
Abstract
Measles remains an important cause of child morbidity and mortality worldwide despite the availability of a safe and efficacious vaccine. The current measles virus (MeV) vaccine was developed empirically by attenuation of wild-type (WT) MeV by in vitro passage in human and chicken cells and licensed in 1963. Additional passages led to further attenuation and the successful vaccine strains in widespread use today. Attenuation is associated with decreased replication in lymphoid tissue, but the molecular basis for this restriction has not been identified. The immune response is age dependent, inhibited by maternal antibody (Ab) and involves induction of both Ab and T cell responses that resemble the responses to WT MeV infection, but are lower in magnitude. Protective immunity is correlated with levels of neutralizing Ab, but the actual immunologic determinants of protection are not known. Because measles is highly transmissible, control requires high levels of population immunity. Delivery of the two doses of vaccine needed to achieve >90% immunity is accomplished by routine immunization of infants at 9–15 months of age followed by a second dose delivered before school entry or by periodic mass vaccination campaigns. Because delivery by injection creates hurdles to sustained high coverage, there are efforts to deliver MeV vaccine by inhalation. In addition, the safety record for the vaccine combined with advances in reverse genetics for negative strand viruses has expanded proposed uses for recombinant versions of measles vaccine as vectors for immunization against other infections and as oncolytic agents for a variety of tumors.
Introduction
M
MeV, the causative agent of measles, is a negative-sense RNA virus with a nonsegmented genome and a lipid envelope that belongs to the morbillivirus genus of the family Paramyxoviridae. The 16 kb genome encodes eight proteins. Six proteins are found in the virion. The envelope has surface projections composed of the viral hemagglutinin (H) and fusion (F) glycoproteins. H interacts with the virus receptor on susceptible cells for attachment and F interacts with H and with the cell membrane for fusion and entry. F is produced as a precursor F0 that must be processed by cellular proteases to the disulfide-linked F1–F2 fusion-active form for virions to be infectious. The helical ribonucleocapsid is formed from the genomic RNA wrapped by the nucleocapsid (N) protein with the phosphoprotein (P) and large polymerase (L) proteins attached. The matrix (M) protein interacts with the ribonucleocapsid and glycoprotein tails for virion assembly. C and V are nonstructural proteins encoded within the P gene that regulate the cell response to infection and modulate IFN signaling (10,21). The V protein interferes with activation of RNA helicase MDA5 by preventing PP1-mediated dephosphorylation of the CARD domain and IκB kinase α to interfere with IFN induction (27,126,132) and with Stat1/2 to inhibit IFN signaling (19,140). The C protein downregulates viral RNA synthesis and production of defective interfering (DI) RNAs to decrease virus detection intracellularly (59,106,121,133). MeV is an antigenically monotypic virus, with 24 genotypes recognized based on the sequence of the C terminus of the N gene (148).
MeV targets several types of cells (e.g., B and T lymphocytes, monocytes, and endothelial and epithelial cells) and uses multiple receptors in a virus strain and cell type-specific manner determined by the H protein. Three receptors have been identified: membrane cofactor protein or CD46, a complement regulatory protein present on all nucleated cells (35,109); signaling lymphocytic activation molecule (SLAM) or CD150, present on activated immune cells (177); and polio virus receptor related 4 or nectin 4, present on epithelial cells (101,114,158). The binding sites for these cellular receptors are overlapping on the lateral surface of the β-propeller head structure of H at sites targeted by neutralizing antibody (Ab) (57,154). MeV probably uses additional receptors for infection of endothelial cells (40) and cells of the central nervous system (91,162). H interactions with cellular receptors promote virus entry by activating F1–F2 trimers and triggering pH-independent fusion of the virus envelope with the cell plasma membrane (134 –136), but other modes of entry may be important in primary cells (46).
Vaccine Development History
Enders and Peebles isolated MeV in tissue culture by inoculating primary human kidney cells with the blood of David Edmonston, a child with measles (38). Two vaccines were developed and licensed: a formalin-inactivated whole virus vaccine and a live attenuated vaccine.
Inactivated vaccine
The formalin-inactivated measles vaccine was delivered with a three-dose schedule, but provided only short-term protection from measles and primed for a more severe disease, atypical measles. Previously vaccinated children exposed to and infected with WT MeV were at risk for developing a prolonged high fever, hemorrhagic or vesicular rash, and severe pneumonitis (142). Studies in monkeys have shown that atypical measles is due to failure of the inactivated vaccine to induce a mature Ab or T cell response with production of low avidity Ab that does not effectively neutralize WT virus. After WT MeV infection, large amounts of low avidity Abs are produced that bind, but do not neutralize, MeV, leading to immune complex formation, vasculitis, and pneumonitis (137,138). Experience with the inactivated MeV vaccine emphasized the potential for vaccine-induced enhancement of disease and was withdrawn.
Live attenuated vaccine
The live attenuated measles vaccine was developed at the same time as the formalin-inactivated vaccine with initial passage of the Edmonston virus in primary renal and amnion cells (Fig. 1). Subsequent passage in chick embryo fibroblasts (CEFs) produced the Edmonston A and B viruses that no longer produced disease in macaques (65). The Edmonston B vaccine was licensed in 1963, but frequently produced fever and rash (64,73). Further virus passage in CEFs yielded the more attenuated viruses Moraten (Attenuvax) and Schwarz that induce fever and rash in <10% of those vaccinated (147,157). Edmonston-Zagreb (EZ) has a slightly different lineage that includes passage in human WI-38 cells (8,128) and is the vaccine strain most widely used in resource-poor countries. Vaccine virus strains, including independently developed CAM-70, Leningrad-16, and Shanghai-191, are all closely related genotype A viruses with few sequence differences (8,127,147).
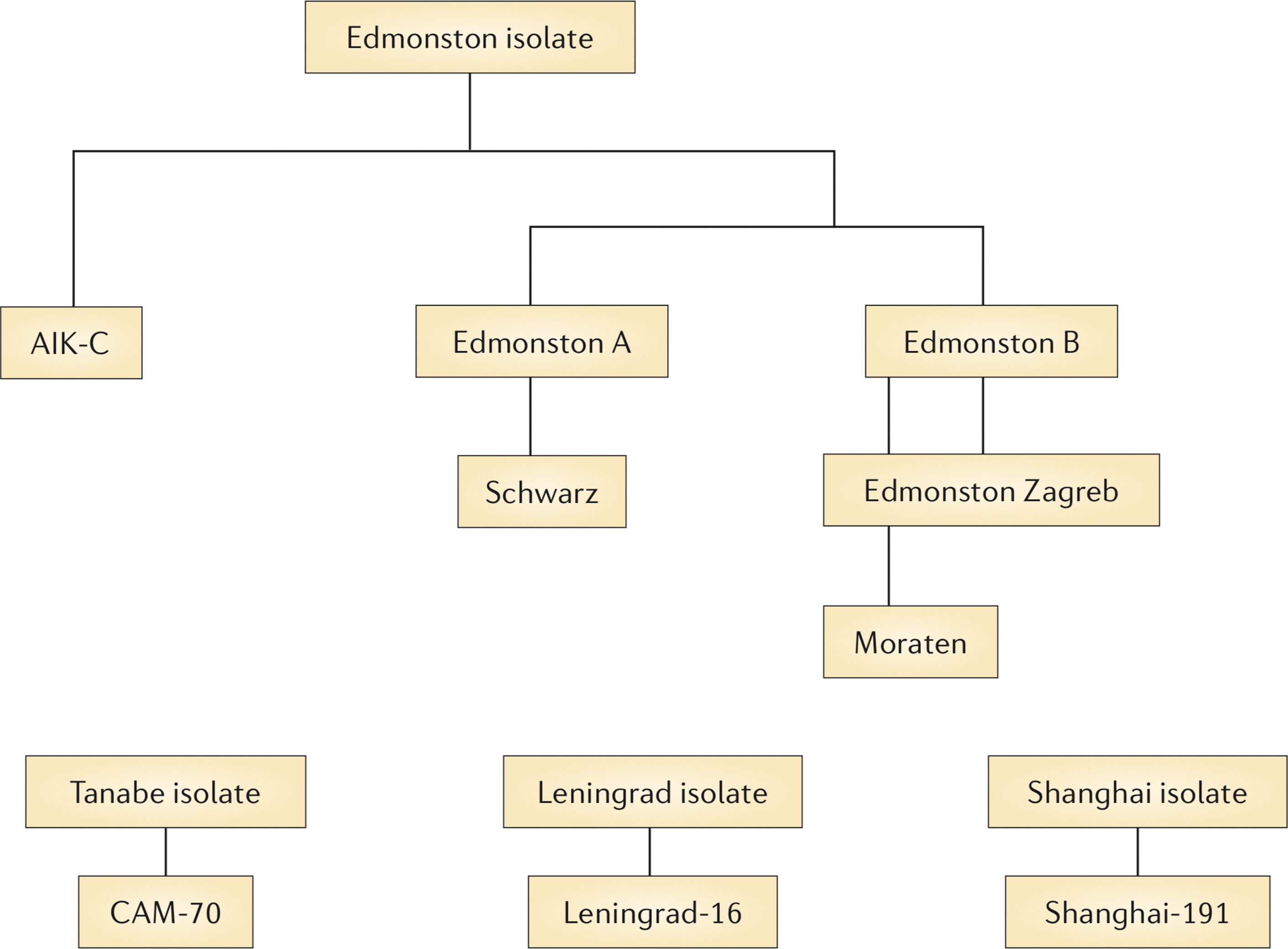
Development of the live attenuated measles virus vaccine. The original Edmonston wild type virus was isolated in human cells and attenuated primarily by passage in chick embryo fibroblasts. Edmonston B was the first licensed vaccine, but fever and rash were frequent complications. The further attenuated Schwarz, Moraten, and Edmonston-Zagreb vaccine strains are widely used throughout the world. From Moss and Griffin (96) with permission of the publisher.
Molecular Determinants of Attenuation
The original Edmonston strain of MeV is not available and genotype A viruses are extinct (148), so it is not possible to directly compare attenuated vaccine viruses with the original WT virus from which they were derived. Although the earliest available passaged virus (Edmonston “WT”) does not cause a rash in macaques, it does produce a viremia (5,181). During adaptation of WT Edmonston to growth in CEFs (67), the virus was attenuated, but continued to replicate well enough to induce an immune response that protected monkeys from experimental infection and children from acquiring measles (37,66). Moraten, Schwarz, and EZ vaccine strains are in widespread use today. Moraten and Schwarz are identical in sequence and EZ differs from Moraten/Schwarz at 21 amino acids (8,127,128). Sequences of vaccine strains compared with current WT strains reveal differences in most viral proteins, any of which may contribute to attenuation and no one change or combination of changes has been identified as responsible for attenuation (6,8,36,63,105,127,128,173 –175).
Because adaptation of WT MeV to growth in Vero cells selects for a virus that no longer causes a rash in monkeys (6,70,175), many studies of virulence have focused on the changes required for growth of WT MeV in Vero cells or CEFs. Changes in multiple protein genes (P/C/V, M, F, H, and L) are selected (7,36,60,94,160,168,169,172,173,178,186). Construction of recombinant viruses using the Japanese IC-B WT and CAM-70 vaccine strains has shown that changes in M, F, H, and L all contribute to efficient growth of CAM-70 in CEFs (63,160), but which one or combination of changes alters in vivo virulence has not been identified.
Potential importance of H
One potentially important biologic difference is the acquisition of efficient use of the CD46 receptor by vaccine strains (17,39,187). Tyr at position 481 of H (present in all vaccine strains) and Gly at 546 (present in Moraten, but not in EZ) are key determinants of the affinity of H for CD46 (9,161), but other residues also contribute to this interaction (88,147, 155,168). The mechanism by which gaining use of the CD46 receptor might lead to vaccine attenuation is not clear, unless the important consequence is loss of another interaction such as H binding to toll-like receptor (TLR) 2 (12).
SLAM is expressed on immature thymocytes, activated lymphocytes, activated monocytes, and mature dendritic cells (DCs) (20,32) and is the most important receptor for MeV infection of lymphoid tissue (28,75). H residues important for binding SLAM are generally shared between MeV strains, and both vaccine and WT viruses use SLAM as a receptor (39,86 –88,111,119,153,155,182,187). Evaluation in cynomolgus macaques of recombinant enhanced green fluorescent protein (eGFP)-expressing WT viruses with vaccine (Ed-tag) H instead of WT (IC-B) H showed attenuation without a change in tropism, suggesting that the important effect is on replication rather than on receptor binding (176). Viruses with WT Asn at H481 interact with SLAM, but not CD46, activate TLR2, and enter peripheral blood mononuclear cells (PBMCs) more efficiently than viruses with Tyr (confers CD46 binding) at this position (12,39,155), but the importance of changes in any of these parameters for in vivo attenuation is unclear.
Potential importance of C and V
Differences in induction of IFN have been proposed to explain the differences between WT (good blocking of IFN induction) and vaccine (poor blocking of IFN induction) strains of MeV in ability to cause disease (118). Multiple in vitro studies have compared type I IFN induction by vaccine and WT strains (110,132,163) of MeV. Some studies have shown more efficient induction of IFN by vaccine MeV, whereas others have not. However, interpretation has been complicated by use of vaccine virus stocks that contain viral particles with DI RNAs that efficiently induce IFN and are produced during MeV replication in tissue culture (68,163,165). The C and V sequences of vaccine and WT MeV strains are similar (44), but the literature on sequence-dependent effects on function has been complicated by the analysis of recombinant viruses that contain mutations (Y110H, C272R) present in the early MeV vaccine cDNA clone (EdTag) used for reverse genetics that are not present in vaccine strains (33,159,171). Analysis of validated C and V proteins from vaccine and WT strains shows no differences in ability to regulate the IFN response (44,105).
There is little evidence of type I IFN induction in humans with measles (190), but this has been difficult to study because infection is not recognized until after the adaptive immune response (rash) appears ∼2 weeks after infection. Therefore, tissues from macaques infected with vaccine and WT strains of MeV were analyzed from the time of infection using quantitative reverse transcriptase (RT)-polymerase chain reaction to assess levels of IFN and IFN-stimulated gene (ISG) mRNAs and a flow cytometry-based bioassay to assess levels of biologically active IFN both early and late after infection. These studies showed little to no induction of type I IFN, type III IFN, Mx, or ISG56 mRNAs in monkeys infected with vaccine or WT MeV and no IFN was detected by bioassay (164). Therefore, there is no evidence that the MeV P/C/V protein gene is a determinant of vaccine attenuation.
Biologic Determinants of Attenuation
The in vivo target cells for vaccine strains of MeV are not well characterized, but replication is restricted compared with WT MeV despite an enhanced ability to use widely distributed CD46, as well as SLAM, as a receptor. Limited in vivo studies suggest that vaccine and WT viruses replicate equally well in the respiratory tract, but that vaccine virus replicates less well in lymphoid tissue resulting in lower levels of virus in circulating PBMCs (viremia), potentially accounting both for less serious disease and a less vigorous immune response to infection (5,78,181). DCs and tissue macrophages are major early myeloid cell targets of WT and vaccine MeV (28,30), and after intramuscular infection of macaques with eGFP-expressing EZ MeV vaccine, infected DCs and macrophages can be visualized at the site of inoculation (144).
During systemic spread of WT MeV, vascular endothelial cells and epithelial cells in multiple organs become infected (40,95). Endothelial cells express CD46 (91) but not SLAM (3), can be induced to express nectin 4 (1), and infection may provide an important pathway for entry into many tissues. In vitro studies of human microvascular and umbilical vein endothelial cells have shown that vaccine strains infect endothelial cells more efficiently and replicate virus better than WT MeV (1,3). Likewise, monocyte-derived DCs replicate vaccine MeV better than WT MeV, do not produce types I or III IFN in response to infection, and develop very limited upregulation of ISGs (165).
In vitro studies of primary human myeloid and lymphoid cells and thymus and tonsil explants tend to show that MeV strains initiate infection similarly in cells, but that less infectious virus is produced by vaccine than WT strains (4,26,30,55,117,165,180). CD4+CD8+ double-positive thymocytes are susceptible to infection with vaccine and WT MeV, but vaccine strains infect less efficiently (require higher multiplicities of infection) and take longer to produce virus than WT strains (117).
Vaccine-Induced Protective Immunity
The live attenuated MeV vaccine induces both Ab and cellular immune responses that mature over months (78,104). The vaccine protects against infection with all genotypes of MeV (8). Protection correlates best with the quality and quantity of neutralizing Ab induced (23,139) but T cells likely contribute (152). Ab that neutralizes infectivity of MeV is directed primarily against the H protein, with some contribution from Ab to F (29). H also contains important CD8+ T cell epitopes (121). High avidity Ab to H is required for neutralization of SLAM-mediated WT MeV infection of lymphoid cells, but not for neutralization of CD46-mediated vaccine virus infection of Vero cells commonly used for measuring levels of MeV neutralizing Ab (138). Studies in macaques have shown that neutralizing Ab predicts protection from disease (rash), but not necessarily from infection, and that T cells alone do not protect from either infection or disease, but do facilitate clearance of viral RNA (80,122 –125).
Although vaccine-induced immune responses are qualitatively similar to those induced by infection with WT MeV, levels of Abs, particularly H-specific neutralizing Abs, are lower after vaccination (116). Young age affects the quality and quantity of infant Ab responses, but has less of an effect on T cell responses (48,49,103,143). Increasing the dose of vaccine ∼100-fold (from 103 to 105 pfu) improved the Ab responses in young infants, but resulted in an unexpected increase in mortality for girls (50,58,61,69), so this WHO recommendation was withdrawn. With time, MeV-specific Abs and CD4+ T cells induced by vaccine decrease (22,24,71,108), and secondary vaccine failure rates are estimated to be ∼5% 10–15 years after immunization (2,89). Lack of a sustained protective response is a particular problem in HIV-infected children (45,98,104,112). Attempts to boost immunity with repeated immunization have generally led to only transient improvements in either levels of neutralizing Ab or MeV-specific T cell responses (42) and raise the possibility that new vaccine approaches may be required for measles control in the future (54).
Vaccine Delivery and Control of Measles Transmission
In industrialized countries, the current vaccine is usually given subcutaneously or intramuscularly in combination with live virus vaccines for mumps, rubella (MMR), and sometimes varicella (MMRV). In resource-poor countries, MeV vaccine is often given alone, but there is an effort for these countries to transition to delivery of combined measles and rubella (MR) vaccine (72).
Not all children respond to the initial dose of MeV-containing vaccine given in infancy (85% at 9 months, 95% at 12 months) (179), so two doses are required to provide a second opportunity for response and achieve a population immunity of 92–95% required to eliminate endemic transmission (51). With institution of mass campaigns to deliver the second dose in developing countries, strides have been made toward global measles control, but logistical and financial difficulties have slowed progress with recent increases in measles cases and deaths in many countries (97,130,166). In addition, complacency, concerns about safety, and philosophical and religious objections to vaccination have resulted in a resurgence of measles in many industrialized nations (43,102,130,145,151). Delivery by inhalation rather than needle and syringe has been proposed to help improve coverage (56,84,120), and both aerosol and dry powder versions of the EZ vaccine are in development for use both for use in primary immunization of infants and for delivery of the second dose to older children (18,34,52,56,76, 78,84,150).
Expanded Uses of MeV Vaccine
Vaccines against other infectious diseases
Efficient systems have been developed for production of recombinant MeVs (25,131,144) and these techniques are being applied to develop MeV vaccine as a vector for immunization against other virus infections (82,100): for example, dengue (13,15,16), West Nile (31), HIV (83,167), MERS (85), and chikungunya (CHIKV) (14). A phase 1 trial of the MeV recombinant CHIKV vaccine showed that it was safe and induced Abs to CHIKV in adults with pre-existing MeV immunity (141).
Cancer treatment
Because the MeV vaccine is cytolytic for many tumor cells in vitro (11,113), recombinant MeVs are also being retargeted as oncolytic agents for cancer therapy. These approaches have shown promise in animal models and clinical trials using doses orders of magnitude higher than those required for vaccination and have been initiated for treatment of multiple myeloma, ovarian cancer, T cell lymphoma, and glioblastoma (47,74,81,99,149,183,188,189).
These new uses for MeV vaccine are likely to continue to expand and increase the need for a better understanding of the mechanisms of attenuation, sites of replication, and immunogenicity of MeV vaccine.
Summary
The live attenuated MeV vaccine in current use is safe and effective and has saved millions of lives. The vaccine induces Ab and T cell-mediated immunity that is durable, but not readily boosted. Despite long use, neither the determinants of attenuation nor of protective immunity have been identified and deserve investigation. The reasons for failures of the formalin-inactivated vaccine and the high titer live virus vaccine are only partially understood and provide cautionary tales for development of other vaccines. Success of the current vaccine has led to expanded development as a platform for other vaccines and as an oncolytic agent.
Footnotes
Acknowledgments
Work from the author's laboratory was supported by research grants from the National Institutes of Health (R01 AI023047, R21 AI095981, and T32 AI007417) and by the Bill and Melinda Gates Foundation. The contributions of many members of the laboratory who have helped with these studies are gratefully acknowledged.
Author Disclosure Statement
No competing financial interests exist.