
Abstract
Due to the limitations and safety issues of the two currently approved live attenuated rotavirus (RV) vaccines “RotaTeq and Rotarix,” studies on nonreplicating sources of RV vaccines and search for proper RV antigens are actively carried out. The adjuvant activity of NSP4 and highly immunogenic properties of RV VP6 protein prompted us to consider the construction of a NSP4112-175-VP6 fusion protein and to assess the anti-VP6 IgG, IgA, and IgG subclass responses induced by Escherichia coli-derived NSP4-VP6 fusion protein compared to that of VP6 protein with/without formulation in Montanide ISA 50V2 (M50) in BALB/c mice. Results indicated to the proper expression of the fused NSP4-VP6 and VP6 proteins in E. coli. Intraperitoneal immunization by M50 formulated NSP4-VP6 fusion protein (M5+NSP4-VP6) induced the highest titration of VP6-specific IgG and IgA responses compared to the other groups. Indeed, the presence of NSP4 resulted to the induction of stronger humoral immune responses against the fused protein compared to that elicited by administration of VP6 protein alone (with/without M50 formulation), implying the adjuvant properties of NSP4 for the fused protein. Moreover, the “M50+NSP4-VP6” formulation induced higher serum IgG2a titers than IgG1 and increased Interferon-γ levels, despite unchanged interleukin-4 amounts compared to other groups, indicating Th1-oriented responses with a possible role of NSP4. In conclusion, this study further highlights the potentiality of NSP4-VP6 fusion protein as an efficient and cost-effective immunogen in the field of RV vaccine development.
Introduction
R
Over the past decades, the RV vaccine development has been encouraged due to high morbidity and mortality in children. In this context, two live attenuated RV vaccines, “RotaTeq (a pentavalent RV vaccine containing five human–bovine reassortant RV strains: G1-G2-G3-G4 P[5] and G6P[8]) and Rotarix (a monovalent vaccine derived from the most common human RV strain G1P[8])” have been recently licensed in many countries and shown to protect efficiently against RV severe diarrhea (3,50,54,55). Despite their high efficacy in the developed world, both vaccines show lower efficacy in developing countries. Moreover, safety issues, including a low risk of intussusception and the risk of reassortment between the vaccine and a wild-type strain, are serious concerns associated with the live attenuated RV vaccines (1,40,62). To address these concerns, a new paradigm toward nonreplicating RV vaccines, including inactivated RV strains, DNA vaccines, virus-like particles (VLPs), and VP6-based subunit vaccines, were introduced (29,34,42,46,57,59,61). Among nonreplicating vaccine approaches, VP6-based RV vaccine candidates have provided promising results. VP6 protein of RV is highly conserved among different types, abundant (in the virus structure) and is immunogenic. It has been reported that VP6 is capable of self-assembling into trimers, nanotubes, nanospheres, and sheets that can provide an excellent immunogenic target (7,9,25,39). VP6-based vaccine candidates induce protective antibodies (Abs) even without any neutralizing (NT) activity (10,20,26). In fact, non-NT IgA Abs against VP6 are capable of clearing chronic RV infections, while both IgG and IgA Abs were further shown to protect suckling pups born from immunized dams against the virulent RV challenge and severe acute diarrhea (33,58).
NSP4, an integral endoplasmic reticulum membrane glycoprotein, plays a role in RV morphogenesis of RVS. NSP4 exhibits enterotoxic activities that might have contribution in dose- and age-dependent diarrhea in infant mice. In this context, Abs against NSP4 may play a significant role in prevention of diarrhea post-RV infection (15,48,56). More recently, a new characteristic was attributed to NSP4, which was due to its adjuvant effect to enhance the immune responses against other fused/combined proteins. In this context, adjuvant properties of NSP4 to enhance the humoral immune responses against coadministered model immunogens such as keyhole limpet hemocyanin, tetanus toxoid, or ovalbumin antigens were reported. Moreover, NSP4 localized within a VLP2/6 also exhibited adjuvant effects. In particular, results of prior studies suggested that both full-length and a cleavage product of SA11 NSP4 (112–175) can exert similar adjuvant activities, localizing this adjuvant activity to the C-terminus of the NSP4 protein (36). In line with these findings, plausible interaction with TLR-2 triggering the secretion of pro-inflammatory cytokines and dominant Th1 lymphocyte immune response due to increased serum IgG2a antibodies is attributed to the adjuvant activity of NSP4 (18,28).
In the present study, we investigated the potential adjuvant properties of NSP4 for VP6 immunization. To this end, mice were immunized intraperitoneally by an E. coli-derived NSP4112-175-VP6 fusion protein or sole VP6 with/without formulation in Montanide ISA 50V2 (M50) adjuvant, and the status of VP6-specific immune responses in sera of immunized mice was compared. In addition to humoral responses, cytokine profile was further studied in immunized mice with VP6 fusion protein or sole VP6 with/without adjuvant.
Materials and Methods
Construction of VP6 expression vectors
The complete 192 nucleotides of NSP4 (residues 112–175) of RV SA11 strain (accession no. AF087678.1) harboring a flexible “glycine-serine [(Gly)4Ser]3 linker” (12) at 3′ of the NSP4 with flanking restriction sites at 5′ (NcoI) and 3′ (SalI and XhoI, respectively) were synthesized (Generay Biotech Co., Ltd., China) and delivered in pUC57 vector (pUC57/NSP4).
Amplification of the complete open reading frame (ORF) of VP6 gene from RV Wa strain (52) was performed using reverse transcription polymerase chain reaction (RT-PCR), with the following primer's pair: VP6-F 5′-ACGC
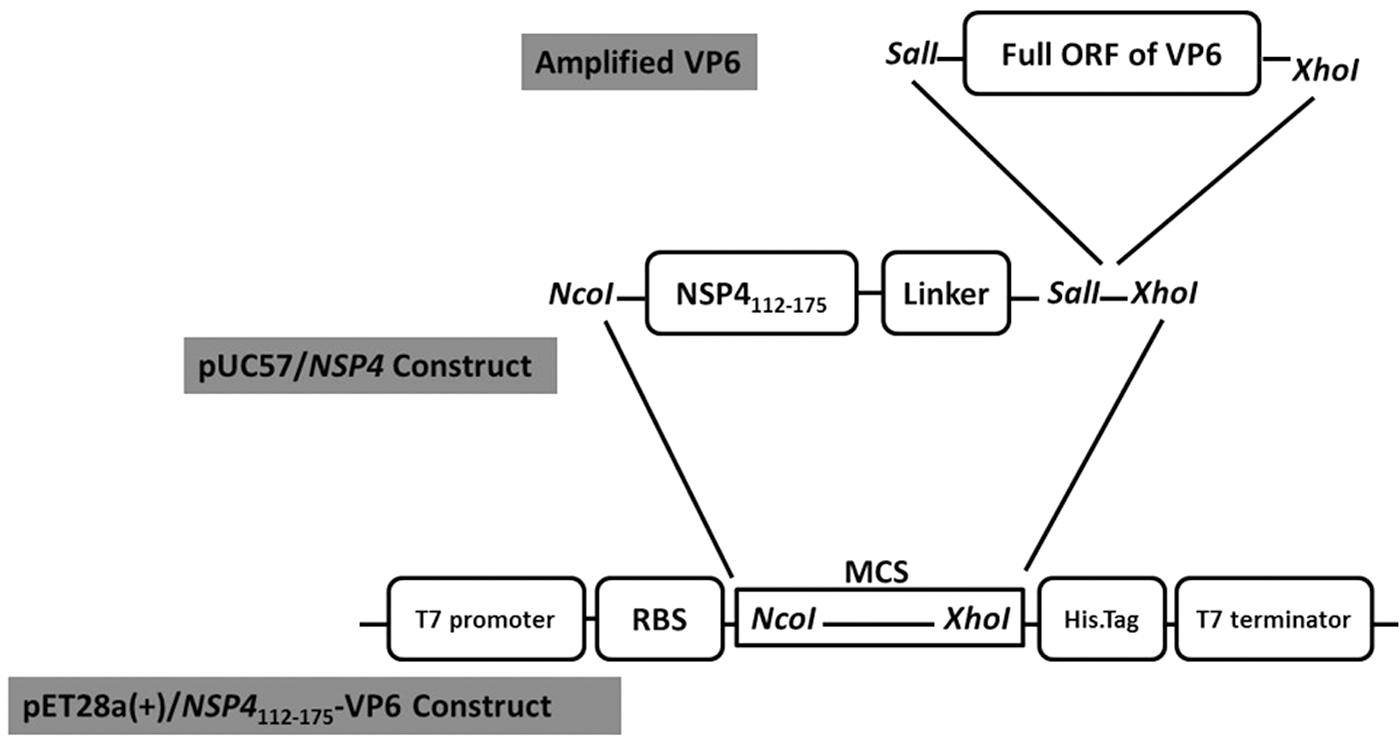
Construction of pET28a(+)/NSP4112-175-VP6. Amplified full ORF of VP6 was used to clone into SalI and XhoI sites of pUC57/NSP4 plasmid. Recombinant pUC57/NSP4-VP6 plasmid was digested with NocI and XhoI restriction enzymes to remove the NSP4-VP6 fragment and subcloned into NocI and XhoI sites of pET28a(+) vector. MCS, multiple clone sites; ORF, open reading frame; RBS, ribosomal binding site.
Protein expression, purification, and analyses
The recombinant pET28a(+)/NSP4-VP6 and pET28a(+)/VP6 were used to transform the E. coli BL21 strain (DE3; Novagen) as expression hosts. The transformed bacteria were incubated in Luria-Bertani (LB) broth (containing 50 μg/mL of ampicillin) at 37°C until the cell density reached OD600 nm: 0.6–1. The expression was performed in BL21 cells by 3 h induction with 1 mM isopropyl β-D-1-thiogalactopyranoside (IPTG). Cells were centrifuged at 10,000 g for 2 min and lysed with Lysis Buffer (50 mM Tris–HCl pH 8, 5 mM ethylenediaminetetraacetic acid (EDTA), 10% glycerol, 0.5% NP40, and 50 mM NaCl). The cytoplasmic fraction was recovered by centrifugation at 10,000 g for 2 min. Subsequently, the bacterial pellet containing insoluble fraction of the expressed proteins (considered as inclusion bodies) was solubilized with 8 M urea according to standard procedures.
Expression of the recombinant proteins was analyzed and confirmed by western blotting. To this end, the cytoplasmic fraction and solubilized inclusion bodies were suspended in gel loading buffer, boiled for 5 min, separated by 10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE), and transferred onto a polyvinylidene difluoride (PVDF) membrane (Roche, Germany). Membranes were blocked by 5% nonfat milk and incubated with His-probe antibody (Santa Cruz Biotechnology, Inc., CA) (1:1,000 dilution in blocking solution) at room temperature for 4 h. After several washing steps of the membranes with PBST (phosphate-buffered saline containing 0.1% Tween 20), secondary Polyclonal Rabbit Anti-Mouse Immunoglobulins/horseradish peroxidase (HRP) (1:1,000 dilution) (Dako, Denmark) were added for 2 h at room temperature. The membranes were finally washed and developed by DAB (3,3′-diaminobenzidine tetrahydrochloride) (Sigma, Germany). The gel Electro Eluter was used to purify expressed proteins according to the manufacturer's instructions (Bio-Rad Laboratories, Hercules, CA) and used for immunization experiments in mice. The concentration of purified proteins was determined by the Bradford method (Bio-Rad Laboratories) using bovine serum albumin (BSA) as standard. The purified proteins were further verified by western blotting as described above, except for using
Animal experiments
Immunization protocol
To evaluate the induction of immune responses by purified RV NSP4-VP6 fusion protein or VP6 protein, BALB/c mice 6–8 weeks of age were randomly divided into six groups (groups 1–6, each group with 5 mice) and injected intraperitoneally with 20 μg purified proteins. Groups 1 to 4 received IP injections of 100 μL of NSP4-VP6 fusion protein and VP6 protein either alone or formulated with M50 (50:50 water-in-oil based on volume), respectively, according to the manufacturer's instructions (Seppic Adjuvants, France). As the negative controls, mice in group 5 and 6 received IP injections of 100 μL of sterile PBS and adjuvant (M50) alone, respectively. The immunization schedule was based on a three-dose regimen, where the booster inoculations were administered at 2 and 4 weeks after the primary injection. Blood samples were drawn from the retro-orbital vein 7 days after last administration, and the mice were then sacrificed according to established guidelines. The approval for the experiments was confirmed by the Local Ethics Committee of Pasteur Institute of Iran. All experiments were performed according to the Helsinki guidelines.
Serological assessment
Sera collected from immunized and control groups were tested for VP6-specific IgG, IgA antibodies, and IgG1, IgG2a subclasses by enzyme-linked immunosorbent assay (ELISA). To this end, Nunc-Immuno F8 MaxiSorp™ plate (Thermo Scientific, Rochester) was coated with 2 μg/mL of recombinant VP6 protein in PBS at 4°C overnight (100 μL/well) and blocked with 1% BSA. Fourfold dilutions of heat-inactivated mouse sera (starting at a 1:250 dilution) were added to each well and incubated for 1 h at 37°C. HRP-conjugated goat anti-mouse IgG (1:5,000 dilution) (Abcam, Cambridge, United Kingdom), IgA (1:5,000) (Abcam, Cambridge, United Kingdom), and IgG1 or IgG2a (Santa Cruz Biotechnology, Inc.) were used to detect total IgG, IgA antibodies, and IgG subclasses. Reactions were developed with 3,3′,5,5′-tetramethylbenzidine (TMB) solution, stopped with 2 N sulfuric acid, and absorbance was read at 450 nm. Positive reactions were determined when the mean absorbance +3 standard deviations exceeded the equal dilutions of control sera.
Cytokine measurement
Spleens were removed from immunized mice at 1 week after last administration. Briefly, single cell suspensions were prepared by mechanical dissociation in complete medium. Cells were added to 24-well plates (Thermo Scientific) in duplicate at a density of 5 × 105 cells per well and cultured with 5 μg/mL of recombinant protein. The cultures were incubated for 4 days at 37°C under 5% CO2. The cell culture supernatants from the wells were then pooled and stored at −70°C for cytokine assays. Interferon-γ and interleukin (IL)-4 were assessed using mouse interferon-γ and IL-4 ELISA Development Kit (Mabtech, Nacka Strand, Sweden), according to the manufacturer's instructions.
Statistical analysis
All the experiments were performed thrice and in triple each time. Statistical analysis was performed with GraphPad Prism Software Version 5 (GraphPad Software, Inc., San Diego, CA). The nonparametric test, one-way ANOVA (Tukey's Multiple Comparison Test), was used to analyze humoral immune responses. p Value <0.05 was considered statistically significant.
Results
Expression of NSP4-VP6 fusion protein and VP6 protein
The expected amplicons of NSP4-VP6 fusion protein and VP6 protein were obtained and cloned into pET28a(+) vector. Colonies carrying recombinant pET28a(+)/NSP4-VP6 and pET28a(+)/VP6 were isolated. Positive colonies were then confirmed by colony PCR and recombinant pET28a(+) vectors further verified by release of a DNA fragment of the expected length using restriction enzyme digestion (Fig. 2). Sequence analysis verified the correct in-frame integration of the gene downstream of the T7 promoter of recombinant pET28a(+) vectors.
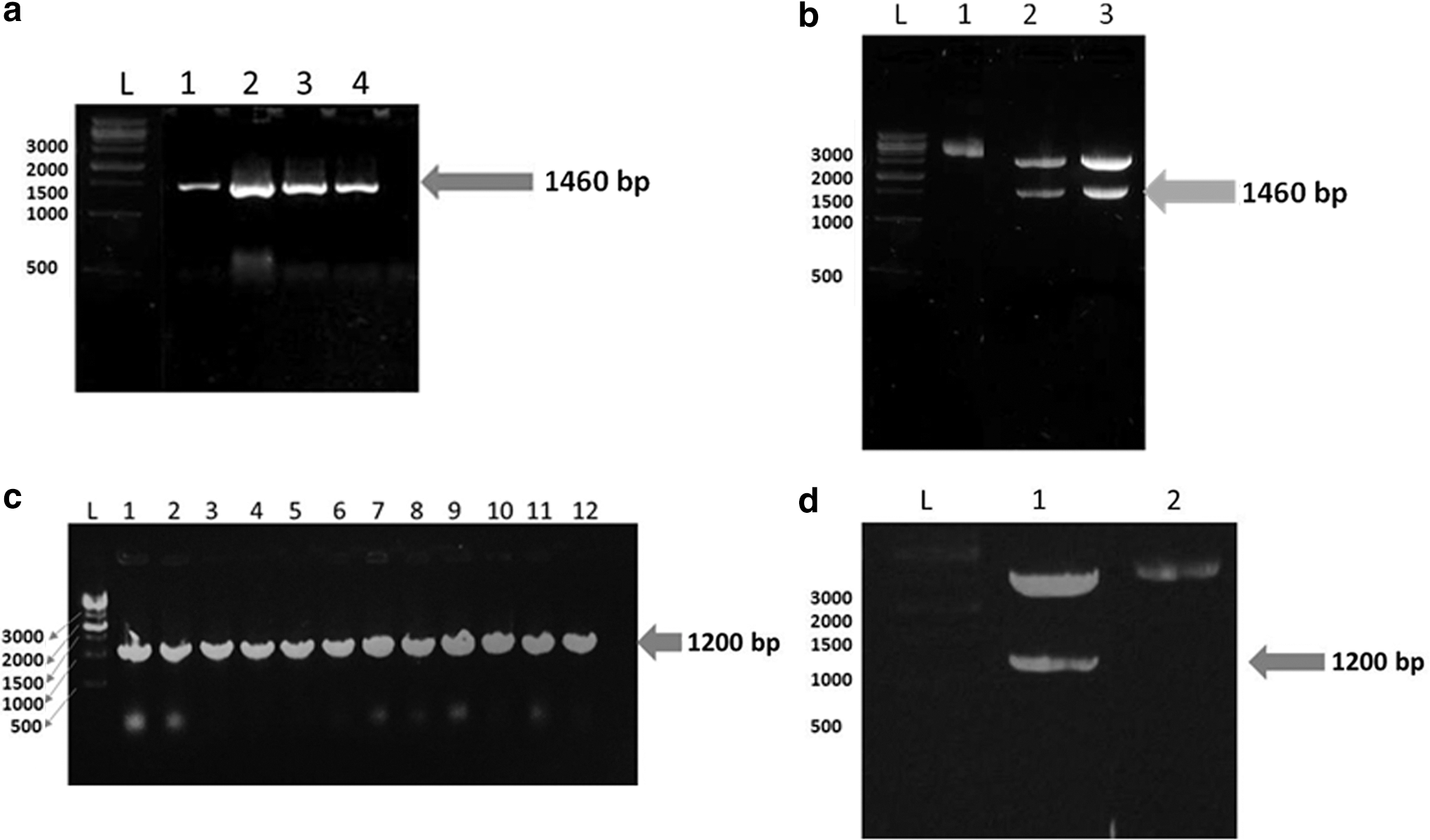
Confirmation of recombinant pET28a(+)/NSP4-VP6 and pET28a(+)/VP6 constructs using colony PCR and restriction enzyme digestion.
The expression of NSP4-VP6 fusion protein and VP6 protein was analyzed after 3 h induction with 1 mM IPTG in both cytoplasmic fraction and inclusion bodies. Total proteins were then separated by 10% SDS-PAGE and transferred onto the nitrocellulose membrane. As shown in Figure 3a, the presence of NSP4-VP6 fusion protein (54 kDa) and VP6 protein (44 kDa) in inclusion bodies was verified by western blot analysis using His-probe antibody. Proteins were also detected in cytoplasmic fraction, but in very low concentrations (the soluble cytoplasmic fraction). Moreover, both purified proteins (NSP4-VP6 and VP6) were further confirmed by western blot using RV capsid antibody (Fig. 3b).

Expression of NSP4-VP6 fusion protein and VP6 protein in BL21 Escherichia coli.
Serum antibody responses
To compare the immunogenicity of NSP4-VP6 fusion protein and VP6 protein, mice were immunized IP with three doses of antigens with or without M50 adjuvant formulation. Immunized and control mice were daily monitored during the immunization and did not show any clinical signs of toxicity. Sera collected from mice analyzed by ELISA and different levels of total IgG (p < 0.0001) and IgA (p < 0.0001) responses against VP6 were observed in each immunized group, but remained negative in control mice (Fig. 4a, b). As shown in Figure 4a, significant anti-VP6 IgG titers were induced following immunization with VP6 protein and NSP4-VP6 fusion protein, reaching from highest titers of 1:1,000 (p < 0.001) to 1:16,000 (p < 0.05), respectively. Accordingly, antigens formulated with M50 adjuvant also induced significant anti-VP6 IgG titers, reaching from highest titers of 1:4,000 (p < 0.001) for VP6 protein to 1:64,000 (p < 0.05) for NSP4-VP6 fusion protein. Comparison of end-point titration of two antigens clearly supports the potentiality of NSP4-VP6 fusion protein for induction of higher levels of anti-VP6 IgG antibodies. Indeed, immunization by NSP4-VP6 fusion protein either alone or formulated with M50 elicited high titers of anti-VP6 IgG compared to the same adjuvant formulation for VP6 protein (1:1,000, p < 0.05 and 1:4,000, p < 0.05, respectively).
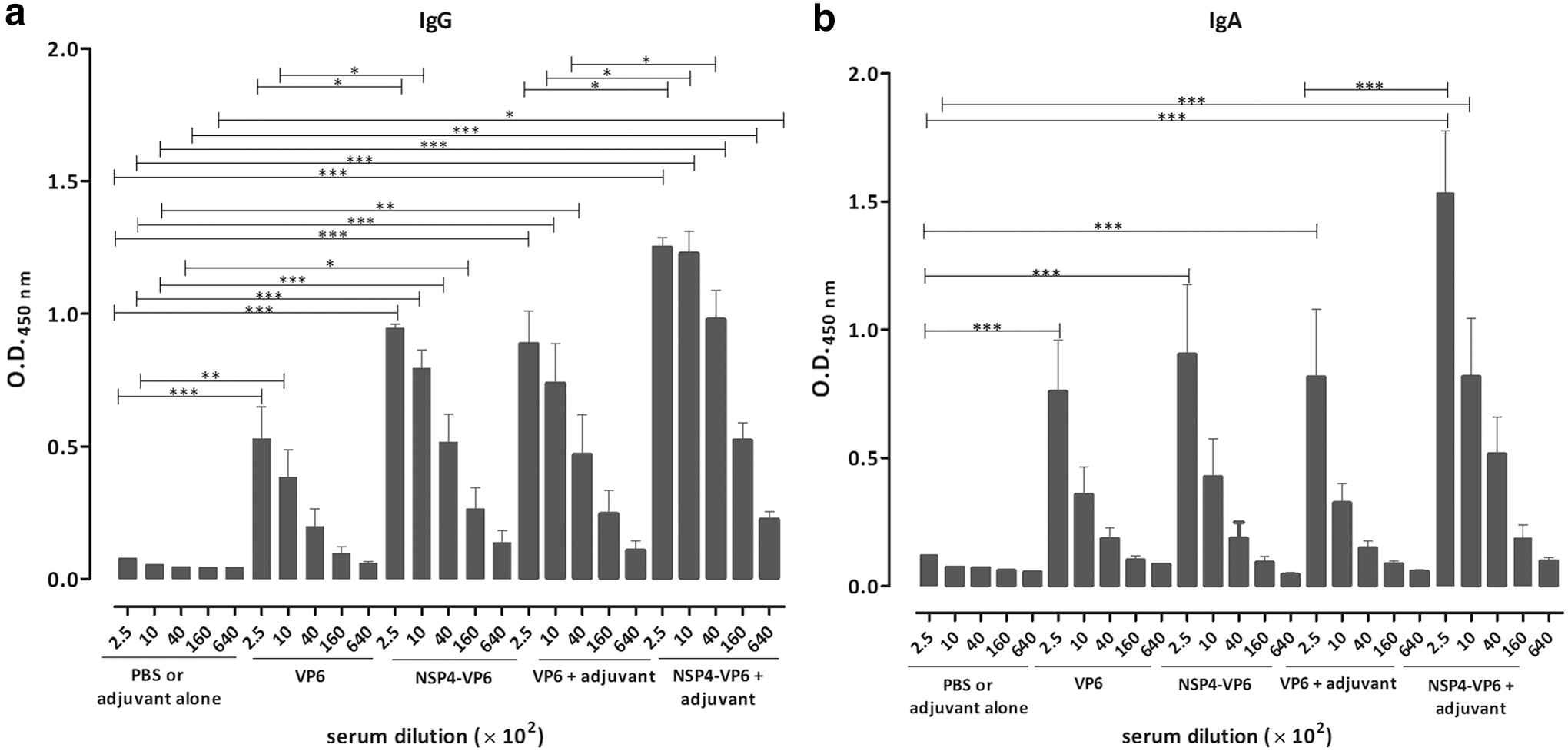
Anti-VP6 IgG
As shown in Figure 4b, immunization with VP6 protein and NSP4-VP6 fusion protein elicited significant induction of anti-VP6 IgA titers, reaching to the similar titers of 1:250 (p < 0.001). Similarly, immunization of these proteins in formulation with M50 also induced significant anti-VP6 IgA titers, reaching from highest titers of 1:250 (p < 0.001) to 1:1,000 (p < 0.001), respectively. Moreover, end-point titration for NSP4-VP6+M50 immunized group showed significant anti-VP6 IgA titers (1:250, p < 0.001) compared to VP6+M50. However, the differences for IgA were not statistically significance for groups immunized by antigens without adjuvant. In addition, VP6-specific IgG subclass titers for IgG1 (representing a Th2 response) and IgG2a (representing a Th1 response) were also measured. The level of IgG1 and IgG2a subclass titers revealed that all immunization groups produced both IgG1 and IgG2a subclass antibodies. Ratio of IgG2a/IgG1 for immunized groups indicated that the IgG2a/IgG1 ratio was higher in mice immunized with NSP4-VP6+M50 than other immunized groups (Fig. 5).
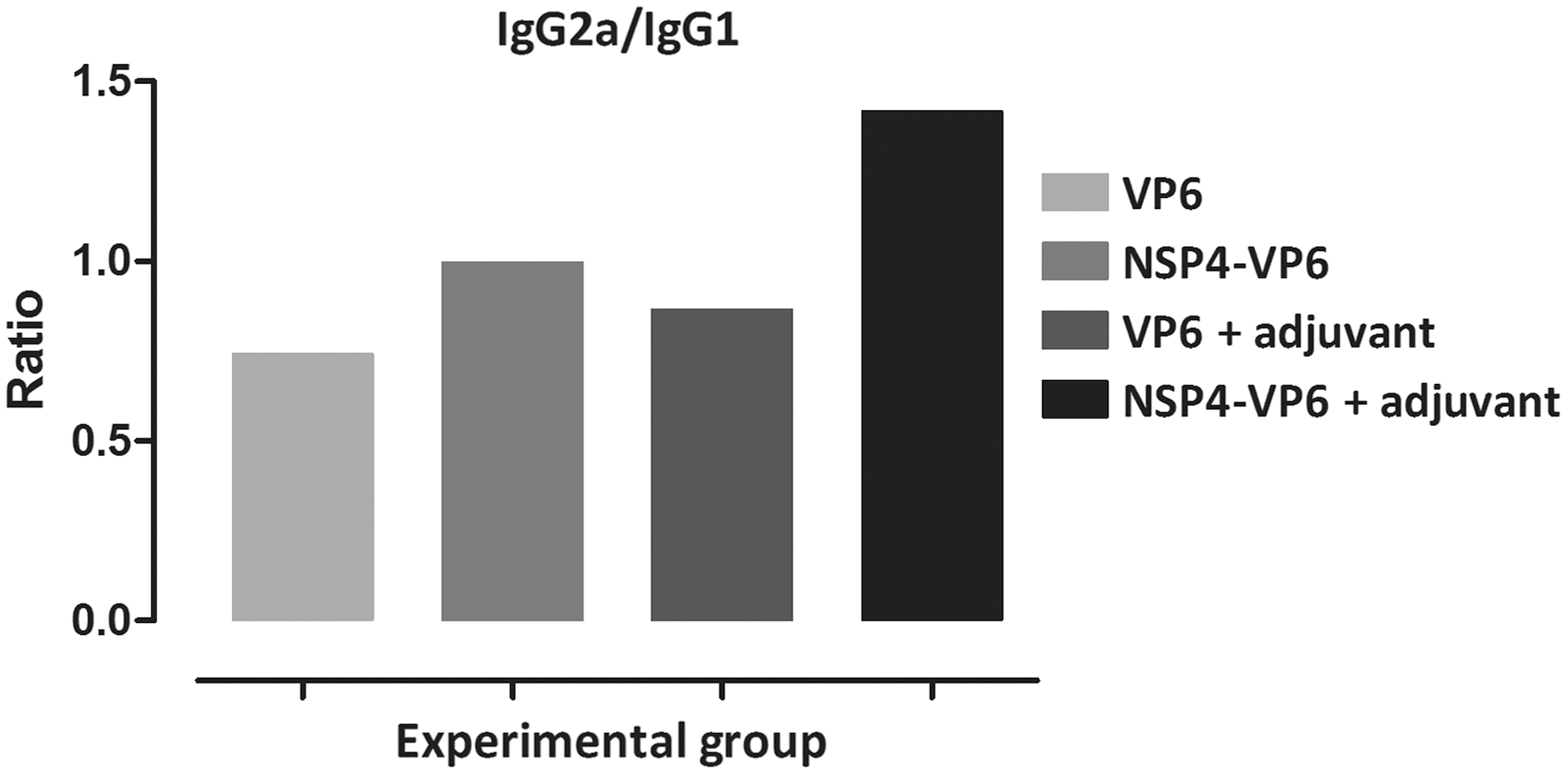
Serum IgG2a/IgG1 ratio in BALB/c mice immunized intraperitoneally. Antibody isotyping was performed by ELISA using HRP-conjugated goat anti-mouse IgG1 and IgG2a. The IgG2a/IgG1 ratio was calculated from the mean OD values obtained from different serum dilutions. Since no antibody could be detected in PBS group, the IgG2a/IgG1 ratio was not determined for this group. ELISA, enzyme-linked immunosorbent assay; HRP, horseradish peroxidase; PBS, phosphate-buffered saline.
Cytokine measurement
The levels of Interferon-γ and IL-4 (as a representative of Th1 and Th2 cell responses, respectively) were measured upon in vitro stimulation of spleen cells. As shown in Figure 6, higher levels of Interferon-γ were produced in NSP4-VP6 fusion protein+M50 group compared to other immunized groups (p < 0.001). Although statistically nonsignificant, NSP4-VP6 alone also showed increased level of Interferon-γ. Moreover, the level of IL-4 was found to be unchanged comparing all groups (data not shown).
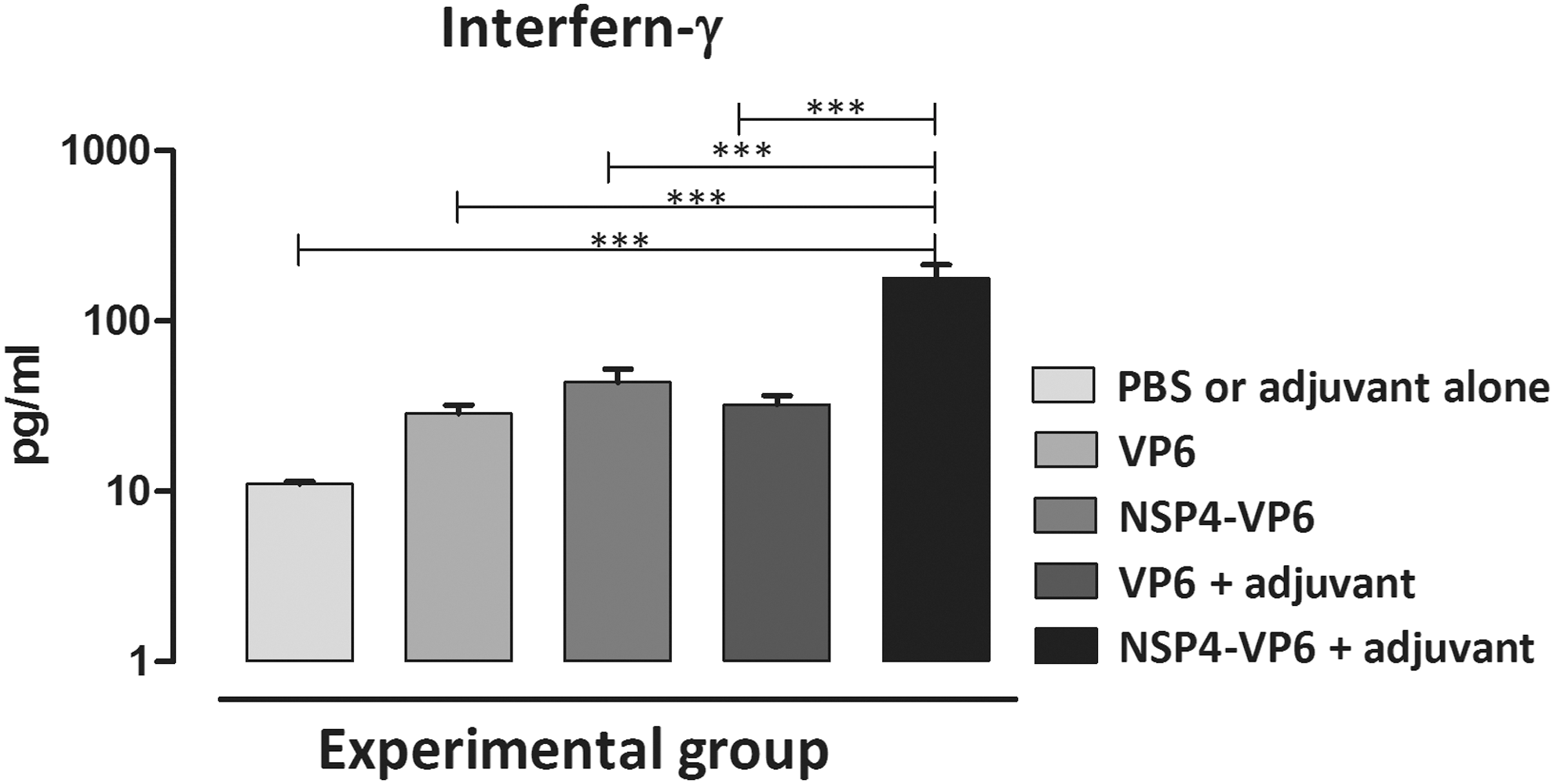
Interferon-γ level at 1 week after last immunization. Interferon-γ concentration was assessed in the supernatant of splenocytes of mice per group that were stimulated. Bars represent the mean ± standard error of the mean (SEM) (one-way ANOVA, with Tukey's Multiple Comparison Test). SEM, standard error of the mean (***p < 0.001).
Discussion
RotaTeq™ and Rotarix™ were introduced and incorporated as part of routine childhood national immunization programs in more than 80 countries (32). Although showed some promising results, raised concerns of the attenuated RV vaccines skewed researchers' focus on alternative RV vaccine candidates such as nonreplicating RV vaccines. The capability of VP6 in inducing protection against all group A RVs makes it a potential universal nonliving RV vaccine candidate. This notion is supported by several studies indicating the immunogenicity of VP6 protein using different platforms of RV VP6 such as VP6 DNA vaccine (11,14,17,30,31,60), VP6 purified from the RVs, VP6 recombinant protein (15,16,45), maltose binding protein-VP6 fusion protein (13), and trimeric, spherical, and tubular structures of VP6 (7,37 –39,41,47). Results obtained from previous studies suggested different levels of protection against RV challenge dependent on different adjuvant or administration route, highlighting the necessity of an efficient adjuvant to improve protection induced by recombinant VP6 protein. Among several adjuvants studied, the cholera toxin (CT) and the heat-labile enterotoxin (LT) together with different VP6 constructs have shown an important role in protection against RV challenge (15 –17,43,44,53). RV NSP4, as a viral enterotoxin, has also been recently shown that it can function as an adjuvant to enhance immune responses (36).
In the present study, NSP4 (112–175) was genetically fused to VP6 protein and RV NSP4-VP6 fusion protein could be efficiently expressed in bacterial host. In agreement with previous studies, our results verified preferential aggregation of the proteins into inclusion bodies by E. coli system (21,51). The NSP4-VP6 fusion protein elicited high humoral immune responses in immunized BALB/c mice, indicating the possibility of using NSP4-VP6 fusion protein as an efficient and cost-effective immunogen protein.
The humoral RV immune responses are crucial in protecting from RV AGE, and several studies support the correlation of antibodies in the form of IgG and IgA with protection and viral clearance in the course of natural infection or vaccination (8,23,49). The important role of T cell responses in protection against RV infection was highlighted by several VP6-based vaccine studies although antibody response against VP6 is shown to be important in protective immunity in some models. Indeed, fecal IgA levels to VP6 were reported to be correlated with protection. There are also some reports indicating that non-NT IgA Abs against VP6 are able to clear chronic RV infections, which suggest intracellular inhibition of RV by IgA following transcytosis (13,33,45,58). Therefore, for an enhanced protection, it is important to improve the anti-VP6 IgG and IgA responses induced by VP6-based vaccines. Indeed, different levels of serum IgG and IgA responses against VP6 were found in each immunized group compared to control groups. In particular, M50 formulated NSP4-VP6 fusion protein was found to efficiently elicit the serum IgG and IgA against VP6. Interestingly, this formulation, induced a 2- and 1-titration rise in anti-VP6 IgG and IgA response compared to VP6+M50 formulation, suggesting a progressive increase of specific serum anti-VP6 IgG and IgA in BALB/c mice. Data obtained from immunization by NSP4-VP6 fusion protein indicated that a significant raised titration against VP6 could be mediated by the presence of NSP4. Moreover, IgG subclass analyses showed that the NSP4-VP6 fusion protein could induce higher serum IgG2a titers than IgG1. In line with our finding, Choi et al., have shown that immunization with chimeric VP6 alone or adjuvanted with LT (R192G) or CTA1-DD or Adjumer induced titers of the two IgG subclasses but the IgG1/IgG2a ratio skewed response toward a Th2-like response. They also suggested that chimeric VP6 adjuvanted with CpG oligodeoxynucleotides (CpG ODN) can increase the IgG1 and IgG2a titers toward a balanced Th1/Th2-like response (13,16). In contrast to this finding, our study indicated that immunization with NSP4-VP6+M50 can induce Th1-like response compared to VP6+M50. However, skewing toward either IgG subclass does not appear to be critical in protection but overall IgG responses are supposed to play an important role in protection against RV. It seems that both Th1 and Th2 cells elicit protective responses following immunization with adjuvants (13,15,16).
Both anti-NSP4 IgG and IgA antibody responses are shown to be induced during natural RV infection in children (48). It has been also shown that NSP4 can induce higher serum IgG2a titers than IgG1 (18,19), indicating a shift toward Th1-like immune responses, as well as adjuvant properties (36). In agreement with previous studies, our data also suggested that NSP4 protein not only could enhance humoral response but also a similar IgG1 and IgG2a subclasses pattern with enhanced Th1 immune response toward a balanced Th1/Th2. This final conclusion was further verified by splenocyte-secreted cytokine analyses that indicated significantly higher Interferon-γ levels in NSP4-VP6+M50 immunized mice compared to other groups despite unchanged IL-4 amounts.
Although RV challenge experiments were not conducted in the present study, we found that immunization with the NSP4-VP6 fusion protein intraperitoneally induced levels of humoral antibodies against VP6 that may confer protective immunity against RV challenge. Due to enterotoxic activities of NSP4, Abs against this protein may also play a significant role in prevention of diarrhea post-RV infection (30 –32). Although not investigated, it should be noted that the NSP4-VP6 fusion protein presented in this study could also elicit high titers of NSP4 Abs, which might be considered as an immunogen with potential of dual functions (i.e., protection against infection by Abs against VP6 and prevention of diarrhea by Abs against NSP4).
Taken together, to our knowledge, for the first time the NSP4-VP6 fusion protein was introduced in this study as an efficient and alternative platform for RV vaccine development. Our results indicated to the adjuvant properties of NSP4 for enhancement of humoral responses against VP6 antigen. Future functional studies are warranted to investigate the mechanisms behind NSP4 adjuvant activity in animal model following RV challenge and capability of the fusion protein to elicit high titers of NSP4 Abs with preventive properties against RV-induced diarrhea.
Footnotes
Acknowledgment
This study was funded and supported by Pasteur Institute of Iran through Grant No. 745.
Author Disclosure Statement
No competing financial interests exist.