
Abstract
The human metapneumovirus (hMPV) is the second leading cause globally of acute infection of the respiratory tract in children, infecting the upper and lower airways. The hMPV may induce an inappropriate Th2-type immune response, which causes severe pulmonary inflammation, leading to the obstruction of airways. Despite its severe epidemiological relevance, no vaccines are currently available for the prevention of hMPV-induced illness. In this investigation, we demonstrated that immunization of mice with the recombinant hMPV nucleoprotein (hMPV-N) mixed with the AbISCO-100 adjuvant reduced viral replication in lungs following challenge with the virus. We found that immunized mice had reduced weight loss, decreased granulocytes in the lung, an increased level of specific nucleoprotein antibodies of IgG1 and IgG2a-isotypes, and a local profile of Th1/Th17-type cytokines. Our results suggest that immunization with the hMPV-N and the AbISCO-100 adjuvant induces a reduction of viral infection and could be considered for the development of an hMPV vaccine.
Introduction
T
Vaccines based on surface proteins have been extensively studied (27). The use of recombinant proteins has induced a neutralizing antibody and protect against lung viral infection and pneumonia in the animal model (10,24). Nonetheless, among all hMPV proteins, the nucleoprotein (protein N) is the most conserved at the amino acid level, with high identity among different genogroups and strains (2). In addition, predictopes from hMPV-N protein induces a strong Th1 profile response and a cytotoxic T-lymphocytes (CTL)-based immunity characterized by secretion of interferon (IFN)-γ (23,28). Moreover, another vaccine against a virus phylogenetically related to hMPV (respiratory syncytial virus) has used nucleoprotein as antigen, showing that the protection is mediated by a Th1 immune response (3,4). However, it is unknown whether the immunization of mice with hMPV-N may prevent hMPV disease development.
Here, we evaluated the property of prototype vaccine based on hMPV-N mixed with AbISCO-100 adjuvant like a potential new target for vaccine development against hMPV. Using the BALB/cJ model of hMPV infection, we immunized, infected, and subsequently evaluated the body weight of animals, infiltration of inflammatory cells and viral RNA into the lungs, Th1/Th2/Th17-type cytokine secretion, and immunoglobulins.
Materials and Methods
Obtaining the recombinant hMPV-N
The hMPV-N gene from a viral strain CZ0107 (clinical isolate obtained from the virology laboratory of the Clinical Hospital of the Pontifical Catholic University of Chile) was cloned into the plasmid pET21a. The plasmid pET21a::hMPV-N was transformed into the Escherichia coli BL21-Codon Plus (DE3) strain. The recombinant nucleoprotein was expressed in Luria Bertani medium, supplemented with 0.4% glucose, IPTG (0.5 mM), at 220 rpm at 37°C for 2 h. Bacterial sediments were recovered by centrifugation at 10,000 g at 4°C for 10 min. Purification of the recombinant nucleoprotein was performed by the Ni-NTA affinity column (Ni-NTA; Invitrogen Purification System), following the manufacturer's instructions. Briefly, the bacterial pellet was resuspended in a binding buffer (20 mM NaH2PO4, pH 8.0, 500 mM NaCl, and 10 mM imidazole), and was treated with lysozyme for 30 min on ice, transferred to a Potter homogenizer, and treated with RNase A (US Biological). The soluble fraction of cell lysis was obtained by centrifugation at 10,000 g for 15 min at 4°C, and the supernatant was filtered through a sterile 0.45 μm filter (Sartorius). The Ni-NTA resin was then rinsed four times with a wash buffer (49.5 mM of NaH2PO4—pH 8.0, 500 mM of NaCl, and 20 mM of imidazole). The recombinant nucleoprotein was eluted using 0.9 mM of NaH2PO4, 500 mM of NaCl, and 25 mM of imidazole, with pH 8.0. Finally, the purified recombinant protein was quantified using the microBCA Protein Assay Kit following the manufacturer's instruction (Pierce).
Detection of nucleoprotein by immunoblotting
The identity of the purified nucleoprotein was analyzed by western blot using a rabbit polyclonal antibody against hMPV (produced in our laboratory). Briefly, purified nucleoprotein was run on sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and transferred to a nitrocellulose membrane for 1 h at 200 mA. The membrane was blocked using phosphate-buffered saline (PBS) 5% bovine serum albumin (BSA) overnight (o/n), and incubated for 1 h at RT with rabbit anti-hMPV antibody (1:1,000). The membrane was washed with PBS-0.05% Tween (PBST) and incubated for 1 h at RT with anti-rabbit IgG antibody (1:1,000) conjugated to alkaline phosphatase (Sigma). The membrane was washed with PBST, and the detection was performed using BCIP/NBT (Calbiochem) as the substrate for alkaline phosphatase. The western blot image was recorded using a Bio-Imaging System (MiniBIS Pro). The whole hMPV protein lysate was used as a positive control for the recombinant nucleoprotein expression.
hMPV preparation
LLC-MK2 (No. CCL-7; American Type Culture Collection, Rockville, MD) cells were used to propagate hMPV serogroup A, clinically isolated CZ0107 (clinically isolated, obtained from the Laboratory of Infectiology and Virology of the clinical hospital of the Pontificia Universidad Católica de Chile). The LLC-MK2 cells were grown in T75 bottles until 70% of confluence in OptiMEM 5% fetal bovine serum (FBS) (Life Technologies Invitrogen, Carlsband, CA), then the cells were inoculated with 2 × 105 plaque-forming unit (PFU) of hMPV in 5 mL of opti-minimum essential media (MEM) supplemented with CaCl2 (100 μg/mL), and incubated for an hour to allow virus adsorption. Then, the medium was changed and incubated at 37°C for 72 h, or until close to 60% cellular detachment was observed. During virus collection, the cells were scraped and the infectious cell culture supernatant was centrifuged twice at 500 g for 5 min to remove the cellular debris. The viral titer of supernatant was quantified by immunocytochemistry using 10-fold serial dilutions over LLC-MK2 cell monolayers 4 × 104 cells per well. The hMPV inoculums were systematically evaluated for potential contamination with LPS or Mycoplasma spp.
Mice immunization and viral challenge
BALB/cJ mice originally obtained from Jackson Laboratories (Bar Harbor, ME) and maintained at the pathogen-free facilities of the Instituto de Salud Pública de Chile (Institute of Public Health in Santiago, Chile) were utilized. All work with animals were done on the basis of institutional guidelines by the institute and supervised by a veterinarian. For immunizations, female mice from 6 to 8 weeks (six per group) received an intraperitoneal injection for each treatment. The immunized group received 20 μg nucleoprotein +14.4 μg of AbISCO-100 adjuvant (hMPV-N/AbISCO group). Control groups received PBS-1X (PBS-1X group), 1 × 106 PFU of ultraviolet light-inactivated hMPV (hMPV-UV group), PBS-1X + 14.4 μg AbISCO-100 (AbISCO group), and 20 μg of nucleoprotein (hMPV-N group). A nonimmunized and uninfected control group (Basal group) was used to provide basal levels of virus infection. At day 14 postimmunization, all groups received an immunization boost. On day 28, mice were challenged intranasally with 1 × 106 PFU hMPV. Finally, mice were euthanized on day 33, following the procedure described below.
Pulmonary cell analyses by flow cytometry
Five days after the infection (33 days postimmunization), the animals were euthanized through an intraperitoneal injection with a mixture of ketamine (110 mg/kg) and Xylazine (5 mg/kg). The lungs were then removed, and the upper, medium, and caval lobes were rinsed with ammonium-chloride-potassium (ACK) lysing buffer. Subsequently, the tissue was cut into small pieces and put in a solution of PBS-1X with collagenase (1 mg/mL) and DNase (50 μg/mL), and the solution was incubated in a shaker (300 rpm) for 15 min. The tissue was then incubated in an ethylenediaminetetraacetic acid and phosphate buffered saline (EDTA-PBS) solution (5 mM) to inactivate the solution with collagenase. The tissue was homogenized using two sterile slides, and finally the cells were filtered with a 40-μm cell strainer (Falcon, BD). Cellular suspensions were spun at 300 g for 5 min and the pellets were resuspended in PBS. Cells were counted in a hematologic chamber. For granulocyte and dendritic cell studies, cells were stained for 30 min at RT with anti-Gr1 (Ly6C/6G)-APC, anti-CD11b-217 FITC, and anti-CD11c-PE (all from BD Pharmingen). Data were acquired on a FACSCanto II flow cytometer (BD Biosciences) and analyzed using FlowJo (TreeStar, Inc.).
Determination of viral load by real-time polymerase chain reaction
Total RNA was isolated from hMPV-infected BALB/cJ lung tissue similarly as described by human respiratory syncytial virus (4,30). Total RNA was isolated from the lungs of the mice using TRIzol (Life Technologies), and 500 ng RNA was reverse transcribed into cDNA using the commercial ImProm-II reverse Transcription System Kit (Promega, Madison, WI), according to the manufacturer's instructions. Primers used for the determination of hMPV-N gene in the real-time polymerase chain reaction (PCR) were 5′-ACAGCAGATTCTAAGAAACTCAGG-′3 (Forward) and 5′-TCTTTGTCTATCTCTTCCACCC-3′ (Reverse), with an amplicon length of 153 bp. Detection of mouse β-Actin was used as the “housekeeping” reference gene. The primers used were 5′-AGGCATCCTGACCCTGAAGTA-3′ (Forward) and 5′-TCTTCATGAGGTAGTCTGTCAG-3′ (Reverse), with an amplicon of 384 bp. The products were detected using the Fast qPCR Master Mix (Applied Biosystems) in a Thermocycler StepOnePlus (Applied Biosystems). The product was detected using Brilliant QPCR Master Mix (Stratagene, La Jolla, CA) in an Mx3000P Thermocycler (Stratagene). The Relative Expression Software Tool (REST), version 2, was used to analyze real-time quantitative reverse transcription polymerase chain reaction (qRT-PCR) data for the normalized target genes (31).
Cytokine detection by ELISA
We evaluated the secretion of IFN-γ, interleukin (IL)-4, IL-10, and IL-17 in supernatants of splenocyte culture. The spleen was removed after 5 days postinfection from all mice used in this study. The spleens were perfused with PBS-1X. Subsequently, this cell suspension was centrifuged at 300 g and resuspended in RPMI 1,640 + 10% FBS. These cultures were then stimulated for 3 days with the hMPV-N (10 μg/mL). The secretion of cytokines was detected from the supernatants of splenocyte cultures utilizing the BD OptEIA ELISA Kit (BD Biosciences–Pharmingen). On day 3 of stimulation, the cell culture supernatants were recovered from the cultures for quantifying cytokine release. The cytokine detection limits were 15 pg/mL for IL-4, 30 pg/mL for IL-10, and 0.5 μg/mL for IFN-γ and IL-17.
Immunoglobulin detection by ELISA
IgG specific against the nucleoprotein was detected by ELISA from the sera of mice on day 5 postinfection. A 96-well plate was activated overnight with 200 ng/well of the nucleoprotein in a carbonate buffer medium (0.1 sodium bicarbonate [pH 9.5]) and blocked for 1 h at RT with 3% BSA in PBS-1X. The serum from the animals was added to each well in serial dilutions from 1:2 to 1:512, and then incubated to RT for 1 h. After three washes with PBS-1X, IgG Absorbance was detected with horseradish peroxidase (HRP)-conjugated anti-mouse IgG (Invitrogen). The absorbance titers in ELISA were expressed as the log of the reciprocal of the final dilution. The IgG1 and IgG2A isotypes were measured with the Mouse Ig Isotyping ELISA Kit (BD Biosciences–Pharmingen), following the manufacturer's instructions. Briefly, a 96-well plate was activated overnight at 4°C with the anti-mouse IgG capture antibody specific for each isotype. The wells were blocked for 1 h with PBS-3% BSA. The serial dilutions of serum were added to the wells and incubated for 1 h at RT. Subsequently, for detection, the Ig anti-mouse antibody marked with HRP was utilized. Detection occurred by color formation by adding a substrate provided in the kit and the optical density (OD) was determined at 450 nm.
Statistical analysis
The results are presented as the mean ± standard deviation. The statistical analysis of the data was performed using the Student's t-test. p-Values <0.05 were considered statistically significant. The analyses were performed using GraphPad Prism (GraphPad Software, Inc.). All experiments were repeated at least three times.
Results
hMPV-N expression
To purify recombinant hMPV-N protein (here as hMPV-N), encoding gene was cloned into the plasmid pET21a and expressed in E. coli BL21 Codon Plus, as described in the Materials and Methods section. The hMPV-N was expressed and purification was done using NI-NTA column (Fig. 1). Then, the purified recombinant nucleoprotein was further used to perform immunization assays in mice and immune response characterization.
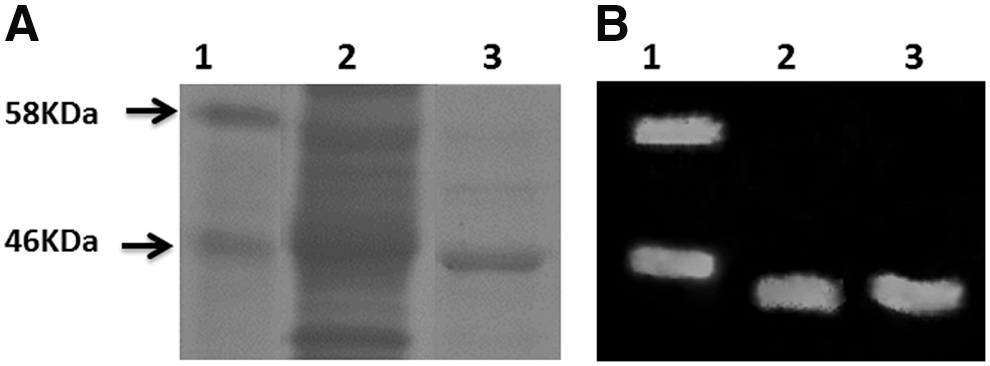
Expression of hMPV-N by Escherichia coli cells.
Immunization with nucleoprotein plus AbISCO-100 decreases the viral RNA and airway inflammation of hMPV-infected mice
We immunized BALB/cJ mice intraperitoneally (six per group), at 2-week intervals, with hMPV-N/AbISCO-100, AbISCO-100 (adjuvant control), hMPV-N, PBS-1X, Ultraviolet-inactivated virus (UV-hMPV). Also, an unvaccinated control group was added, which was not challenged with the virus (basal control group), as described in the Materials and Methods section (Fig. 2).
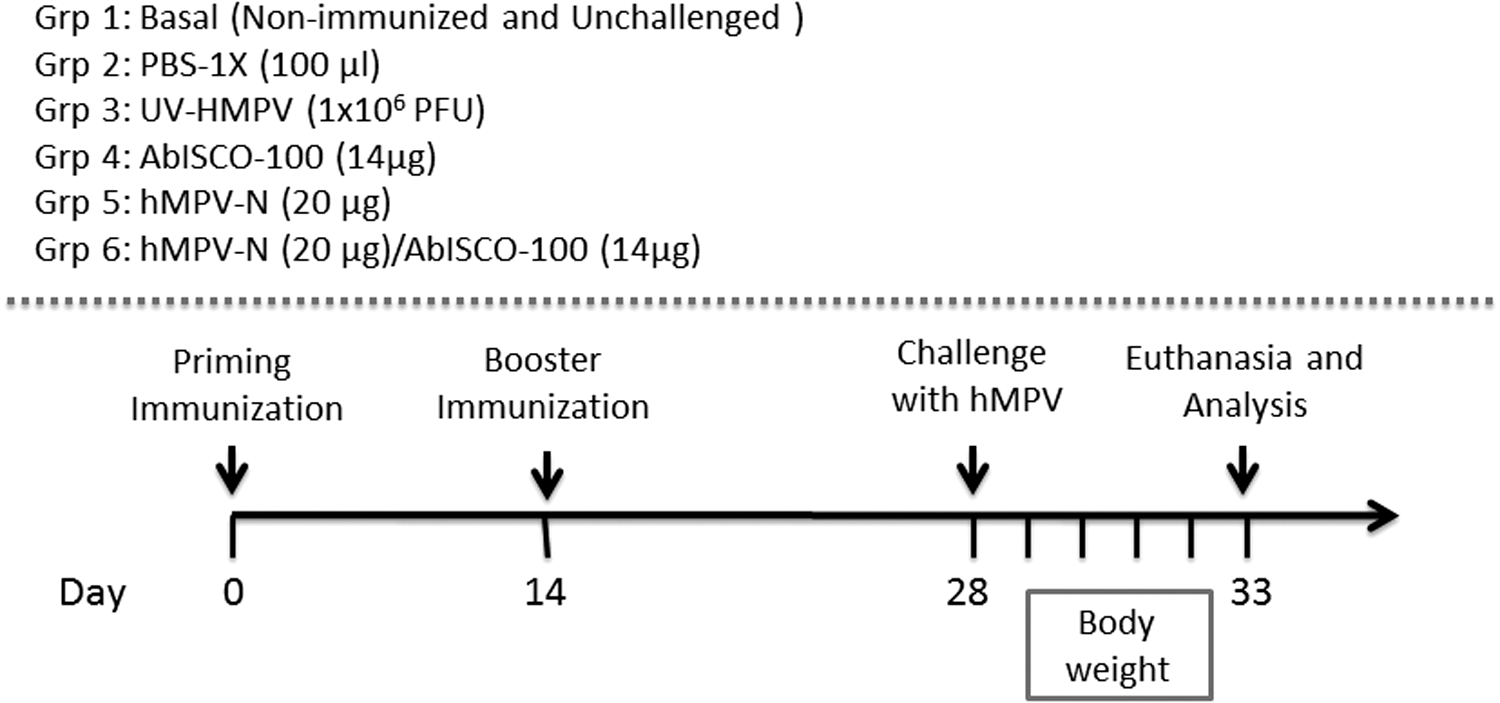
Animal immunization schedule. Schematic representation of the immunization used in the BALB/cJ mice experiment. BALB/cJ mice (six per group) were intraperitoneally immunized two times with UV-hMPV (1 × 106 PFU), hMPV-N protein (20 μg), AbISCO adjuvant (14 μg), PBS-1X (100 μL), and hMPV-N (20 μg)/AbISCO (14 μg). As basal condition, we use a nonimmunized and no-challenge group of mice. PBS, phosphate-buffered saline; PFU, plaque-forming unit.
To evaluate whether our immunization decreases hMPV infection, we evaluated the viral RNA in the lungs of the immunized mice that were subsequently infected with hMPV. On day 5 postinfection, lungs from mice were removed immediately by RNA extraction. Quantitative reverse transcription PCR (RT-qPCR) was performed to quantify the abundance of the hMPV RNA, as described in the Materials and Methods section. We observed high amounts of N-hMPV RNA in the lung of PBS-1X-, AbISCO-100-, UV-hMPV-immunized mice. Conversely, mice immunized with recombinant nucleoprotein or hMPV-N/AbISCO showed reduced viral RNA in their lungs as compared with PBS-1X-immunized mice (***p ≤ 0.0001 and *p ≤ 0.05, respectively). However, the hMPV-N/AbISCO-immunized animals showed no significant hMPV RNA as compared with unvaccinated mice (Fig. 3).
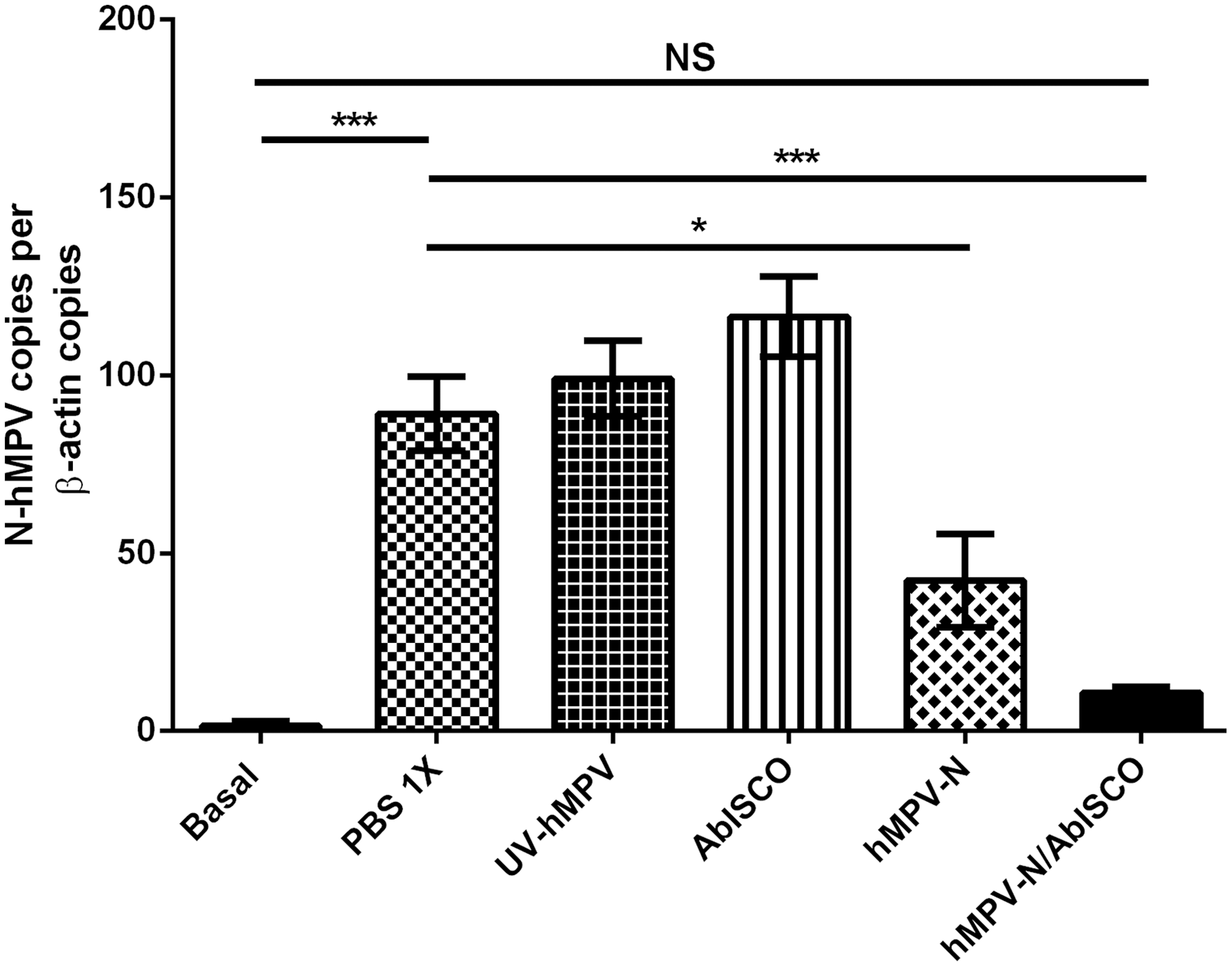
Immunization with the nucleoprotein plus AbISCO-100 decreases the viral RNA. The determination of the total viral RNA of the infected animals' lungs was detected by quantifying the number of copies of hMPV-N genes by RT-qPCR. Data are expressed as the ratio of the number of copies of the hMPV-N gene per copies of β-actin gene (not significant [ns] by Student's t-test between hMPV-N/AbISCO-immunized mice compared with basal control group; ***p < 0.0001 by Student's t-test for basal control group and hMPV-N/AbISCO-immunized compared with PBS-1X-immunized mice; *p < 0.05 by Student's t-test for hMPV-N-immunized mice compared with PBS-1X-immunized mice). RT-qPCR, quantitative reverse transcription PCR.
Because body weight loss in mice relates to the development and severity of the respiratory disease caused by hMPV, we recorded weights daily in immunized and subsequently infected mice. As expected, by day 5 postinfection, hMPV-N/AbISCO immunized mice showed an accelerated recovery of body weight compared with other infected animals reaching the weight of the basal group of mice (nonsignificant at day 5) (Fig. 4).
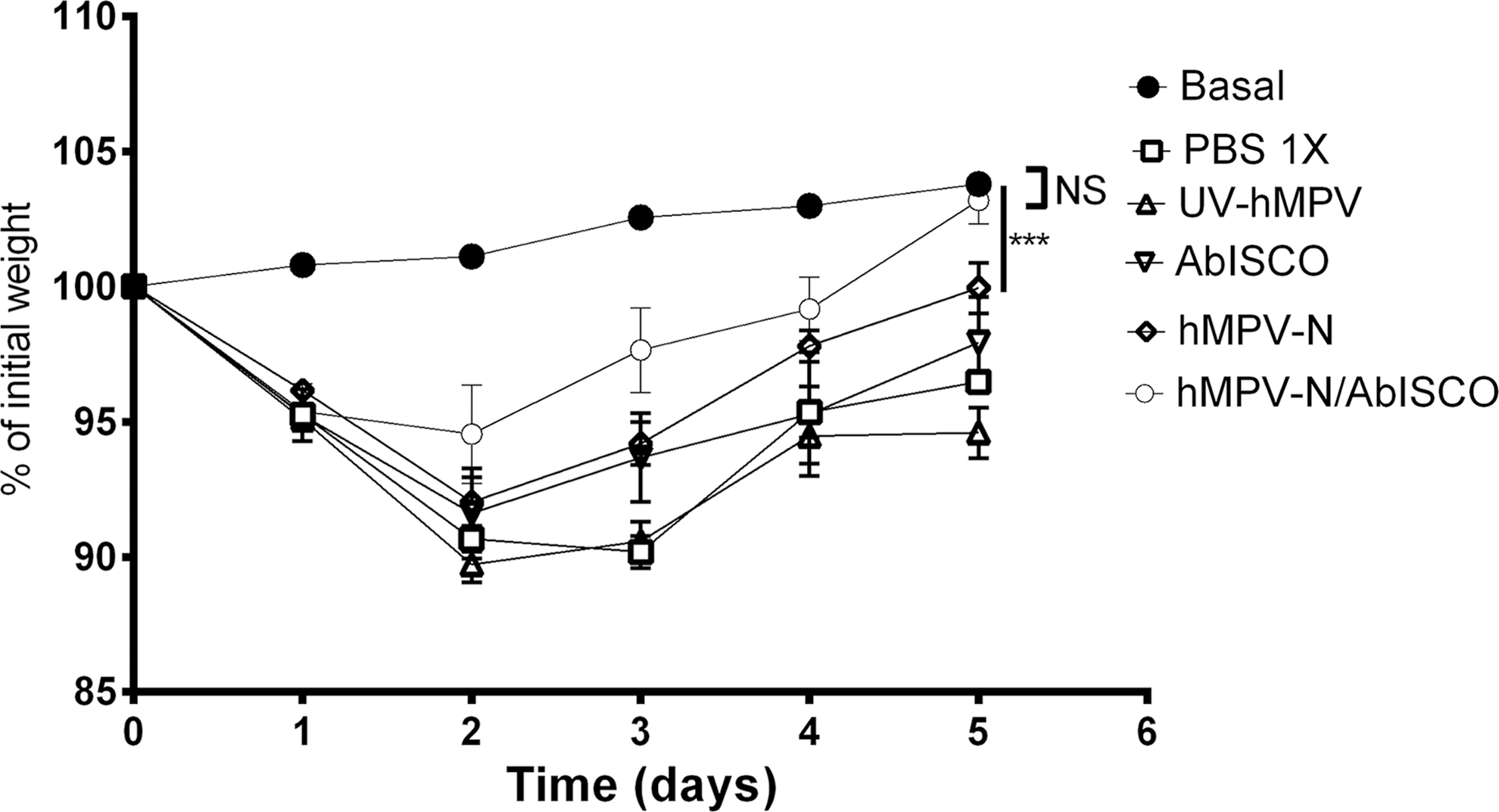
Weight loss of the mice immunized after hMPV infection. On day 5 postinfection, there were no significant weight differences recorded between the mice immunized and basal control mice (NS by Student's t-test between hMPV-N/AbISCO-immunized mice compared with basal control mice; ***p < 0.0001 by Student's t-test for hMPV-N-immunized compared with basal control mice). NS, not significant.
To study the inflammatory process in the lungs of the infected mice, we determined by flow cytometry the percentage of lung tissue following viral infection. We observed low amounts of granulocytes infiltrated in the lung of hMPV-N-, and hMPV-N/AbISCO-immunized mice compared with PBS-1X control group (p ≤ 0.01; and p ≤ 0.0001) (Fig. 5). However, we observed an equal percentage (nonsignificant, p ≥ 0.05) of granulocyte infiltration in the hMPV-N/AbISCO-immunized mice.
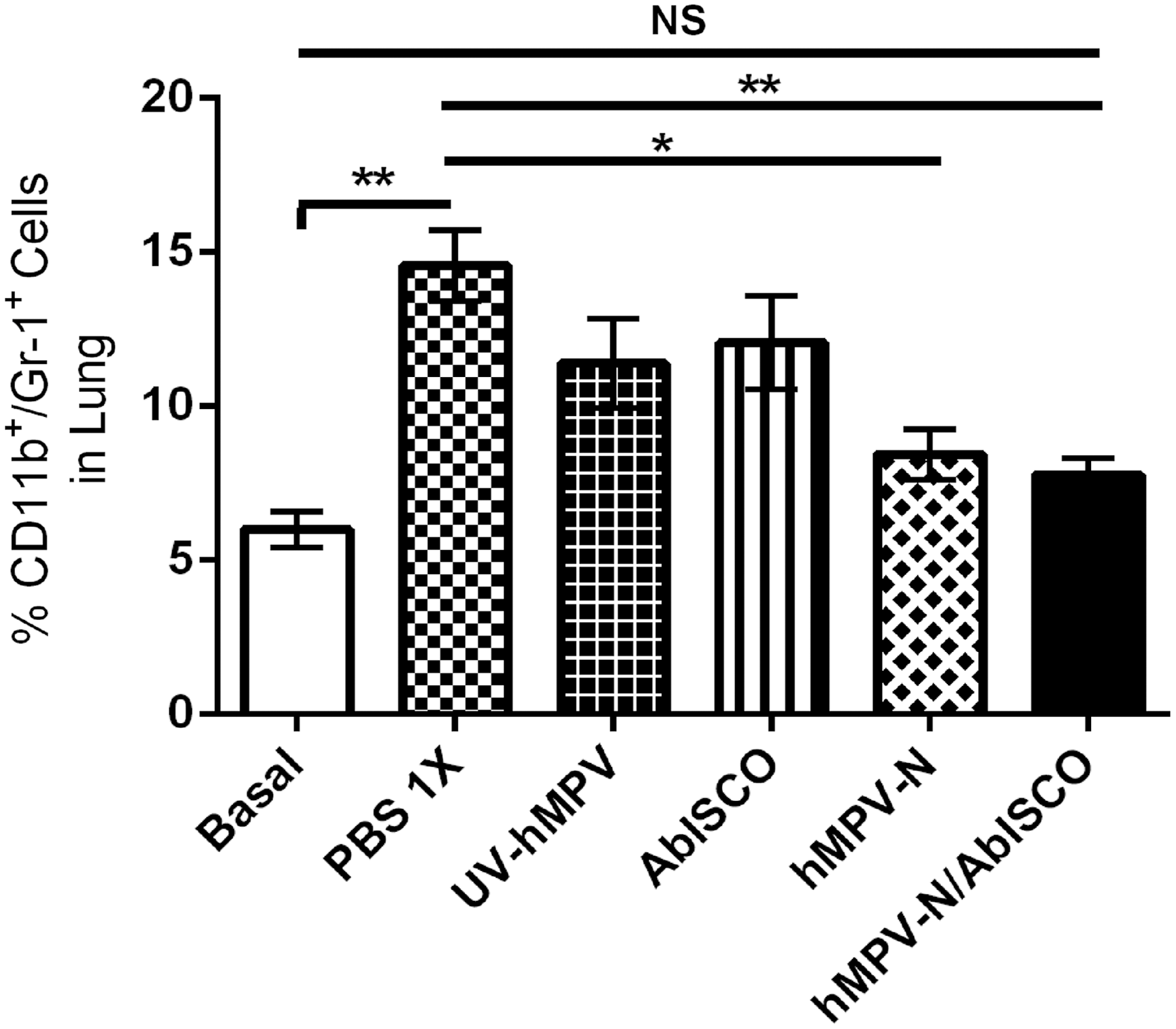
Immunization with hMPV plus AbISCO-100 nucleoprotein induces lower granulocyte infiltration in the lungs of infected mice. The recruitment of granulocytes was evaluated on day 6 postinfection with 1 × 106 PFU of hMPV. Cells recovered from the lungs were stained with FITC-labeled anti-CD11b and APC-labeled anti-Gr-1 and analyzed by flow cytometry. Cytometric quantification of the polymorphonuclear cells shows that the immunized mice underwent less polymorphonuclear cell infiltration than infected. No significant differences were observed between the immunized group and the basal control group, therefore the immunization allows the number of polymorphonuclear cells present in the lung to be brought to normal levels (not significant [ns] by Student's t-test between hMPV-N/AbISCO-immunized mice compared with basal control group; **p < 0.001 by Student's t-test for hMPV-N/AbISCO-immunized mice and basal control group compared with PBS-1X-immunized mice; *p < 0.05 by Student's t-test for hMPV-N-immunized mice compared with PBS-1X-immunized mice).
The CD11b
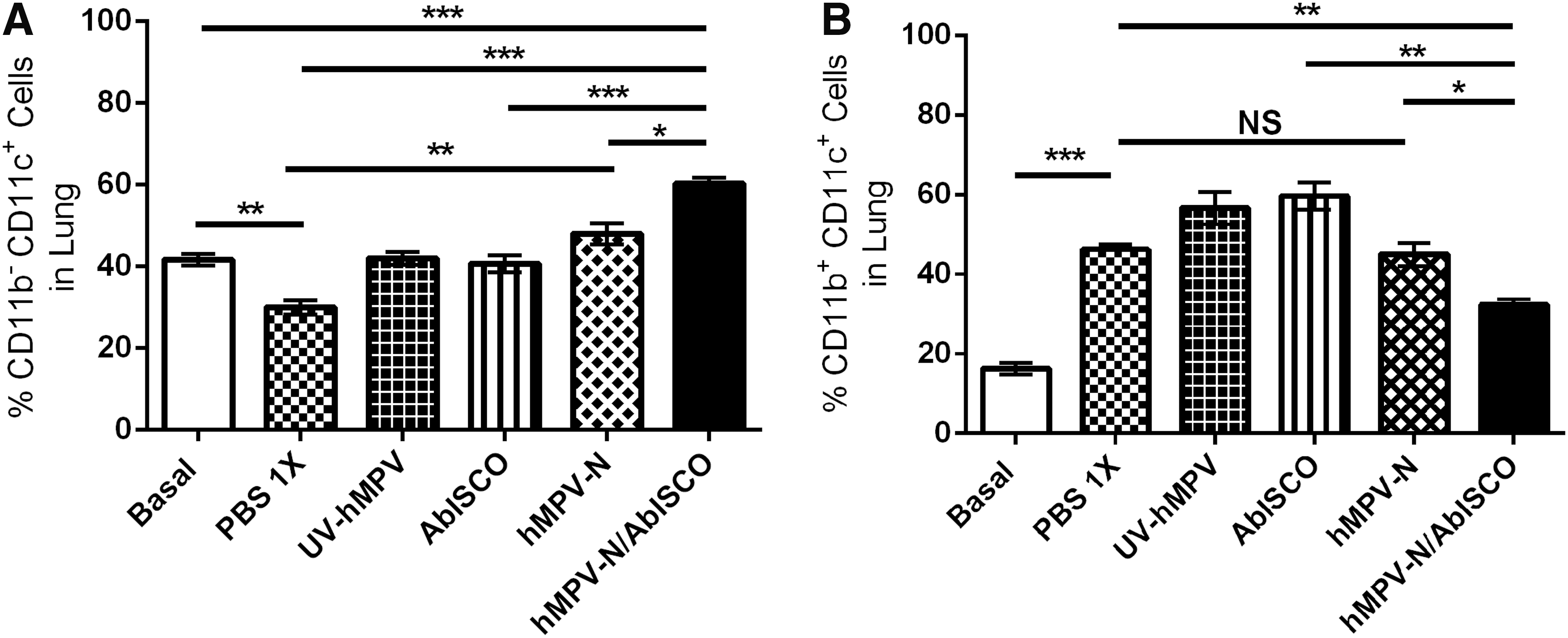
Immunization in the hMPV-N/AbISCO group affects the recruitment of dendritic cells in post-hMPV lung infection. Recruitment of dendritic cells was evaluated on day 6 postinfection with 1 × 106 PFU of hMPV.
Immunization with the hMPV-N plus AbISCO-100 induces an IgG1 and IgG2a isotype production
We studied the effect of our hMPV-N/AbISCO immunization, in the secretion in serum of IgG1 and IgG2a. Sera were obtained from the different experimental groups, according to the descriptions found in the Materials and Methods section. We evaluated IgG1 and IgG2a isotypes based on its function of Th2 and Th1 immune response, respectively. We observed an increase of the IgG2a and IgG1 isotypes, in the sera of mice immunized with hMPV-N and hMPV-N/AbISCO, compared with the remaining control groups (Fig. 7). Moreover, the hMPV-N/AbISCO-immunized mice only showed higher amounts of the isotype of IgG2a as compared with hMPV-N-immunized mice (p ≤ 0.05). These results show that antibody responses against both formulations are dependent on nucleoprotein immunization.
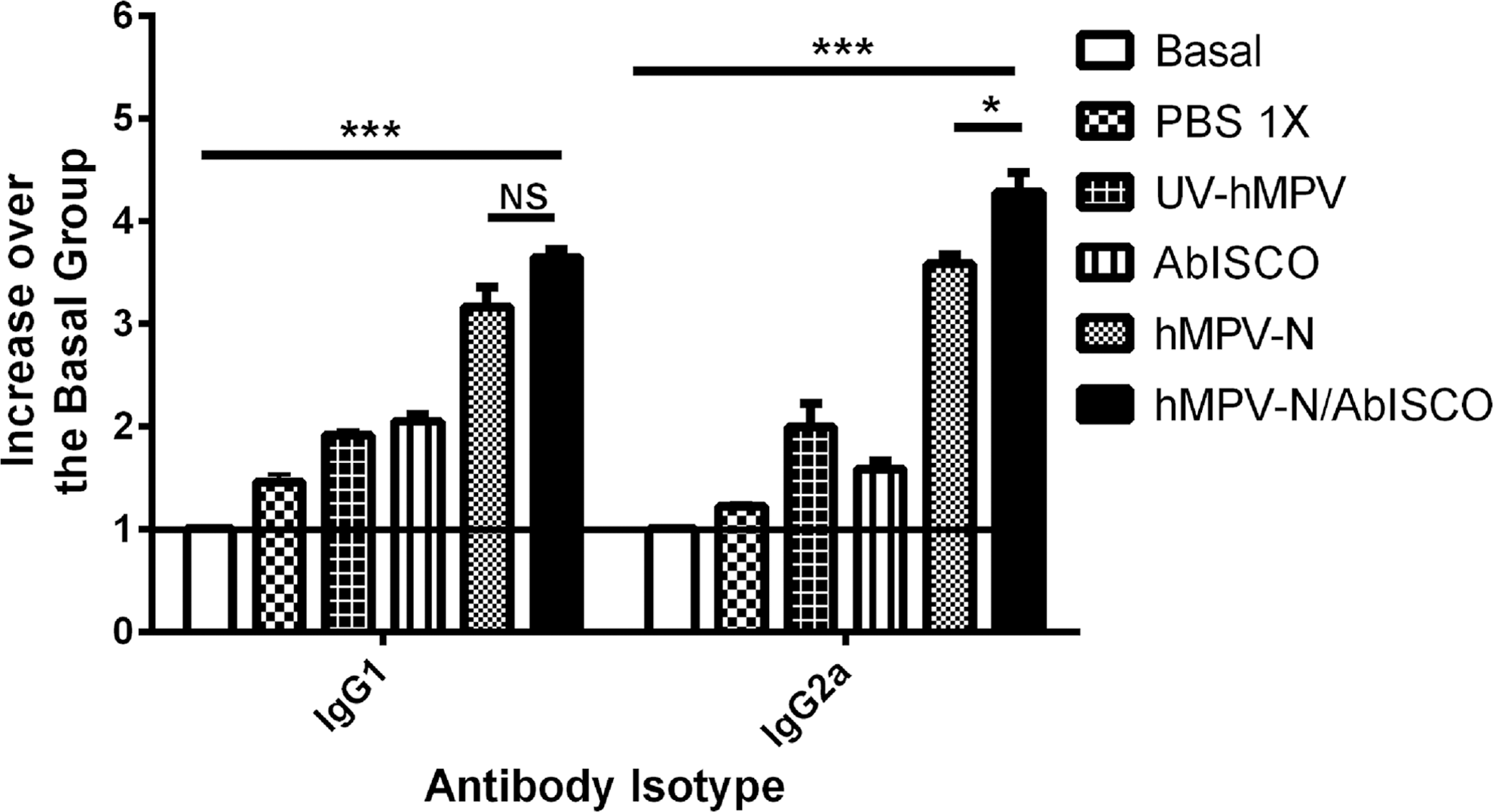
Immunization with the hMPV-N/AbISCO formulation promotes a humoral response with an increase of IgG1 and IgG2a. The detection of immunoglobulin was made using ELISA from the sera of the groups of mice infected with hMPV. Antibodies of IgG1 and IgG2a isotypes postinfection. Mice immunized with the hMPV-N/AbISCO formulation showed higher IgG1 and IgG2a isotypes in serum of BALB/cJ mice compared with the groups of mice immunized with PBS-1X, UV-hMPV, AbISCO (***p < 0.0001 by Student's t-test for basal control group compared with hMPV-N/AbISCO-immunized mice; *p < 0.05 by Student's t-test for hMPV-N-immunized mice compared with hMPV-N/AbISCO-immunized mice; not significant [ns] by Student's t-test between hMPV-N-immunized mice compared with PBS-1X-immunized mice).
The immunization induces Th1/Th17 cytokines profile in splenocytes culture
To characterize the T cell cytokine profile induced by the hMPV-N/AbISCO, we determined by ELISA the relative amount of IFN-γ, IL-17A, IL-10, and IL-4 in the culture supernatants of splenocytes stimulated with recombinant nucleoprotein, as described in Material and Methods section. We found that upon stimulation with the nucleoprotein, the splenocytes of the hMPV-N/AbISCO group secreted higher relative amounts of IFN-γ, IL-17A, and IL-10 (Fig. 8A–C). The amounts of IL-4 were significantly lower in mice immunized with the hMPV-N/AbISCO, AbISCO-100, and hMPV-N, compared with PBS-1X control group (Fig. 8D). Furthermore, the groups immunized with AbISCO-100 generate a decrease in the amounts of IL-4 compared with PBS-1X control group, which is consistent with previous studies (36). However, the splenocytes of the group immunized with the hMPV-N also generate a decrease in IL-4 levels compared with PBS-1X control group, and this suggests a Th2-biasing effect of the protein alone (Fig. 8). These data allow us to conclude that the hMPV-N/AbISCO immunization induces a mixed Th1/Th17 repertoire of T cells secreting large amounts of IFN-γ, IL-10, and IL-17A upon recall compared with the other experimental groups.
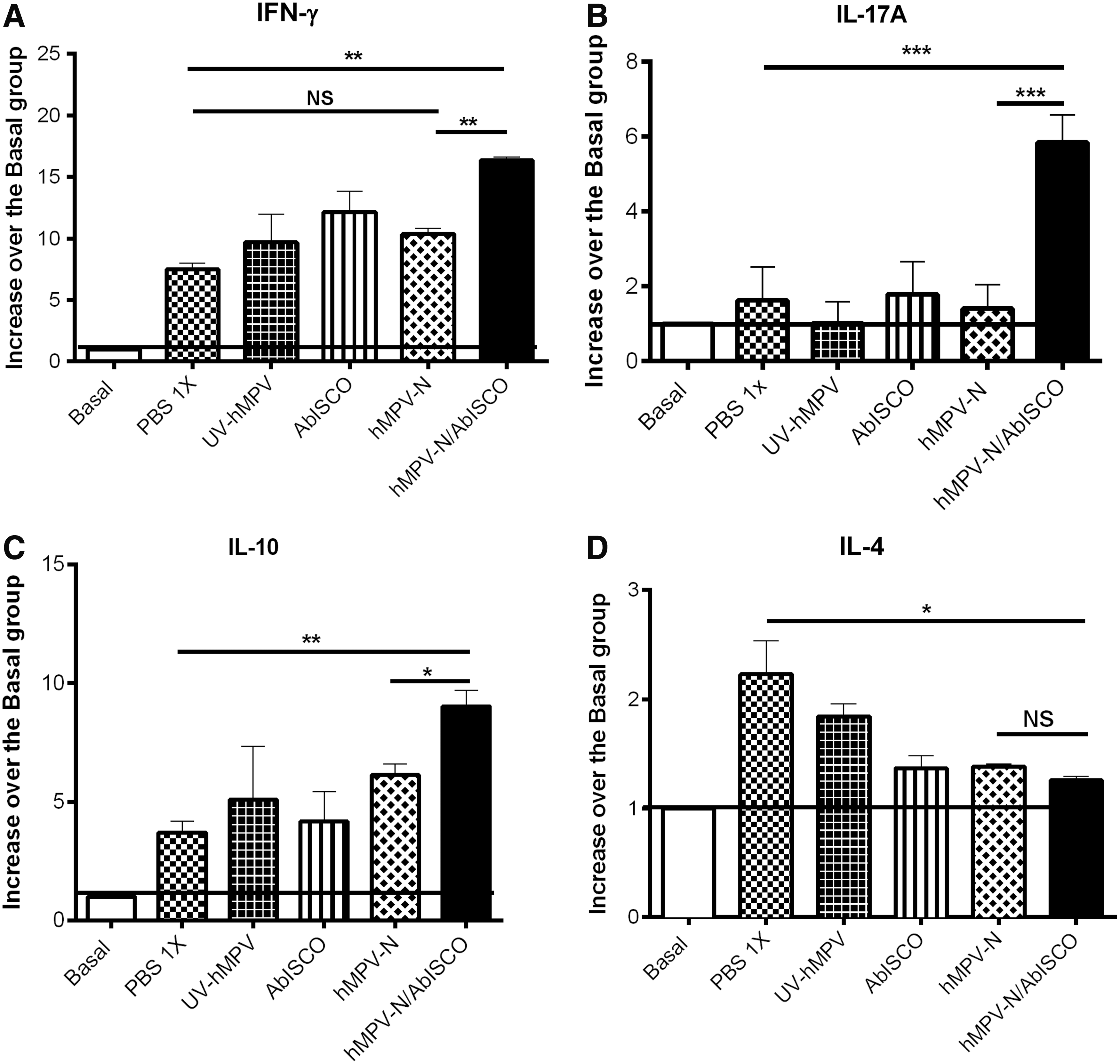
Cytokine production in the splenocytes of mice immunized with hMPV-N/AbISCO. Increase of IFN-γ, IL-17A, IL-10, and IL-4 release to the cell culture supernatants as fold increase over basal control groups. The production of IFN-γ, IL-17A, IL-10, and IL-4 was quantified by ELISA from splenocyte culture supernatants of immunized and basal control mice that were sacrificed after 5 days postinfection with hMPV. Levels of
Discussion
hMPV is currently a significant global public health burden and the development of a vaccine is a major concern (34). To date, several vaccine prototypes have been formulated and evaluated, including those based on inactivated chimerical virus, which later has harmful results on the respiratory system after viral infection (27,40). On the other hand, vaccine prototypes have been evaluated in base hMPV recombinant proteins, or smaller predictors (10,24,27,33); an immunization strategy that is considered safe against viral infection. In this work, we show that protein immunization composed of the hMPV-N plus the AbISCO-100 adjuvant induces a decrease of viral infection characterized by a Th1/Th17-type response; also we observed a decreasing of symptomatology to viral infection in the animal model.
The disease arising from hMPV infection in BALB/cJ mice are characterized by weight loss, viral replication, and granulocyte infiltration in the lungs (1,7,11,20). In our experimental results, we observed a decrease in the parameters described above, indicating that hMPV-N/AbISCO immunization prevents the viral infection. It has been shown that the severity of the respiratory disease is correlated with the phenotype of the dendritic cells infiltrated in the lungs after viral infection (5,18). Specifically, the CD11b+CD11c+ phenotype has a proinflammatory function that induces damage to the lung tissue (13). On the contrary, it has been described that the CD11b
When analyzing the secretion of splenocytes from mice immunized with hMPV-N/AbISCO and pulsed with nucleoprotein, we observed a mixed Th1/Th17 repertoire being expanded in response to the antigen, with a concomitant significant secretion of IL-10. This cytokine profile could be associated with a better immune response against hMPV. The Th1 is a specialized response in the fight against intracellular pathogens, which allows the activation of CD8+ T cells that are crucial in the elimination of persistently infected cells, and the activation of macrophages with antiviral activity. Otherwise, immunization with hMPV proteins requires investigations on the effects of Th1/Th2/Th17 balance (32). The secretion of IL-17A has been recently reported as necessary for the subsequent infiltration of Th1 profile cells (21,25). On the other hand, the Th1 response of hMPV-N/AbISCO immunization is associated with the secretion of IFN-γ in conjunction with a high secretion of IL-10, which suggests the participation of regulatory populations and the development of a more regulated immune response capable of clearing the virus from the airways while preventing exacerbated tissue damage and disease.
In the case of the closely related virus, human respiratory syncytial virus (hRSV), IgG1 and IgG2a immunoglobulins contribute to virus neutralization and elimination (19,39). Moreover, the presence of higher levels of IgG2a isotype against hMPV infection in our study is consistent with Th1-type cytokine profile described in the Results section. In the murine model, the IgG2a isotype may be mediating complement activation by the classical pathway and opsonization leading to phagocytosis by neutrophils and macrophages (29). The IgG2a isotype could be contributing to the elimination of infected cells by mediating antibody-dependent cell cytotoxicity (22).
The use of purified protein as vaccine nonadjuvanted has proven to be safe and immunogenic against pulmonary disease (15). In our work, although immunization with the recombinant hMPV-N protein alone decreased the amount of viral RNA in mice infected by hMPV, it failed at preventing weight reduction, at reducing the CD11b+CD11c+ antigen-presenting cells, and at inducing T cell cytokine profile production against infection. Therefore, the combination of hMPV-N and adjuvant AbISCO-100 was required to improve the immune response against viral infection. Consistent with the foregoing, ISCOMATRIX adjuvant is a potent immune modifier, linking innate and adaptive immunity and creating a favorable milieu for Th1 type immune responses (14). Moreover, the ISCOMATRIX adjuvant has been tested in a number of experimental settings, including studies of therapies against cancer and malaria, as well as in antiviral vaccines (8,14,38). Finally, the hMPV-N/AbISCO immunization described in this study could be an attractive formulation for the development of a vaccine against hMPV infection.
Footnotes
Acknowledgments
This research was funded by P IMII P09/016-F, funded by the ICM Program granted by A.M. Kalergis and with the institutional support by A.E. Vásquez of the Instituto de Salud Pública de Chile (Institute of Public Health in Santiago, Chile). The authors recognize Bárbara Riveros and América Abarca for their technical assistance and Luis Rodríguez for support in the statistical analysis.
Author Disclosure Statement
No competing financial interests exist.