
Abstract
To assess if the difference in species-specific immune response to RV-C correlates with a higher frequency of reinfection, shorter time to reinfection, or different symptom severity than infections with RV-A or RV-B. Forty-three patients were enrolled of which 34 were successfully tracked longitudinally over 3 months, with nasal swabs and symptom questionnaires provided every 2 weeks to identify rhinovirus (RV) strains and the concurrent symptomatology. No difference was found in the time to reinfection with an RV species between RV-C and RV-A or RV-B (p = 0.866). There was a trend toward more rapid reinfection with the same species in RV-C than RV-A (55.1 days vs. 67.9 days), but this failed to reach statistical significance (p = 0.105). RV infections were generally associated with only minor symptoms, with rhinorrhea being the only significantly associated symptom (p = 0.01). RV-C was shown to have higher levels of lethargy and wheeze than other RV species. Time to reinfection with subsequent RV is not influenced by the species of the preceding RV.
Introduction
R
There are now three identified species of RV: RV-A, RV-B, and RV-C. Within each of these there are multiple strains (also called “serotypes” or “types”) (28). RV-C was first identified in 2006 (21) from respiratory samples collected in Queensland and New York (3,20), and following further analysis was designated a separate RV species in 2010 (8). All RV species have the potential to cause disease, yet there is increasing evidence that RV-C is more pathogenic than other RVs as it uses a different host receptor to RV-A and RV-B (5), and also RV-C has been associated with increased asthma exacerbation severity (4) and increased likelihood of readmission to hospital, particularly in children with atopy (9).
The innate immune response to RV is complex and involves the interaction between many components of the immune system (6). It has been found that immune response varies between RV species. In healthy adult sera, the species-specific antibody binding to RV-C was lower than that for RV-A and RV-B (13). A follow-up report by the same team showed markedly lower species-specific IgG titers to RV-C and RV-B than to RV-A in asthmatic children and even in patients where RV-C had been concurrently detected (12). This low level of species-specific response to RV-C may imply a less effective immune response to RV-C, and thus, a heightened risk of subjects to repeated reinfection.
Objectives
We aimed to assess if there was a different rate of reinfection or symptom severity between each RV species. Our hypothesis was that the lower level of species-specific immune response to RV-C would correlate with a higher frequency of reinfection, with shorter duration between periods of RV detection in cases where RV-C was the primary infection.
Methods
Study population
Cases were enrolled in this study over the course of 3 months during autumn and winter as part of the Mechanisms of Acute Viral Respiratory Infection in Children (MAVRIC) cohort. These children were younger than 16 years and presented to our tertiary children's hospital emergency department with an acute wheezing illness. Parents and children were asked to participate in a substudy investigating the temporal pattern of RV infection over 12 weeks. Controls who had no evidence of a current respiratory infection were also recruited and consisted of siblings of the cases or healthy children from the community. Children with asthma were not excluded as controls, but could not have had a presentation to hospital with acute wheeze in the previous 6 months. Children were excluded from the study if they were born premature (≤36 weeks) or had a chronic underlying disorder apart from asthma.
Parental consent was obtained for all participants, and the Princess Margaret Hospital Ethics Committee provided approval for this study.
Sample and data collection
At recruitment, a questionnaire was given to the parents of both cases and controls asking about current symptomatology as well as medications used in the past or for the current episode of illness (if relevant). If symptoms of respiratory infection had been apparent before their presentation, parents were asked to recall which symptoms they had and their duration. An extensive questionnaire assessing their history of respiratory illnesses and environmental exposures was completed by the parents at the time of recruitment. During follow-up, every 2 weeks for 12 weeks, parents were asked to record the presence, duration, and severity (mild, moderate, or severe) of any cough, wheeze, dyspnea, fever, fatigue, runny nose, congestion, sneezing, or any “other” symptom. No guidance was provided with regard to assessing severity and was subjectively assigned by the participant and/or their parent.
Nasal specimens were obtained during recruitment and every 2 weeks for 12 weeks. At recruitment, a Copan® flocked swab (Murrieta, CA) and/or a nasal-blow specimen were obtained. Nasal-blow specimens were obtained in older children who were able to blow their nose into a plastic zip-lock bag, which then had 1 mL of sterile saline added before storage at −80°C for later analysis. The flocked swabs were used on younger children and also for all parent-obtained follow-up samples together with universal transport medium. Parents were instructed on how to collect the nasal swab samples and were provided with six collection kits that included 1 mL of universal transport media. Samples were placed at −20°C until collection of the swabs on ice from patients' homes at 6 and 12 weeks after recruitment. Swabs and labels were then verified and stored at −80°C until analysis.
Molecular detection and sequencing of RV
The nasal samples had RNA extracted and complementary DNA prepared. A two-stage PCR and gel electrophoresis were used to detect the presence or absence of RV RNA determined by successful amplification of the HRV 5′ untranslated region and visualization of the PCR product on a 2% agarose gel containing ethidium bromide. Following detection, DNA sequencing was carried out using purified DNA samples and RV phylogenetic analysis was performed. Clustal X (v 2.0.12) software was then used to align the sequenced strain with the sequences of all 153 known RV strains. Separate samples identified as having the same HRV strain were further investigated and Sequencher (v .48) used to determine persistent HRV infections and reduce the likelihood of results having occurred through contamination due to physical proximity during RNA extraction. Assessment was made of differences and similarities in base-pairs, and samples with the same HRV strain but differing base-pairs were considered noncontaminated. The reference strain that aligned to the unknown HRV strain with the highest percentage (>93%) was identified as the strain present in the subject.
Statistical analysis
Due to similarities in demographics and rates of viral recurrence between cases and controls, data were pooled to allow for more robust analysis.
For recurrence analysis, each instance of viral detection was recorded and paired with the time to subsequent RV detection. For instances where the same viral strain was detected in more than one consecutive sample, these were categorized as a single infection. If the same strain was detected in nonconsecutive samples, it was considered a separate episode of infection. Different strains of RV detected in the same child in consecutive samples were classed as separate episodes of infection. Time points where two strains were detected simultaneously or the strain was unclear were removed from recurrence analysis. If a sample was not collected between one RV detection and another in the same child, then this recurrence episode was excluded. Other respiratory viruses were tested for at all time points, however, only three episodes of non-HRV respiratory virus were detected—two of which were in the same child—and so were excluded from analysis. The duration between episodes of viral detection was analyzed using Kaplan–Meir survival curves, and the Mantel–Cox test was performed to determine if time to reinfection differed between RV species. A chi-squared test was used to determine if there were differences in the frequency of species-to-species and species-to-none recurrence in this population.
Differences in symptom frequency were assessed using chi-squared analysis, and differences for mean symptom severity and duration between RV species were analyzed with the Kruskal–Wallis test. Analysis was performed using SPSS version 22.0.0.0.
Results
Subjects
From April to June 2013, 20 cases and 23 controls were recruited for this study. Six cases recruited from MAVRIC and two controls did not return any specimens subsequent to recruitment and were excluded from the analysis. One further case did not return any swabs for three consecutive time periods so was excluded, leaving a total of 13 cases and 21 controls for final analysis. Of these 34 participants, there were 19 females and 15 males with an age range of between 5.5 months to 15 years, a mean of 4.5 years and a standard deviation (SD) of 3.7 years. No patient required representation or admission to hospital during the study period.
At the end of the trial period, 217/238 (91.2%) possible swabs and symptom diaries were collected. There were 21 time periods where swabs were either not collected or not performed and where the symptom diary was incomplete. These periods were excluded from symptom analysis.
Virology and recurrence
Patients recruited as cases had RV detected at recruitment in 12/13 (92%) samples, whereas the rate of detection in controls was 8/21 (38%) at recruitment (p = 0.0001). There was no significant difference in the mean number of RV detections in both cases (37/13) and controls (60/21) over the course of the study, with a mean of 2.8 episodes (SD 1.4) versus 2.8 (SD 1.3), respectively (p = 0.94). Analysis of recurrence data was completed on the pooled results of all children due to this similarity in cases and controls and the high frequency of viral detection.
Of the 217 swabs collected, 97 (44.7%) had detectable RV and of those, RV-C was the most commonly isolated species (n = 48, 49%), followed by RV-A (n = 37, 38%) and RV-B (n = 11, 11%). There was only one episode in which two RV strains were detected from the same sample. Only two participants had no detectable virus throughout the duration of the study. RV was detected in 15 of the last samples collected, so no recurrence data were available for those episodes. In three cases, a single strain of RV-A was detected in two consecutive samples, and in one case, it was detected in four consecutive samples. In two cases, a single strain of RV-B was detected in two consecutive samples. The same strain of RV-C was not found over two consecutive time points.
There was a high rate of reinfection across all RV subtypes. As seen in Table 1, RV recurrence was found 65 times in the 74 positive samples suitable for recurrence analysis, and reinfection with the same species occurred 30 times. There is no evidence that RV-C had a higher rate of reinfection with the same species than RV-A (51.2% vs. 36.4%, respectively, p = 0.257).
RV, rhinovirus.
Time to RV species recurrence was similar for each RV species. As seen in Figure 1, the majority of recurrence occurred within 40 days of the primary infection. The mean time to reinfection for RV-A was 43 days (95% CI 28.9–57.5), for RV-B was 40.6 days (95% CI 21.4–59.8), and for RV-C was 37.3 days (95% CI 29.5–45.1), (p = 0.866).
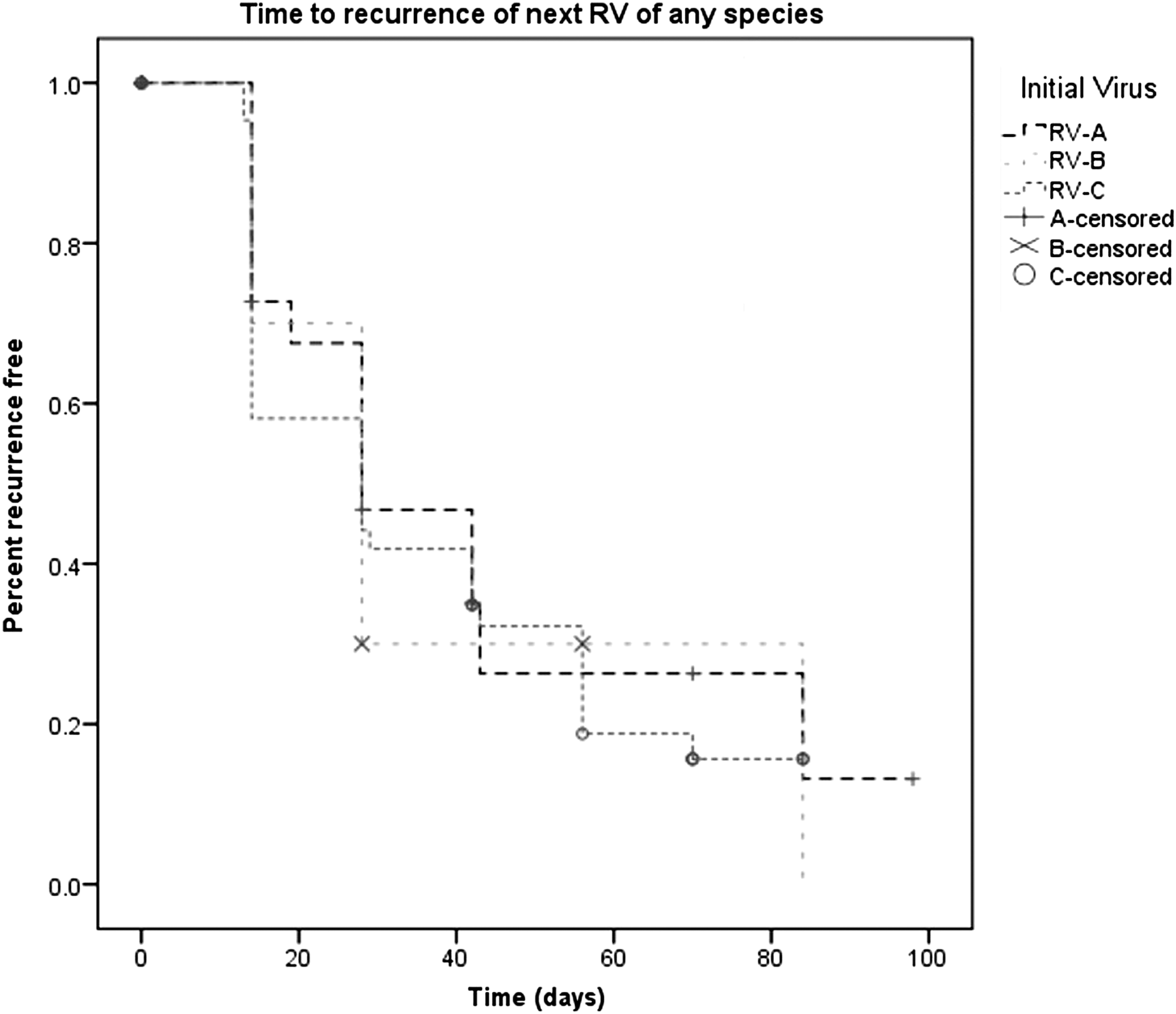
Kaplan–Meier survival curve showing time to reinfection with any rhinovirus (RV) species.
Further analysis (as seen in Fig. 2) showed that when RV-A and RV-C were compared, there was a trend toward a significant difference in the mean time-to-recurrence with the same species (p = 0.105). The mean time from the primary RV-A infection to subsequent RV-A infection was 67.9 days (95% CI 55.9–79.9) and from RV-C to RV-C was 55.1 days (95% CI 45.5–64.8). There were no episodes of reinfection with the same species seen if the initial virus was RV-B.
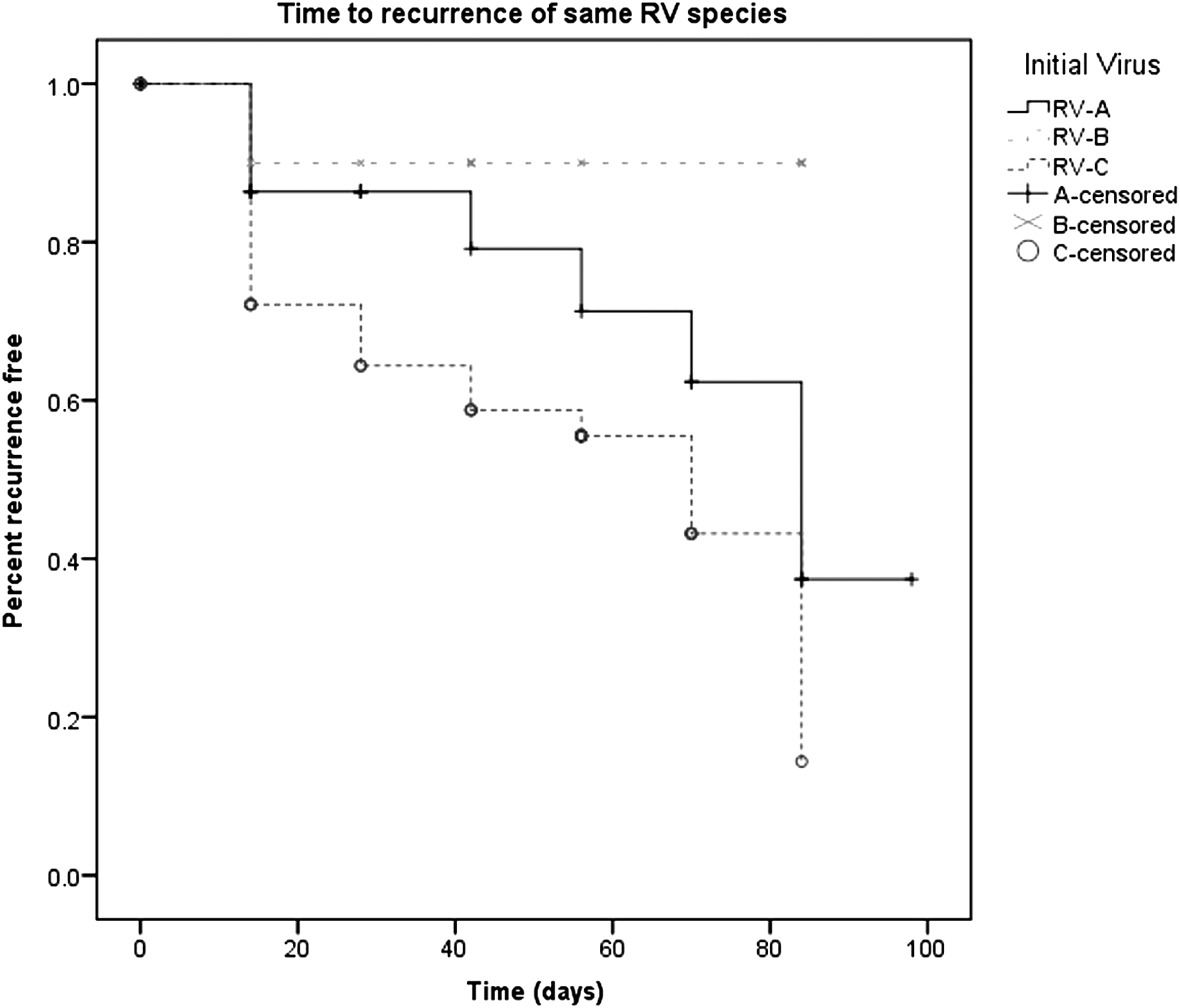
Kaplan–Meier survival curve showing time to reinfection with the same rhinovirus species.
Symptomatology
Symptoms were reported in the majority of diary cards with 137/217 (63%) reporting at least one symptom. Rhinorrhea was the most frequent symptom and was present in 87/217 (40%) of reports. Cough was the second most common symptom (n = 78, 36%) followed by congestion (n = 44, 20%), wheeze (n = 40, 18%), lethargy (n = 35, 16%), sneezing (n = 33, 15%), fever (n = 27, 12.4%), and dyspnea (n = 25, 11.5%).
Of the 80 time points during which children reported no symptoms, RV was detected in 29 of their concurrent nasal samples (36%). RV-C was the most frequently detected virus in asymptomatic children (n = 14/80, 17.5%) followed by RV-A (n = 10/80, 12.5%) and RV-B (n = 5/80, 6.3%). In children where RV-A was detected, symptoms were absent in 10/37 cases (27%) and in those with RV-C detected, 14/48 cases (29%) had no symptoms. RV-B had asymptomatic carriage rates of 45% (5/11), but the frequency of detection was much lower than other species.
As seen in Table 2, rhinorrhea was associated with the presence of an RV infection (50% vs 32.5%, p = 0.01), whereas no other symptom was present at a higher level in those with detectable RV than without. RV-C was more associated with lethargy than RV-A (29.2% vs. 10.2%, p = 0.04), but no other significant interspecies differences were found. There were other indicators that RV-C was associated with higher frequency of symptoms than RV-A with wheeze (31.3% vs. 16.2%, p = 0.11), dyspnea (13.8% vs. 8.1%, p = 0.16), and fever (18.8% vs. 8.1%, p = 0.16), all approaching but not reaching statistical significance.
Discussion
This prospective study of 34 patients yielded 217 time points, each providing symptomatic information and nasal swabs to detect the presence of RV. This study provides unique longitudinal insight into RV duration and recurrence. Over the 3-month period of this study, there was a high rate of RV detection with an average of 2.8 incidents per child and RV detection occurring in 44% of samples. RV-C was the most commonly detected RV, comprising 49% of positive samples. There was a trend toward significance that episodes of RV-C to RV-C recurrence may be more rapid than RV-A to RV-A recurrence. No difference was demonstrated in time to reinfection with any or the same species of RV.
Most episodes of RV infection had resolved by the time the subsequent sample was taken 2 weeks later. The same strain of RV-C was never detected at two consecutive collections, demonstrating that there is an effective immune response that results in viral eradication. This is similar to previous work that showed a shedding time for RV-C of 1 week, shorter than the 2 weeks seen in RV-A and RV-B (30). The majority of infections were associated with reported symptoms, although the presence of symptoms was not a significant predictor to the presence of RV. This highlights that in the majority of instances, RV infections produce only minor symptoms that can also be present even in the absence of RV.
The sample size of the group was the major limitation of the study and unfortunately resulted in several trends being seen that may have reached significance if a larger group had been enrolled. The reliance on parent-reported symptoms may have led to inaccuracies as several criteria such as congestion and dyspnea have few obvious signs. The subjectivity of severity reporting means that there was likely a degree of interparental differences in how the symptoms were ranked. While precise instructions were provided detailing how to collect the samples and previous works have demonstrated the utility of parent-collected samples (31), we cannot guarantee that every sample was collected accurately. Nevertheless, the strengths of the study lie in the high rate of sample acquisition and the successful longitudinal follow-up of the participants over the 3-month time course. Given that 92% of samples were collected, it is unlikely that the missing time points would significantly alter the findings. Also, there were a high number of symptom questionnaire responses that were matched to samples to provide detailed information about RV symptomatology.
The proportion of positive RV-C cases was comparable to that seen in other studies where RV-C comprised between 31.4% and 44.7% (1,2,25,33) of overall RV-positive samples. Overall, RV detection was slightly higher than seen in some other studies where detection rates were between 19.6% and 23% (1,30,34). There is great variability between studies, with Principi et al. finding RV detection frequency in a similar age group of only 23%, whereas the study by Wright et al. showed a frequency as high as 54% in their infant cohort. The high rate of RV detection in this study may be due to recruitment occurring from children who presented to an emergency department with respiratory infection, although the overall frequency of RV detection was no higher in these cases than in healthy controls. One possible reason for this is that four of the controls were siblings of the hospitalized children, and so, there may have been sibling-to-sibling transfer of viruses. Analysis was not performed to detect any evidence of transmission between related participants.
Iwasaki et al. have previously shown that species-specific responses to RV-C were lower than those to RV-A and RV-B (12). However, we found no significant difference in time to reinfection between RV species. Our initial hypothesis was that we would see a higher rate of reinfection in cases where the initial virus detected was RV-C. The trend toward more RV-C to RV-C infections compared with RV-A to RV-A or RV-B to RV-B may represent a less effective immune response to RV-C, or may be due to the generally higher prevalence of RV-C seen during the study. The higher proportion of RV-C in this cohort may be due to the 3-month recruitment occurring over the winter months, when RV-C is known to be more prevalent (23). The lack of reinfection in the RV-B to RV-B group likely reflects the lower prevalence of this species, rather than being due to a difference in immune response although this cannot be excluded.
It has been demonstrated that despite the differences in immune response between RV species, there is still a high level of IgG cross-reactivity (13) that may partially explain the similarity in time to reinfection. It is possible that there may be differences between strain to strain recurrences, with the immune response to RV primarily directed against a small number of frequently mutating structures (18). Analysis of strain to strain recurrences was limited by the sample size and duration of the study. While sample acquisition every 2 weeks is likely to have caught the majority of infections given the shedding time in RV-A and RV-B of 2 weeks, the shorter shedding time of RV-C may have resulted in missing some episodes (30).
Unlike the article by Principi et al. (30), symptoms were present for the majority of time points where RV was detected. That study showed no symptoms despite detection in 64.1% of cases, whereas this study aligns more with data from Byington et al. (7) where only 30% had no symptoms. In all cases, the symptoms were almost exclusively mild. This study showed that rhinorrhea was the only symptom positively associated with RV, which is the same finding as in another study of RV in the aboriginal Australian population (1). It also reinforces that RV-C seems to have some unique characteristics that make it more pathogenic, with higher rates of lethargy and wheezing than seen in RV-A. This would support previous work that showed RV-C is more associated with wheezing episodes requiring admission to hospital and increased wheezing severity (4,9).
Given that there is an established relationship between RV-C and significant wheezing episodes requiring hospitalization (26) and exacerbation of symptoms in children with asthma (32), the reason many RV infections are asymptomatic or minimally symptomatic, whereas others cause significant morbidity, is yet to be systematically ascertained. This study supports the evolving evidence that RV-C produces a more symptomatic illness than other species and further work should be done to explore the ways in which this occurs. Despite the lack of evidence that time to recurrence was different in RV-C, the lower IgG response to this species may have other implications for its pathogenicity beyond recurrence, and this should be investigated further.
Conclusion
No difference was demonstrated in time to reinfection with any or the same species of RV. In the majority of instances, RV infections produce only minor symptoms, which can also be present even in the absence of RV; however, RV-C is more associated with wheezing episodes requiring admission to hospital and increased wheezing severity.
Footnotes
Acknowledgment
We acknowledge the participants who took part in this study.
Author Disclosure Statement
There are no declared conflicts of interest.