-linked immunosorbent assay; FAdV, fowl aviadenoviruses; PBS, phosphate-buffered saline; S/P, sample to positive.
Cytokine gene expression in tissues
To determine whether FAdV-9 dUTPase modulates the host immune response in vivo as it does in vitro, the mRNA expression of cytokine genes for IFN-α, IFN-β, IFN-γ, IL-10, and IL-8 was investigated in the spleen, liver, and cecal tonsil by real-time PCR.
In spleens, treatment with all viruses resulted in an increase in the mRNA expression of IFN-α at 1, 3, 5, and 7 d.p.i., compared with the mock-infected group (Fig. 2). Infection with resORF1 and wtFAdV-9, but not ORF1stop showed significant upregulation of IFN-α expression at 1 and 3 d.p.i. (Fig. 2). At these time points, there was a significantly lower level of IFN-α expression for the ORF1stop compared with wtFAdV-9. No significant difference was found between any two groups at 5 and 7 d.p.i. (Fig. 2). For the mRNA expression of IFN-β in spleens, in spite of some increase in all virus-infected groups, no significant difference was found between any two groups at all time points.
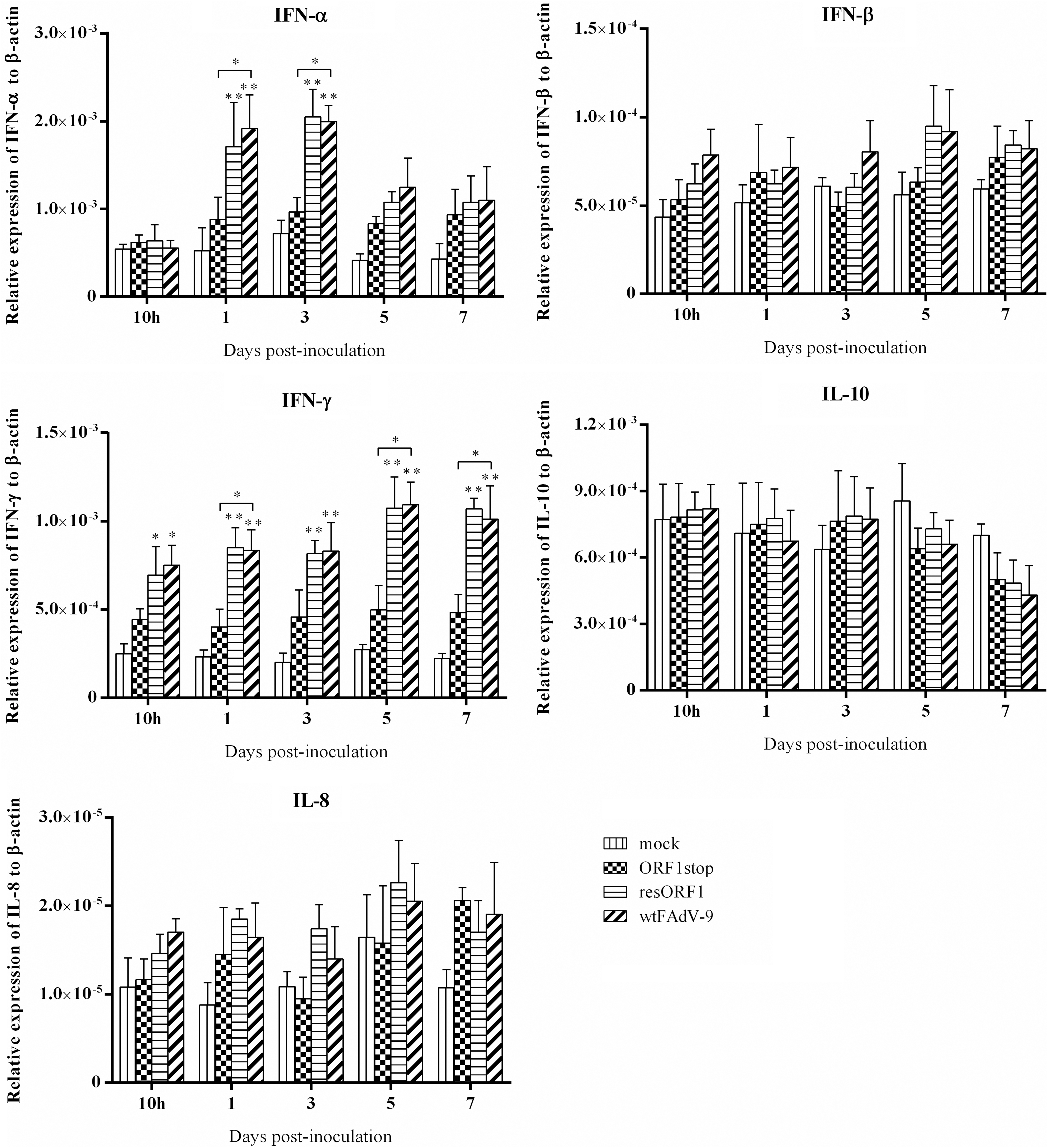
Cytokine mRNA expression in spleen samples of chickens. Chickens were orally inoculated with ORF1stop, or resORF1, or wtFAdV-9, or mock infected. The mRNA expression of target and reference genes was quantified by real-time PCR, and is presented relative to β-actin expression and normalized to a positive plasmid calibrator. Error bars represent SEMs. Statistical significance was indicated by *p < 0.05 or **0.001 < p < 0.05, compared with mock control. Brackets above the bars indicate comparison between ORF1stop group and wtFAdV-9 group. IFN, interferon; IL, interleukin; PCR, polymerase chain reaction; SEMs, standard error of the means.
Similar to IFN-α, the mRNA expression of IFN-γ was upregulated by all three viruses at all time points, compared with the mock control. An increase that was significantly different from time 0 was found in both the resORF1 and wtFAdV-9 groups throughout the study, but not in the ORF1stop group. There was a significant difference in the mRNA expression of IFN-γ between wtFAdV-9 and ORF1stop at 1, 5, and 7 d.p.i. as well with ORF1stop being lower. The mRNA expression of IL-10 was relatively stable in spleens, although increased expression was found at 3 d.p.i. and decreased expression was found at 5 and 7 d.p.i. in virus-infected groups, in comparison to mock-infected birds. No significant difference was found between any two groups for IL-10. In terms of the mRNA expression of IL-8, in spite of some upregulation by all three viruses, there was no significant difference between any two groups at any time point.
The mRNA expression of IFN-α, IFN-β, IFN-γ, IL-10, and IL-8 in liver samples is presented in Figure 3. All viruses induced higher IFN-α mRNA expression compared with mock control at 3, 5, and 7 d.p.i. At 5 d.p.i., a significant increase was found in both resORF1 and wtFAdV-9 groups, but not for ORF1stop. Similarly, the mRNA expression of IFN-β was upregulated by all the viruses throughout the study. There was a significant increase in both resORF1 and wtFAdV-9 groups, but not in ORF1stop at 5 d.p.i. Also, at this time point, significant difference was found between ORF1stop and wtFAdV-9. For IFN-γ, in spite of the increased mRNA expression induced by all viruses compared with mock control at all time points, wtFAdV-9 induced significantly higher level of IFN-γ mRNA than ORF1stop both at 3 d.p.i.
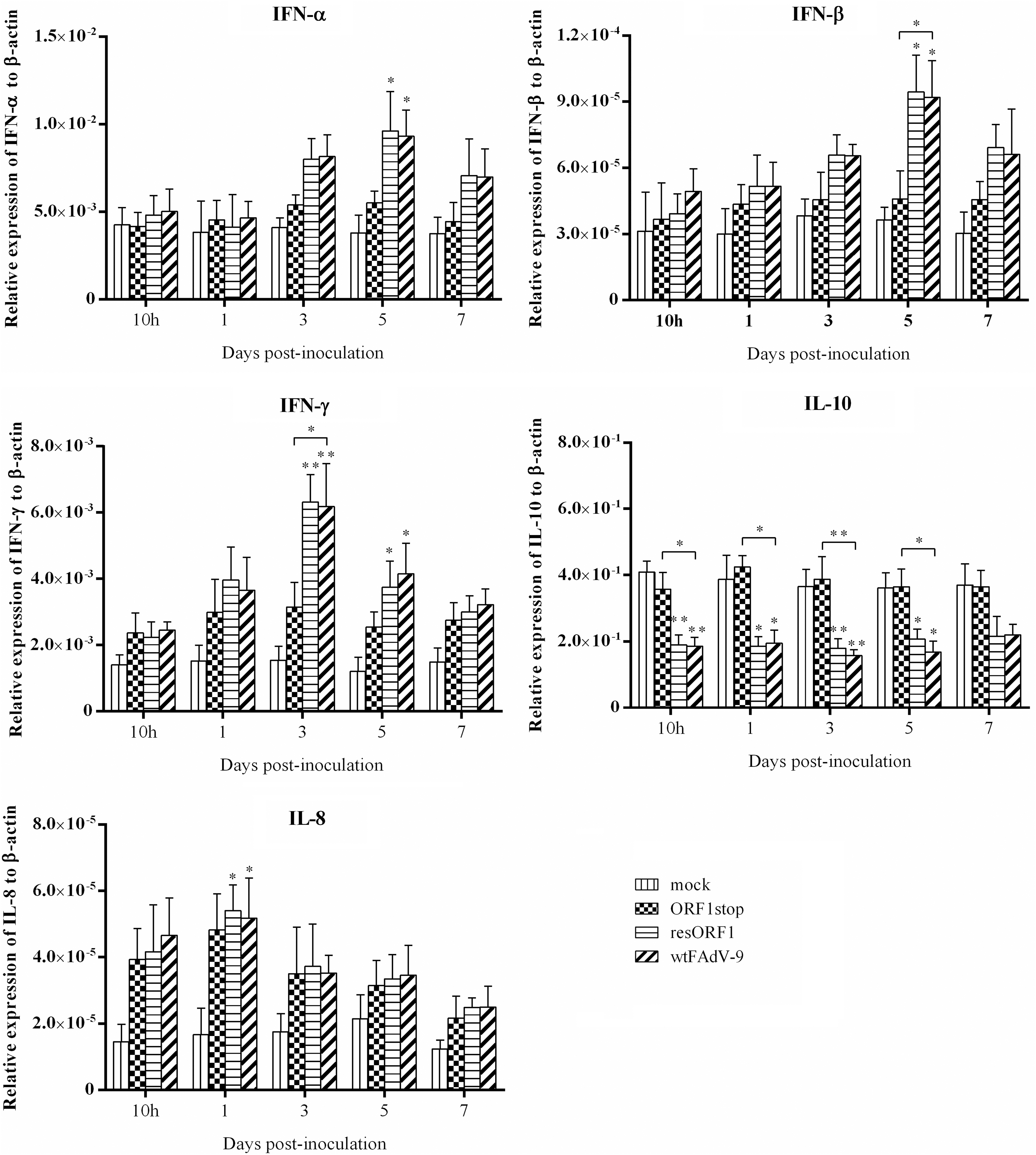
Cytokine mRNA expression in liver samples of chickens. Chickens were orally inoculated with ORF1stop, or resORF1, or wtFAdV-9, or mock infected. The mRNA expression of target and reference genes was quantified by real-time PCR, and is presented relative to β-actin expression and normalized to a positive plasmid calibrator. Error bars represent SEMs. Statistical significance was indicated by *p < 0.05 or **0.001 < p < 0.05, compared with mock control. Brackets above the bars indicate comparison between ORF1stop group and wtFAdV-9 group.
The production of IL-10 mRNA expression by ORF1stop was comparable to that induced by mock control at all time points. However, compared with mock control, there was a significant decrease of IL-10 mRNA expression in both resORF1 and wtFAdV-9 groups at 10 h.p.i., 1 d.p.i., 3 d.p.i., and 5 d.p.i. At 7 d.p.i., decreased IL-10 mRNA expression was also found in both resORF1 and wtFAdV-9 groups, but not in ORF1stop, but there was no significant difference between any two groups. The mRNA expression of IL-8 was upregulated by all viruses at all time points, compared with mock control. A significant increase was found only for both resORF1 and wtFAdV-9 groups, but not for ORF1stop at 1 d.p.i.
The mRNA expression of IFN-α, IFN-β, IFN-γ, IL-10, and IL-8 in cecal tonsil samples is presented in Figure 4. Compared with mock control, there was increased mRNA expression of all cytokines, including IFN-α, IFN-β, IFN-γ, IL-10, and IL-8 in all virus-infected groups throughout the study (except for a slight decrease for IFN-γ by ORF1stop at 1 d.p.i.). Although the difference was not significant, resORF1 and wtFAdV-9 had higher levels of IFN-α mRNA expression at 3, 5, and 7 d.p.i. than ORF1stop.
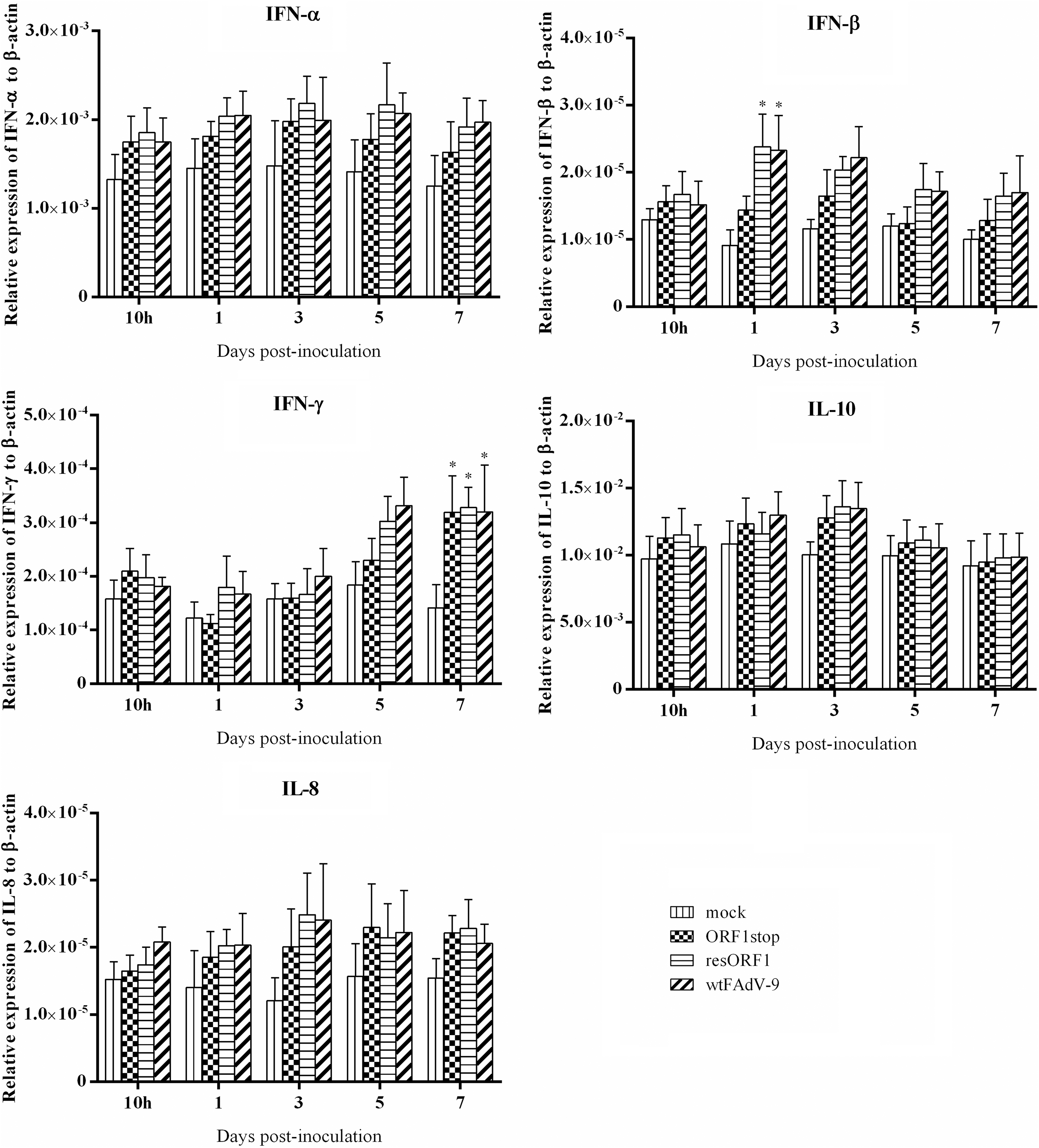
Cytokine mRNA expression in cecal tonsil samples of chickens. Chickens were orally inoculated with ORF1stop, or resORF1, or wtFAdV-9, or mock infected. The mRNA expression of target and reference genes was quantified by real-time PCR, and is presented relative to β-actin expression and normalized to a positive plasmid calibrator. Error bars represent SEMs. Statistical significance was indicated by *p < 0.05 compared with mock control.
Increased mRNA expression of IFN-β was found in all virus-infected groups. In addition, resORF1 and wtFAdV-9, but not ORF1stop showed a significant increase in mRNA expression of IFN-β at 1 d.p.i., compared with mock control. resORF1 and wtFAdV-9 had generally higher levels of IFN-β mRNA expression than ORF1stop at 1, 3, 5, and 7 d.p.i. Increased IFN-γ mRNA expression was found in all virus-infected groups at all time points (except for ORF1stop at 1 d.p.i.), while a significant increase was found only at 7 d.p.i., compared with the mock control. There was no significant difference regarding the mRNA expression of IL-8 between any two groups at all time points, despite some upregulation by all viruses.
Discussion
The early genes of human adenoviruses have been extensively studied (3,17,37), whereas only a few early genes have been functionally identified in FAdVs. We identified FAdV-9 ORF1 as a dUTPase enzyme, which is not required for virus replication, but is able to upregulate the expression of type I IFNs in CH-SAH cells and Celi cells (8). In the present study, we demonstrated that ORF1stop, the dUTPase knockout virus, did not replicate as efficiently as the wild-type virus at the early phase of infection (days 1 and 3 p.i.). This was evidenced by significantly less virus shedding and lower viral loads in tissues in the ORF1stop group compared with wtFAdV-9. However, no significant difference was observed between the two groups from 5 d.p.i. to the end of the study.
ORF1stop induced significantly less expression of IFN-α, IFN-β, IFN-γ, and significantly more expression of IL-10 in tissues, and elicited significantly lower antibody responses, compared with wtFAdV-9, suggesting important roles of dUTPase in virus replication in vivo and possibly in modulating the host immune response as well.
Several researchers have reported the roles of viral dUTPase in virus replication both in vitro and in vivo. For the in vitro studies, the most recognized conclusion is that the viral dUTPase is not essential in dividing cells, but is required for efficient replication in nondividing host macrophage cells. This was demonstrated for the dUTPase of African swine fever virus (26) and equine infectious anemia virus (34). We also found that FAdV-9 dUTPase did not affect virus replication in CH-SAH cells and Celi cells (8). However, the dUTPase of bovine immunodeficiency virus is critical for virus replication even in dividing cells, although viral cDNA synthesis was not reduced (35). On the other hand, different results were reported for the roles of viral dUTPases in vivo. Deletion of HSV dUTPase resulted in reduced replication of virus in the central nervous system of mice (29). However, the dUTPase of vaccinia virus is not required for virus replication in vivo (28).
In the present study, we showed that the replication of ORF1stop, the dUTPase knockout virus, was retarded in chickens at the early phase of infection (1 and 3 d.p.i.), but achieved the same level as wild type from 5 d.p.i. to the end of the experiment. This was demonstrated by the ORF1stop having significantly lower virus titer in feces and lower viral DNA genome copy number in tissues at early times p.i., compared with the wtFAdV-9. This suggests a role for FAdV-9 dUTPase in virus replication in vivo, at least at the early phase of infection. One possibility is that the absence of dUTPase in ORF1stop virus affects the viral DNA replication during the virus life cycle at the early phase of viral infection. After all, one of the major roles of dUTPase is to catalyze the cleavage of dUTP to dUMP and pyrophosphate and to maintain a low dUTP/dTTP ratio for accurate DNA replication.
However, the virus may adapt to recruit the cellular machinery and employ cellular dUTPase for effective viral DNA replication at later days. Interestingly, François et al. demonstrated that the transfection of the dUTPase-deficient CELO virus (FAdV-1) cosmid did not produce cytopathic effects (CPE) in chicken hepatocarcinoma cells, and the infection with the transfected cell lysates produced a CPE later than the wild-type CELO virus cosmid-transfected cell lysates (11). Together with our data, it is hypothesized that the lack of FAdV dUTPase partially retards virus replication.
The antibody level in wtFAdV-9-infected chickens increased throughout the experiment, which is consistent with the result of our previous study (9). Interestingly, the antibody level in ORF1stop-infected chickens was significantly lower than that in the wtFAdV-9 group in all time points, except for 7 d.p.i., which suggests that FAdV-9 dUTPase plays a role in the regulation of the host antibody response. This observation might be related to the ability of the FAdV-9 dUTPase to upregulate the expression of type I IFNs. In fact, it has been demonstrated that the production of type I IFNs is able to enhance the development of the Ab response against acute viral infections or vaccine antigens. For example, induction of endogenous type I IFNs potently enhances the primary antibody response through stimulating dendritic cells in vivo (19).
Fink et al. showed that type I IFNs-mediated signals on B cells specifically enhance antiviral humoral responses against vesicular stomatitis virus infection (10). Theofilopoulos and coworkers reported that type I IFNs can promote antibody-mediated immune responses through promoting adaptive T and B cell responses (33). Similarly, Zhu et al. demonstrated that upon adenoviral infection, type I IFN signaling through both B and CD4 T cells is required for the production of virus-specific IgG2a and IgG2b isotypes and protective neutralizing antibody (38). However, further experiments involved in testing B and CD4T cells are needed to determine whether the lower antibody level induced by ORF1stop is directly associated with the ability of FAdV-9 dUTPase to regulate type I IFNs.
Previously, we demonstrated that despite inducing a significantly higher level of antibody, compared with the mock control, a multiORFs-deleted virus FAdV-9Δ4, which lacks ORFs 0, 1, 1A, 1B, 1C, and 2, induced significantly lower level of FAdV-specific antibodies than the wild-type virus (5,8). Together with our current data, this suggests that of the six deleted genes, dUTPase, if not the only one that is associated with the mediation of the antibody response. Additional gene knockout studies that are ongoing in our laboratory will help determine if there are other genes besides ORF1 that are involved in the mediation of the humoral immune response.
Among numerous innate immune cytokines, type I IFNs are pleiotropic cytokines that are capable of exerting a direct antiviral immune response against virus infection. Induction of type I IFNs is part of the innate immune response to adenovirus infection. We previously found that FAdV-9 dUTPase is important for the upregulation of type I IFNs in CH-SAH cells and Celi cells. In this study, the mRNA expression of IFN-α and IFN-β was investigated in spleens, livers, and cecal tonsils. Infection with the dUTPase knockout virus, ORF1stop, resulted in significantly less expression of IFN-α mRNA in spleen at 1 and 3 d.p.i., and significantly less expression of IFN-β mRNA in liver at 5 d.p.i., compared with the wild-type virus. This indicates that the role of FAdV-9 dUTPase in upregulating type I IFNs is not limited to an in vitro situation, but also applies in vivo.
In our in vitro study, in which both CH-SAH cells and Celi cells were infected with ORF1stop or wtFAdV-9, no significant difference was found in the mRNA expression of IFN-γ between these two groups. However, the in vivo study showed that there was significantly less expression of IFN-γ mRNA in livers of the ORF1stop group than that of wtFAdV-9 group at 3 d.p.i.; as was the case in spleens at 1, 5, and 7 d.p.i. This could be because organs such as spleen and liver have more types of cells capable of producing INF-γ, compared with a single type of cell line.
IL-10 plays a pleiotropic role in the modulation of the immune response. Although IL-10 is known predominantly as an anti-inflammatory cytokine by suppressing the activity of Type 1 T helper (Th1) cells, NK cells, and macrophages (7), it has been also demonstrated to be associated with virus clearance, including influenza virus, hepatitis B virus, hepatitis C virus, and human immunodeficiency virus (31).
In the present study, significant downregulation of IL-10 mRNA expression was noticed at 10 h.p.i., 1, 3, and 5 d.p.i. in livers of virus-infected groups, compared with that of the mock-infected group. This is consistent with our previous in vitro data (8). In addition, the mRNA expression of IL-10 induced by ORF1stop was significantly lower than that of the wtFAdV-9 and resORF1 groups. Perhaps one of the mechanisms that FAdV-9 recruits to escapes from the host clearance of virus is to downregulate the IL-10 expression. Furthermore, we hypothesize that ORF1 plays an important role in the downregulation of IL-10.
IL-8 is an important mediator of the inflammatory response to many stimuli, including viruses, which is responsible for the recruitment of neutrophils that participate in the local inflammatory infiltrate, contributing to airway closure (2). Many viruses can induce the IL-8 expression, including respiratory syncytial virus, vaccinia virus, herpes virus, and adenovirus (22).
In the present study, we also sought to explore the IL-8 expression by FAdV-9 infection. Despite a minor trend of upregulation, there is no significant difference between the virus-infected groups and mock-infected group in all tissues throughout the study, except in liver at 1 d.p.i. This observation is contrary to one of our previous studies where we demonstrated that FAdV-8 downregulates the IL-8 expression (13). Recent studies have shown that the induction of IL-8 by adenovirus is type specific (20). For example, human adenovirus 7 and 19, but not 5 can stimulate the IL-8 expression (20,30).
Overall, our data demonstrated that deletion of FAdV-9 dUTPase retarded virus replication at the early stage of the infection and was associated with modulating the host immune response against virus infection as well. We believe our study provides useful data that will shed more light on the mechanism of the host immune response against the FAdV infection.
Footnotes
Acknowledgments
This work was supported by the Natural Sciences and Engineering Research Council of Canada, the Canadian Poultry Research Council, and the Ontario Ministry of Agriculture, Food, and Rural Affairs. L.D. was a recipient of China Scholarship Council PhD fellowship. The authors thank the personnel of the Isolation Unit for their professional animal care and assistance.
Author Disclosure Statement
No competing financial interests exist.