
Abstract
Human parainfluenza viruses (family Paramyxoviridae), human metapneumovirus, and respiratory syncytial virus (family Pneumoviridae) infect most infants and children within the first few years of life and are the etiologic agents for many serious acute respiratory illnesses. These virus infections are also associated with long-term diseases that impact quality of life, including asthma. Despite over a half-century of vaccine research, development, and clinical trials, no vaccine has been licensed to date for the paramyxoviruses or pneumoviruses for the youngest infants. In this study, we describe the recent reclassification of paramyxoviruses and pneumoviruses into distinct families by the International Committee on the Taxonomy of Viruses. We also discuss some past unsuccessful vaccine trials and some currently preferred vaccine strategies. Finally, we discuss hurdles that must be overcome to support successful respiratory virus vaccine development for the youngest children.
Reclassification and Comparison of Paramyxoviridae and Pneumoviridae
T
The most current taxonomy listings and virus names can be found online at
Renamed virus species that use non-Latinized binomial names similar to nomenclature already implemented for six of the eight families of Mononegavirales are listed first (ICTV code 2016.011aM). Names of exemplar virus members and abbreviations are listed in parentheses.
The paramyxoviruses and pneumoviruses have a variety of similarities and differences. The polymerase (L) proteins of viruses from these two families are well separated phylogenetically, having an almost equal phylogenetic relationship with members of the Filoviridae family (e.g., Ebola virus) as with each other (Fig. 1A). On the other hand, the fusion (F) surface glycoproteins of pneumoviruses are much more closely related phylogenetically to those from paramyxoviruses than they are to the glycoproteins (GP) of filoviruses (Fig. 1B). Accordingly, the structures of the F proteins of the paramyxovirus PIV5 and the pneumovirus human respiratory syncytial virus (HRSV or RSV) in their prefusion forms are similar to each other and quite distinct from Ebola virus GP (48,54,96). In addition to having similar domain structures (Fig. 2), the PIV5 and HRSV F proteins have similar intermediate structures that can be inhibited by heptad repeat-derived peptides (46,69,71) and form similar six-helix bundle structures in their postfusion forms (6,56,84). As the F surface glycoprotein is a primary antigen of the pneumoviruses and paramyxoviruses, similar vaccine strategies for both virus families may prove successful.
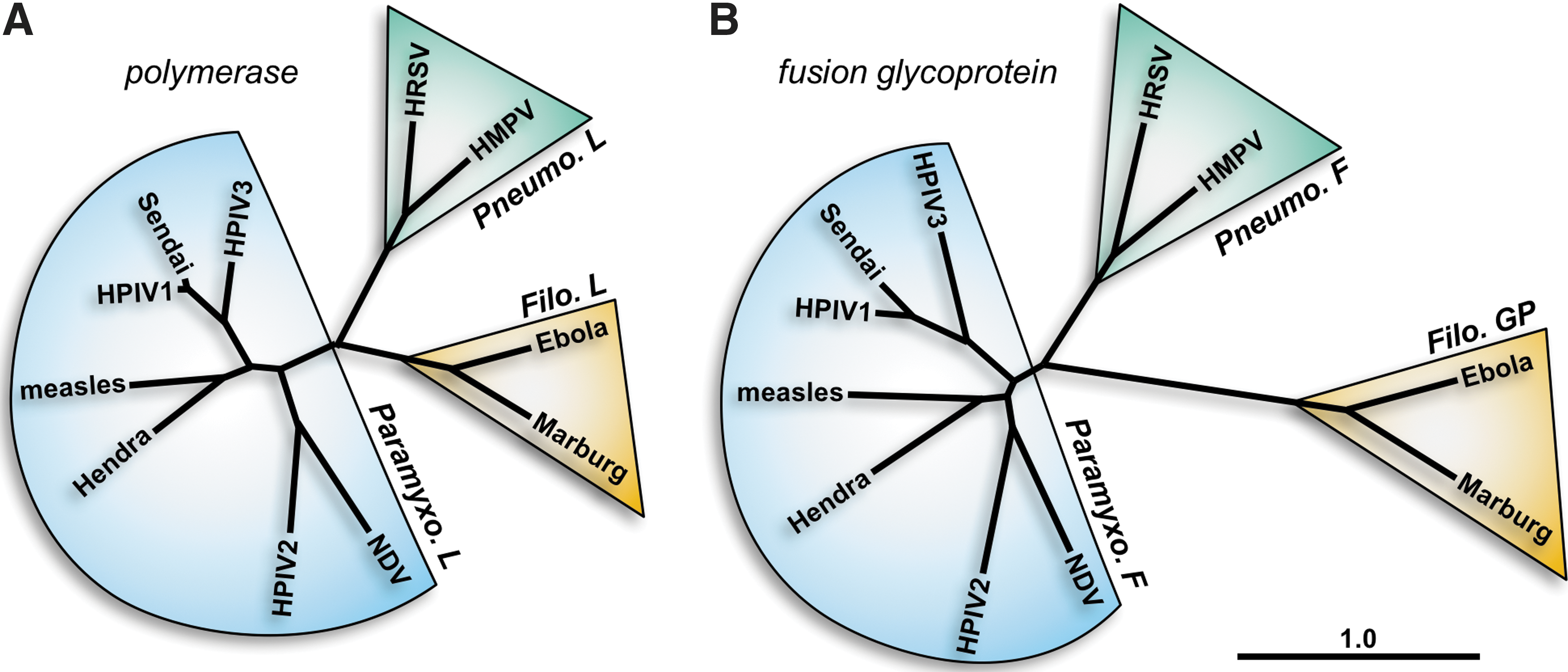
Phylogenetic relationships between representative members of the Paramyxoviridae, Pneumoviridae, and Filoviridae families. Amino acid sequences of the polymerase
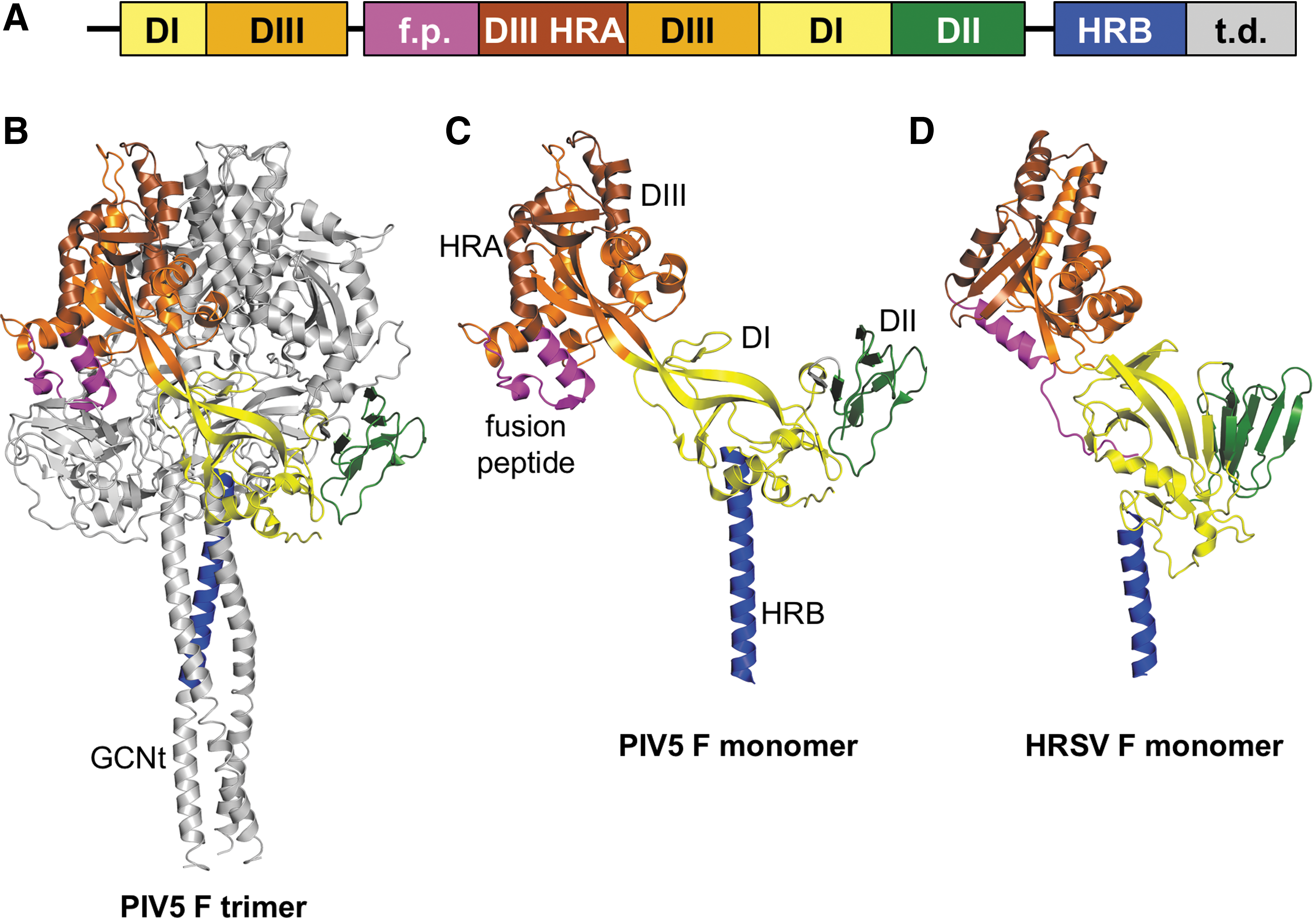
Prefusion structures of the fusion (F) proteins of PIV5 and HRSV.
Virion Structure
As members of the order Mononegavirales, the paramyxoviruses and pneumoviruses have nonsegmented, linear, single-stranded negative-sense RNA genomes (Fig. 3). Their viral RNA (vRNA) genomes are encapsidated by nucleoproteins (N). The genetic cores of these viruses are RNP complexes that consist of vRNA, N protein, phosphoprotein (P), and the large (L) polymerase protein (19). The matrix (M) protein has two compact beta-rich domains connected by a linker (59). The M protein facilitates virus assembly through interactions with the RNP, plasma membrane, and envelope glycoproteins (75,87). While the F glycoproteins of the paramyxoviruses and pneumoviruses are phylogenetically and structurally similar (Fig. 2), the hemagglutinin–neuraminidase (HN), hemagglutinin (H), and glycoprotein (G, for HMPV and HRSV) surface attachment proteins differ in sequence, structure, and binding properties (13,31,47,55,97). For both virus families, the F protein and the attachment protein are exposed on the surfaces of virions and infected cells, help promote virus entry, and provide targets for vaccine development.
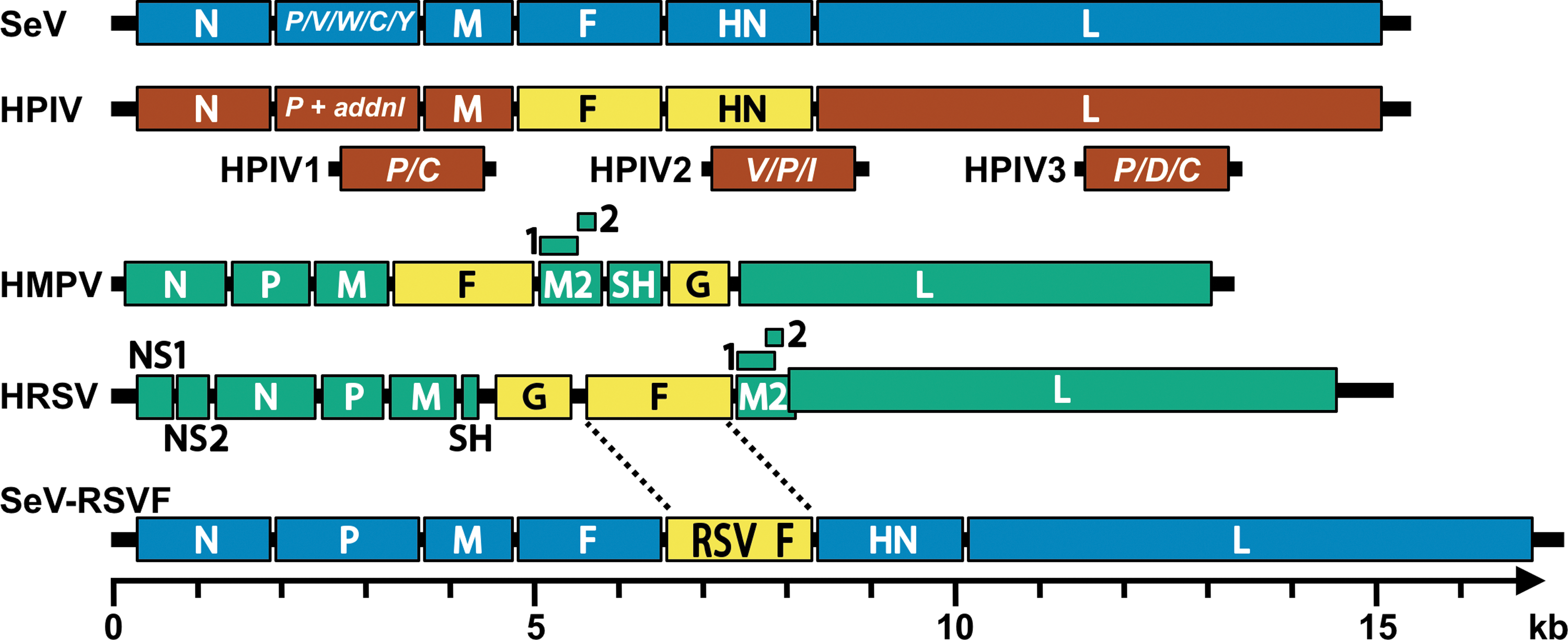
Schematic diagrams of paramyxovirus and pneumovirus genomes. Genomes shown include Sendai virus (SeV, blue), a generic human parainfluenza virus genome (HPIV, brown), human metapneumovirus (HMPV, green), human respiratory syncytical virus (HRSV green) and an RSV F vaccine vectored by Sendai virus (SeV-RSVF, blue with yellow insert). The additional (termed “addnl” in the figure) proteins expressed from the P genes of HPIV1, HPIV2, and HPIV3 are shown in brown below the generic HPIV genome. Surface glycoproteins of the human viruses are shown in yellow. In each genome, the 3′ leader is on the left terminus and the 5′ trailer is on the right. The intergenic junctions are not shown but contain transcription stop, intergenic, and transcription start sequences. In the HMPV and HRSV genomes, the M2 gene contains two overlapping products, M2-1 and M2-2. The RSV M2 and L genes overlap. Genomes are drawn to scale (bottom), except for the generic HPIV genome.
Diseases Caused by the Paramyxoviruses and Pneumoviruses in Infants and Young Children
Lower respiratory tract (LRT) infection is the leading cause of death in low-income nations and the third-leading cause of mortality worldwide (
The Measles Virus Vaccine, a Successful Vaccine for Older Children
Impressive clinical successes in the paramyxovirus field include development of the human mumps and measles virus vaccines. These licensed vaccines are usually formulated as elements of the mumps, measles, and rubella (MMR) pediatric vaccine product, widely distributed throughout the world (44). When communities of individuals reject MMR vaccinations, new measles epidemics and associated morbidities can arise, demonstrating the significant positive influence of the MMR vaccine on human health (25). The MMR vaccine is usually delivered intramuscularly with priming and booster doses.
Measles virus vaccines have changed over the years. One of the first measles virus vaccines was a formalin-inactivated (FI) product. A noted risk with this product was that when children were naturally exposed to measles virus after vaccination, they could suffer enhanced immunopathology (33). Today, live attenuated vaccines are used. Vaccine success is high (>95%) and clinicians hope that the vaccine will eventually eliminate measles virus from the human population. It is noteworthy that the MMR vaccine is usually recommended only for older infants or children over the age of 1 year. These children no longer harbor protective maternal antibodies at high titer, and are therefore particularly vulnerable to measles virus infections (44).
The mumps component of the MMR vaccine is a live attenuated virus developed by the serial passage of wild-type virus. Following vaccination, durable neutralizing antibodies are induced and protection from infection is virtually complete (10).
Past and Current Strategies for Pneumovirus and Paramyxovirus Vaccine Development
The past and current strategies for pneumovirus and paramyxovirus vaccine development for the youngest infants are many, and will be described only briefly in this report. Despite convincing studies using research models, and numerous phase I clinical studies, few of these vaccines have advanced to phase III clinical trials (9,11).
RSV vaccines have been heavily studied, because RSV infections cause high-frequency morbidity and mortality in infants. One strategy that has yielded many candidates for phase I testing has been the attenuation of wild-type human RSV (7,15,23,40,42,51,67,72,83,94). This was originally accomplished by selecting cold-adapted mutants (e.g., RSV cps2, NCT01852266, and NCT01968083-2013) or by deletion of whole genes (e.g., MEDI ΔM2-2, NCT01459198-2011 and RSV LID ΔM2-2, NCT02040831; NCT02237209-2014) or by combinations of both strategies (e.g., RSV LID cp ΔM2-2, NCT02948127-2016). Mutations were also introduced deliberately into genes for internal and/or external proteins (e.g., RSV LID ΔM2-2 1030s, NCT02952339; NCT02794870-2016, RSV ΔNS2/Δ1313/I1314L, NCT03227029; NCT01893554-2013, RSV D46/NS2/N/ΔM2-2-HindIII, NCT03099291; NCT03102034-2017, and RSV 276, NCT03227029). When candidate vaccines were advanced for clinical testing, they were often abandoned or tagged for further mutation if (1) the vaccine was overattenuated and did not induce a robust immune response, or (2) the vaccine was underattenuated and associated with adverse events such as wheezing or reversions to a less-attenuated phenotype (38,52). The production of sufficient vaccine virus for distribution has been another key challenge with this approach.
Another popular strategy has been to produce recombinant vaccines by expressing RSV proteins in a non-RSV viral construct. Examples of live viral delivery vehicles have included a bovine PIV-type 3/HPIV3 chimera (b/hPIV-3) (49,74) and Sendai virus (SeV) (1,32,34,36,68,81,85,86,98 –100). For example, MedImmune advanced a b/hPIV-3 construct carrying a recombinant RSV F gene. Immune responses toward this vaccine were unfortunately lower than expected and there was evidence of RSV F gene instability (88,95). New strategies using the b/hPIV-3 construct are now being developed (50). A chimpanzee-derived adenovirus vector, ChAd155-RSV, has also been tested in adults (NCT 2491463-2015) and is currently being investigated in a phase II study in RSV seropositive infants aged 12–23 months (NCT02927873-2016). The SeV platform has been advanced at St. Jude Children's Research Hospital (1,32). For example, a recom-binant SeV-expressing RSV F has been proven effective in small animals and nonhuman primates, and is expected to advance to a phase I clinical trial soon (34 –36,68,86,98 –100).
Recombinant constructs may be used as vaccines directly, or may be used to produce purified RSV protein vaccines. As an example, Novavax is testing a baculovirus-derived RSV F protein vaccine using recombinant nanoparticle technology. This vaccine was immunogenic in both alum adjuvanted and unadjuvanted preparations in elderly subjects, but the unadjuvanted preparation failed to demonstrate disease prevention in a pivotal phase III trial in the elderly (as described by Fries, L. at RSV 2016, 29 September 2016, Bariloche, Argentina). Currently, a second phase II trial is being conducted in 300 elderly subjects in Australia comparing the immunogenicity of vaccine with no adjuvant, alum adjuvant, and a novel adjuvant called Martrix M-1 (NCT 030263348). Immunogenicity and safety of the alum adjuvanted preparation in women of childbearing age (4,26) and in pregnant women were demonstrated (please see
Other developers (e.g., Vaccine Research Center) are testing prefusion F proteins derived from mammalian cells (28,37,45,50). A study to evaluate the safety, reactogenicity, and immunogenicity of the GlaxoSmithKline RSV investigational vaccine (GSK3003891a) in healthy pregnant women and infants born to vaccinated mothers, using a novel prefusion form of RSV F protein, was cancelled due to instability of the PreF during manufacturing (NCT03191383). Debates continue as to which protein vaccine is best, and which form of protein should be used (e.g., prefusion or postfusion F) (4,45,53 –56,84,101). Today, dozens of RSV vaccine candidates are under development (9).
Recombinant vectors have also been used to target paramyxoviruses or pneumoviruses other than RSV. For example, Mok et al. (58) have used Venezuelan equine encephalitis virus replicon particles, and Russell et al. (70) have used SeV recombinants to present the F protein of HMPV. SeV additionally serves as a Jennerian vaccine against HPIV1, because the two viruses are closely similar, both by protein sequences and by protein conformations (16,66,82). Similarly, BPIV3 or BPIV3 chimeras have been tested as Jennerian vaccines for HPIV3 (5,29,39,41,73). Newer strategies include RNA-based vaccines, such as the HMPV/PIV3 vaccine produced by Moderna, mRNA-1653 (17). Finally, we note that cocktail vaccines have been studied, so that several pathogens can be represented in a single vaccine formulation (e.g., a mixture of three SeV recombinants protected against RSV, HPIV1, HPIV2, and HPIV3 in a cotton rat model) (34,100).
Hurdles That Must Be Overcome
It is perhaps surprising that the vast research dedicated to the paramyxovirus and pneumovirus vaccine fields has not generated licensed products for the youngest infants. One hurdle to be considered is that maternal antibodies may weaken vaccines in young infants, but experiences in other fields and with vaccine models (36) show that vaccines can be efficacious shortly after birth. We consider two additional hurdles as follows.
Lack of community awareness
When a young infant suffers from a respiratory virus infection, the disease is often inaccurately termed “flu.” The reality is that acute lower respiratory viral infections in the youngest children are most often caused by paramyxoviruses or pneumoviruses (61,77). Furthermore, infections with the paramyxoviruses and pneumoviruses are not routinely reported to the Centers for Disease Control and Prevention or other regulatory agencies, meaning that communities cannot easily tabulate the number of infections per year or the disease outcomes [agencies contemplate making RSV infections notifiable in select states on a trial basis to assist future evaluation of candidate vaccines (43)]. The vast morbidity and mortality caused by infections with the paramyxoviruses and pneumoviruses perhaps go unrecognized by the public eye, and the value of potential protective vaccines is not understood. Instead, the risk of vaccine development may be perceived as too high, and research funds may be diverted elsewhere.
Vaccine-induced inflammation in the respiratory tract: beneficial or pathogenic?
The paramyxoviruses and pneumoviruses are somewhat unique in that they often strike the respiratory tracts of the youngest infants. If a vaccine can induce antibodies and effectors (e.g., plasma cells and T cells) that target virus in the respiratory tract, this may have the desired effect of inhibiting virus replication in the lung. However, by definition, entry of cells into tissues is termed “inflammation,” a condition that is generally feared in the context of the pediatric respiratory tract. The situation differs from vaccination with other products such as the polio vaccine, after which cellular responses in the mucosa are expected and tolerated (64).
A fear of inflammation in the pediatric respiratory tract is warranted, because excessive cell influx into the airways can block respiration. An experience of the 1960s provides an example. When an FI-RSV vaccine was tested in a clinical trial, vaccinated children fared worse than their control counterparts following natural exposure to RSV (reminiscent of the immunopathology associated with the FI-measles vaccine previously described). There were numerous hospitalizations and two deaths caused by the vaccine (12). The explanation for the FI-RSV vaccine outcome remains unknown. Perhaps, because there were no neutralizing antibodies induced by the vaccine, virus trafficked to the LRT unperturbed; innate and adaptive immune cells then infiltrated the LRT and constricted the airways (60).
To avoid consequences similar to those observed with FI-vaccines, new respiratory virus vaccines must induce balanced inflammatory responses. A robust, acute, local immune response in respiratory tissues may be desired to support rapid virus clearance and to avoid tissue damage [and consequent, enhanced inflammatory responses (65)], but the initial cell recruitment into respiratory tissues must not be so great as to constrict the airways. How robust should the response be? This question is difficult to answer, in part because responding cells are heterogeneous in nature, and in part because a cell population that might benefit one individual might harm another. Outcomes will depend on the age, size, and lung constitution/environment of each vaccinee.
When particular cytokines, CD4+ T cells, or eosinophils (to name a few factors) are upregulated locally or systemically by vaccination with a new candidate respiratory virus vaccine, concerns are raised in the research community that can discourage advanced vaccine development. Debates then continue as to the benefits/risks of particular immune effector populations, and models/assays by which effectors should be measured (8,24,63,76,89,91 –93). Currently, such debates are unresolved, but their eventual resolution may greatly accelerate the development of paramyxovirus and pneumovirus vaccines for the pediatric arena.
Conclusions
Mumps and measles virus vaccines mark two successes in the paramyxovirus field, but these are often recommended for children age 1 or older. Neither the paramyxovirus nor pneumovirus field has yet to produce a licensed vaccine for the youngest infants. While numerous promising vaccine candidates exist, major hurdles are also present. The public is often unaware of the diseases caused by paramyxoviruses and pneumoviruses, and therefore does not appreciate the importance of associated vaccines. Also, researchers must be assured that vaccine-induced inflammation in the respiratory tract will be effective without constricting the airways. These significant hurdles must be overcome if vaccines for the paramyxoviruses and pneumoviruses are to be developed for the youngest children.
Footnotes
Acknowledgments
Work was supported by NIH NCI P30 CA21765 and ALSAC.
Author Disclosure Statement
Julia L. Hurwitz and Charles J. Russell are authors of a patent describing a Sendai virus-based vaccine vector. No competing financial interests exist.