
Abstract
This study focuses on preparing the secretory recombinant J subgroup of avian leukosis virus (ALV-J) gp85 protein using Pichia pastoris and evaluating its immunoprotection as vaccine antigen combining with CpG-ODN adjuvant. The secretory recombinant plasmid pPIC9-gp85 containing ALV-J gp85 gene was designed and was transfected into the genome of P. pastoris (GS115) cells. The recombinant plasmid was expressed under the induction of methanol. The expressed products in the medium of the cells were purified and identified with endoglycosidase digestion assay and western blot mediated with monoclonal antibody (MAb) JE9. The purified product combining with CpG-ODN adjuvant was inoculated intramuscularly into 7-day-old chickens and three booster inoculations were performed on 21 days post first inoculation (dpfi), 42, and 56 dpfi. The antibody responses and cellular immune responses were detected, and the protective effects were analyzed after challenge with ALV-J. The results showed that the secretory pPIC9-gp85 plasmid was successfully constructed and could be stably expressed in GS115 cells. The expressed products were N-acetylglucosylated and could specifically combine with MAb (JE9). The secreted gp85 protein combining with CpG-ODN adjuvant could induce higher antibody response and spleen lymphocyte proliferation response and IFN-γ-inducing response, and could protect all the inoculated chickens against the viremia and the immunosuppressive lesions caused by ALV-J challenge. The results of neutralizing test in vitro suggested that the antisera with some ALV-J antibody titers could neutralize ALV-J strain and inhibit the growth of virus in vitro. The result of IFA showed that IgG antibody in the antisera could specifically combine with ALV-J strain in cells. It can be concluded that the secretory recombinant gp85 protein, as a new acetylglucosylated gp85 protein, was successfully prepared and combining with CpG-ODN adjuvant could protect the inoculated chickens against ALV-J infection. This study first reported the methods on preparing the secretory recombinant ALV-J gp85 protein using P. pastoris and evaluated its immunoprotection.
Introduction
A
Methylotrophic yeast Pichia pastoris is an effective expression system with the advantages of easy manipulation, high production levels, and low culture cost. Moreover, due to culture at high cell densities, it can be used in the production of recombinant proteins to replace E. coli expression system (4,6,23). Prepared proteins using this method also have better bioactivity due to the glycosylation and structural folding of the expressed products (13,16,30). Furthermore, the expressed recombinant products could be secreted into the supernatant medium of P. pastoris cells (2,32). Interestingly, whether the recombinant ALV-J gp85 protein expressed using this method can be used as subunit vaccine antigen has yet to be determined. In this study, we prepared the secretory recombinant gp85 protein using P. pastoris and evaluated its immunoprotection combining with CpG-ODN adjuvant against ALV-J infection in chickens.
Materials and Methods
Viral reference strain, antibodies, and adjuvant
The ALV-J-NX0101 strain was isolated from a commercial broiler chicken house in Ningxia Province, China (8). The ALV-J gp85-specific mouse monoclonal antibody (MAb) JE9 was a generous gift from Prof. Zhizhong Cui, and the anti-chicken and anti-mouse IgG-FITC conjugates were obtained from Sigma, USA, and Bangalore Genei, respectively. The CpG-ODN sequence TCGTCGTTTTGTCG TTTTGTCGTT was synthesized at Shanghai Sangon Biological Engineering Technology & Services Co., Ltd. (Shanghai, China) and was used as adjuvant for gp85 subunit vaccine in this study.
Host strains and plasmid vectors
The P. pastoris host strain used in the study was a histidine requiring auxotroph GS115 (His4). The plasmid vector pPIC-9 and yeast transfer vector contained the P. pastoris AOX1 promoter and transcription termination sequences separated by MCS for insertion of a foreign gene of interest. The GS115 (His4) host strain and plasmid pPIC-9 were obtained from Invitrogen.
Construction of the secretory recombinant plasmid pPIC9-gp85
The primers were designed according to the ALV-J-NX0101 gp85 gene sequence published in GenBank (No. AY897227) and synthesized. The gp85 gene was amplified using the following primers with proviral cDNA extracted from ALV-J-infected cells as the template. The forward primer 5-CCC
The polymerase chain reaction (PCR) product was cloned into the pMD18T vector and amplified in E. coli (TOP10). The amplified PCR product corresponding to truncated gp85 was gel purified using the wizard PCR preps DNA purification system (Promega) and cloned into the pPIC-9 yeast transfer vector (Invitrogen) at EcoRI and NotI sites according to our designed figure (Fig. 1). The recombinant colonies grown on ampicillin plates were initially screened by colony PCR, using vector-specific 5′AOXI (5′-GACTGGTTCCA ATTGACAAGC-3′) and 3′AOXI (5′-GCAAATGGCATTCTGACATCC-3′) primers (Invitrogen), followed by restriction enzyme digestion of plasmid DNA. The gp85 gene insert into the pPIC-9 vector was confirmed using DNA sequencing.
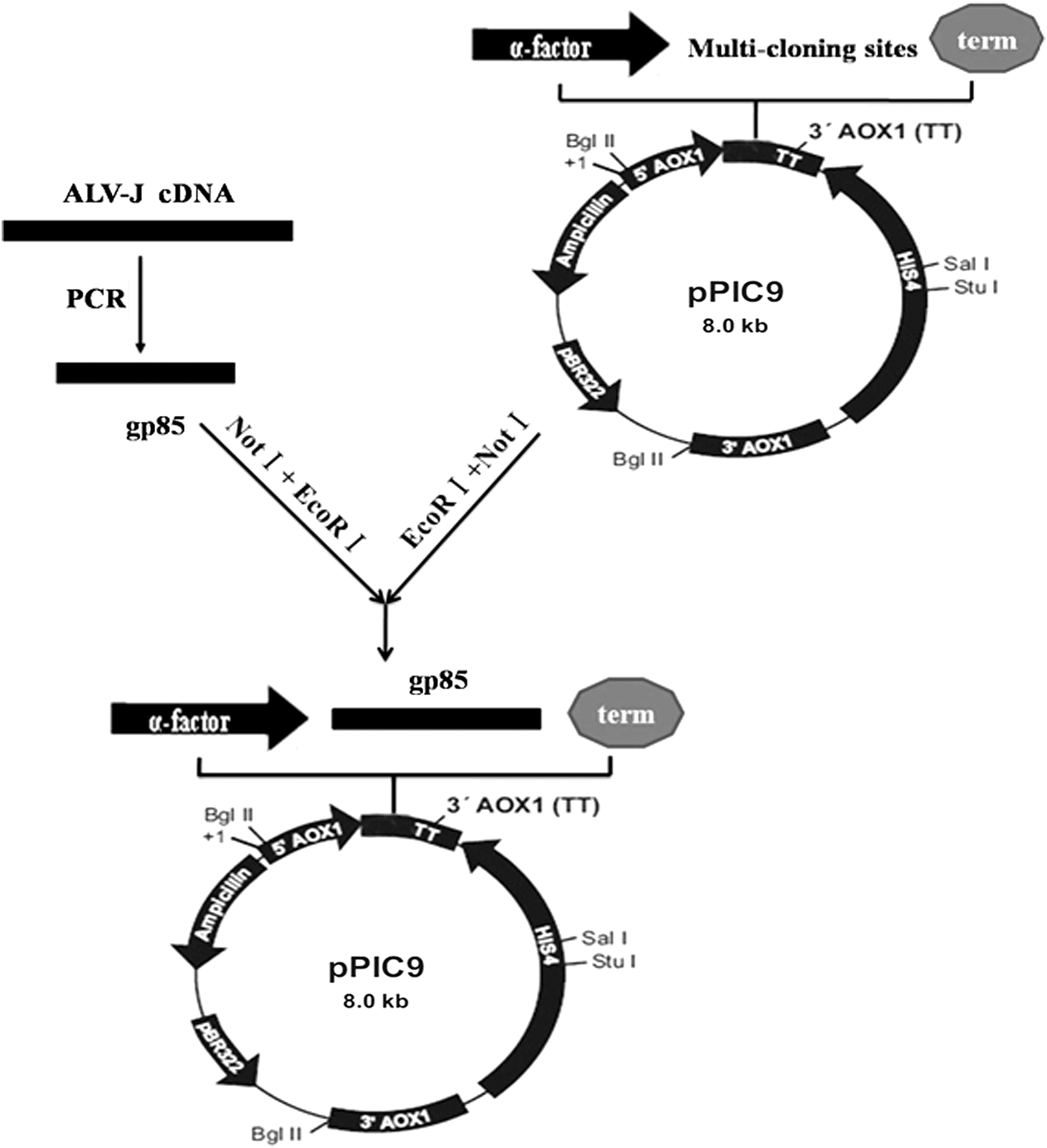
Construction map of the recombinant expressed plasmid pPIC9-gp85 by inserting gp85 gene of ALV-J at the EcoRI and NotI sites of pPIC9 vector. ALV-J, J subgroup of avian leukosis virus.
Preparation and purification of the secretory recombinant gp85 protein using P. pastoris
The recombinant ALV-J gp85 gene was expressed in P. pastoris cells according to previously published methods (1). Briefly, the plasmid with gp85 insert was linearized with Sa1 I and electroporated into the GS115 cell strain of Pichia using the Bio-Rad Gene Pulser. The positive cells were harvested and resuspended in Buffered Methanol-Complex Medium. The flasks were incubated at 30°C to continue growth, and methanol was added to a final concentration of 0.5% every 24 h to sustain the induction for 96 h. After 96 h, the entire culture supernatant was harvested.
The purification procedures were performed according to the method in the reference (13) and with several modifications as follows. The supernatant was collected after the cultures were induced. To determine the optimum precipitation conditions, ammonium sulfate powder was added to the supernatant at 4°C to adjust solutions to final saturations of 50%, and kept at 4°C overnight. The precipitated proteins were collected and dissolved in 10 mM phosphate-buffered saline (PBS) and desalted against 1× PBS. The protein content was estimated using the dye-binding assay. The lyophilized protein was further concentrated and dissolved in an equilibration buffer (PBS, pH 7.4) and purified by two gel filtration runs using SD200 Nickel column (GenScript USA, Inc., Nanjing, China). The resin was washed with 50, 100, 200, and 500 mM imidazole, respectively. Flow-through material and elution fraction were collected respectively, and analyzed by SDS-PAGE in 12% polyacrylamide gels.
Endoglycosidase digestion assay and western blot analyses
To further identify the glycosylation of the expressed products, the purified protein product was digested with specified endo-β-N-acetylglucosaminidase H (Endo-H) as described previously (31) and the digested products were analyzed by SDS-PAGE. The products before and after being digested were identified using western blot mediated with MAb (JE9).
The protein concentration was determined by performing thin-layer chromatography scanning and the Bradford total protein content assay using the Bio-Rad Protein Assay Kit (Bio-Rad); bovine serum albumin was used as the standard.
Immunization of chickens
Seven-day-old SPF chickens were randomly divided into four groups: control group, CpG group, gp85 group, and gp85+CpG group. Each group contained not less than 13 chickens, and the chickens were maintained in the dedicated SPF animal room. The chickens in each group were injected intramuscularly four times at 0, 21, 42, and 56 days post first inoculation (dpfi). Each chicken in the control group was inoculated with the emulsions containing PBS as the negative control, and in CpG group was inoculated with the emulsions containing 50 μg CpG-ODN as the adjuvant control, and in gp85 group was inoculated with the emulsions containing 100 μg purified gp85 protein, and in gp85+CpG group was inoculated with the emulsions containing 100 μg purified gp85 protein plus 50 μg CpG-ODN adjuvant. After the first vaccination, the serum samples were collected from each chicken at a 7-day-interval until 77 dpfi. The animal experiments were approved by the Animal Ethics Committee at the Shangdong Province Animal Protection and Welfare Institute (No. SDAU-201509).
Evaluation of immunoprotection against ALV-J challenge
Except some of the inoculated chickens on 56 dpfi that were used to analyze cellular immune responses, the other chickens were challenged intraperitoneally with 103.625 TCID50 of the ALV-JNX0101 strain on 77 dpfi. The viremia and the immunosuppressive lesions caused by ALV-J challenge were detected at the third week post challenge.
Indirect ELISA for ALV-J antibody detection
The commercial ALV-J Antibody Test Kit (IDEXX USA, Inc., Beijing, China) was used according to the manufacturer's protocol to determine ALV-J antibody titers in the sera. The relative antibody titers were determined by calculating the sample to positive (S/P) ratio [(mean of sample optical density) − (mean of negative control optical density)]/[(mean of positive control optical density) − (mean of negative control optical density)]. The sera from each group were tested in triplicate and the serum sample with a S/P ratio not less than 0.6 was considered ALV-J antibody positive.
ALV-J viremia detection
The serum samples were collected from each challenged chickens and were checked for the presence of the virus using the ALV P27 Antigen ELISA Test Kits (IDEXX USA, Inc.). The relative P27 antigen titers were determined by calculating the S/P ratio using the formula shown in Indirect ELISA for ALV-J Antibody Detection section. The samples from each group were tested in triplicate, and the serum samples with an S/P ratio higher than 0.2 were considered virus positive.
Detection of immunosuppressive lesions in chickens
Each of the challenged chickens was weighed on 21 days post challenge, and the thymus, the spleen, and the bursa were weighed, and their development indices (DI) were calculated using the following equation: (organ weight)/(body weight) × 100%, respectively.
The neutralization test of antisera
The antisera of the vaccinated chickens were randomly obtained from the control group, gp85 group, and gp85+CpG group on 63 dpfi and were used to analyze their neutralization against ALV-J strain. The essential procedure was similar to that described by Rweyemamu et al. (26) with modifications. The antisera with some titers were diluted in Eagle's maintenance medium in a twofold dilution starting from 1:4 to 1:16 in flat-bottomed tissue culture 96-well plates. The viral suspension, which was obtained from the ALV-JNX0101 strain with titers of 102.625 TCID50/0.1 mL, was added to each antiserum well, and the mixture was incubated for 2 h at 37.8°C in a humidified CO2 incubator. A 50 mL DF1 cell suspension (∼1.5 × 106/mL) was added to each well and further incubated. The appropriate serum, virus, and cell controls were included in the test. The cell culture supernatants on 1 day post incubation (dpi), 3 dpi, and 5 dpi, were confirmed for the presence of the virus using the ALV P27 Antigen ELISA Test Kits (IDEXX USA, Inc.). The relative antigen titers were determined by calculating the S/P ratio using the formula shown in ALV-J Viremia Detection section. Samples from each group were tested in triplicate and the plasma sample with an S/P ratio higher than 0.2 was considered virus positive.
IFA of specific antibody
ALV-J-NX0101 strain was grown in DF1 cells in six-well plates and was used to identify the specific antibody in the serum using IFA. Briefly, DF1 cells containing virus (with P27 antigen titers more than 1.5) were dipped into the confining liquid for 30 min. The confining liquid was discarded, and the antisera (dilution 1:20) obtained from the chickens in gp85+CpG group or in gp85 group or in control group were added directly to DF1 cells, without washing, for 2 h at 37°C. The tissue samples were washed three times with PBS and the FITC-labeled anti-chicken IgG antibody (dilution 1:200; Sigma) was added. The samples were placed in a wet box for 2 h at 37°C, washed three times with PBS, and a small amount of 50% PBS glycerol mounting solution was added. The samples were viewed under a fluorescence microscope. Samples exhibiting a green fluorescence in the cytoplasm were considered positive for ALV-J.
Lymphocyte proliferation assay
The essential procedure of lymphocyte proliferation assay was similar to that described by Li et al. (17). The splenic lymphocytes were isolated aseptically from six chickens of each group using the Chicken Lymphocyte Separation Kit (Dakewe, China) at 56 dpfi. The lymphocytes were seeded into 96-well plates at 4 × 105 cells per well, the cultures were stimulated for 48 h with concanavalin A (ConA, 5 μg/mL; Sigma) as positive control, purified recombinant gp85 protein (rgp85) at 5 μg/mL as the specific antigen, or medium as negative control at 37°C with 5% CO2. The proliferative response was measured by the Cell Counting Kit-8 (CCK-8; Dojindo Laboratories, Kumamoto, Japan). The optical density (OD) of each well was determined at 450 nm. The stimulation index (SI) was calculated as the ratio of the average OD of antigen-stimulated cells to the average OD of unstimulated cells.
IFN-γ release assay
Chicken splenic lymphocytes were prepared as above on 56 dpfi and were incubated in 96-well plates at 4 × 105 cells per well. Triplicate wells containing medium alone and 5 μg/mL rgp85 protein were incubated for 48 h, and 72 h before culture supernatants were collected. The concentrations of IFN-γ in the supernatant were detected with the chicken IFN-γ ELISA Kits (Sino Biological Inc., Beijing, China) according to the manufacturer's instructions.
Data analysis
Data are represented as the mean ± standard deviation (
Results
The construction of the secretory recombinant plasmid containing ALV-J gp85 gene
To obtain the secretory recombinant plasmid containing ALV-J gp85 gene according to the constructed map in Figure 1, ALV-J gp85 gene was first amplified from proviral cDNA of ALV-J using the designed primers. The results in Figure 2A and the sequencing result showed that it contained 945 bp of gene. Then gp85 gene was successfully inserted into the specified site of transfer plasmid pPIC9 in Figure 2B and the result of sequencing showed that the recombinant expressed plasmid pPIC9-gp85 was 1,437 bp of gene containing 945 bp of gp85 and 492 bp of pPIC9 vector. Third, the result in Figure 2C showed that two specified bands, including 1,437 bp of pPIC9-gp85 recombinant plasmid and 2,200 bp of AVOX1 gene could be amplified from the positive GS115 cells, which suggested that the linearized recombinant expressed plasmid pPIC9-gp85 was successfully transfected into the genome of GS115 cells. The results of sequencing also confirmed that (See Supplementary Data; Supplementary Data are available online at
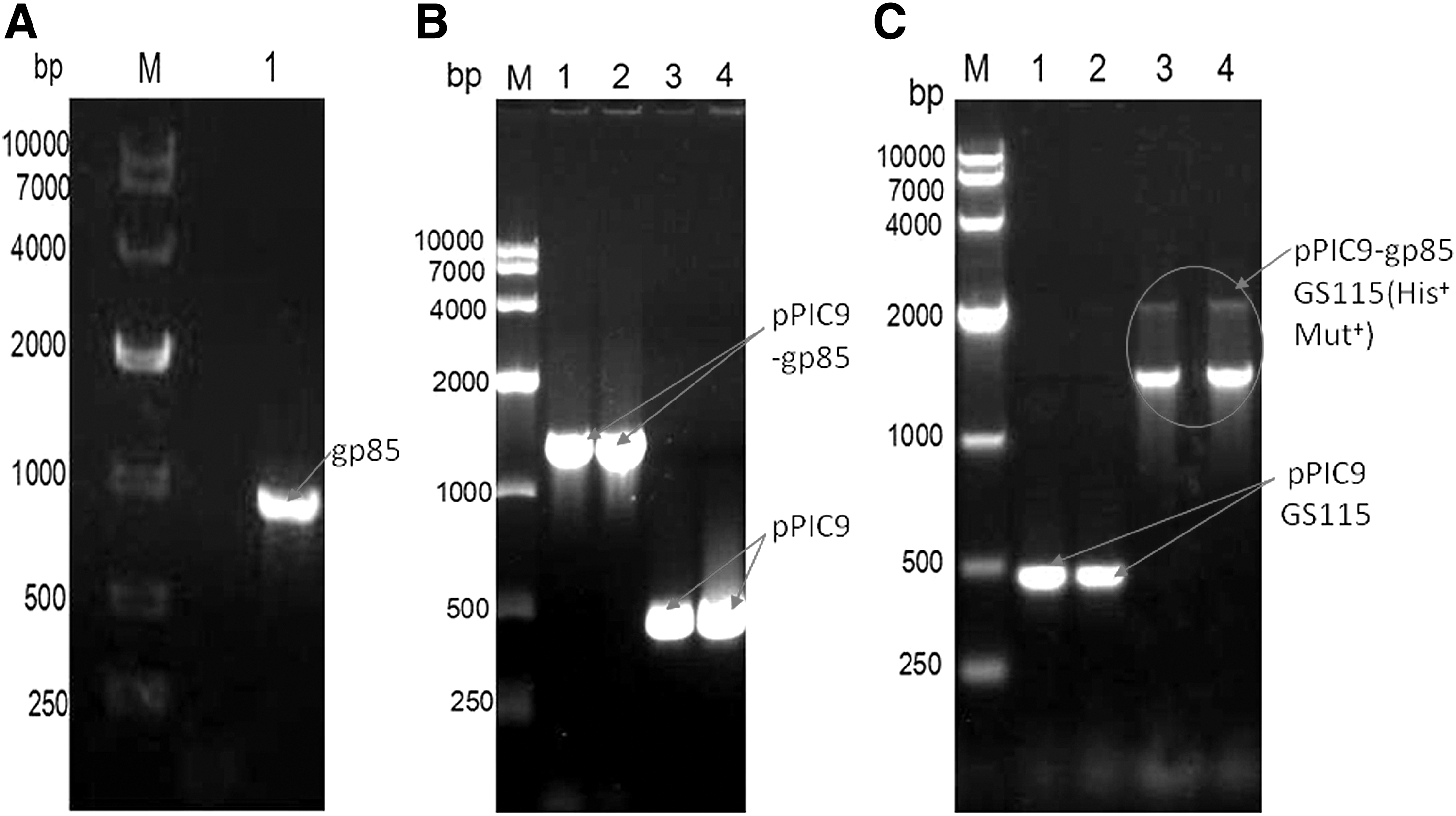
Construction and identification of the recombinant expressed plasmid pPIC9-gp85 containing ALV-J gp85 gene.
Expression, purification, and glycosylation analysis of the secretory recombinant gp85 protein
To prepare the secretory recombinant protein containing gp85 protein using the above constructed plasmid, the PCR-positive Pichia clones were grown separately and induced for expression with 0.5% methanol. Because the inserted gene was present downstream of the secretory signal sequences, the expressed gene product was expected to be secreted into the medium. Thus, the cells' supernatant was collected at 96 h and was purified using high-affinity chromatography followed by gel filtration chromatography. The results of SDS-PAGE showed a purified protein band of ∼50 kDa (Fig. 3A, lane 2). The purified protein was digested with endo-β-N-acetylglucosaminidase H and showed two specified protein bands, including the protein band of ∼50 kDa and the protein band of ∼30 kDa (Fig. 3A, lane 1), which suggested that the secretory recombinant protein was N-acetylglucosylated.
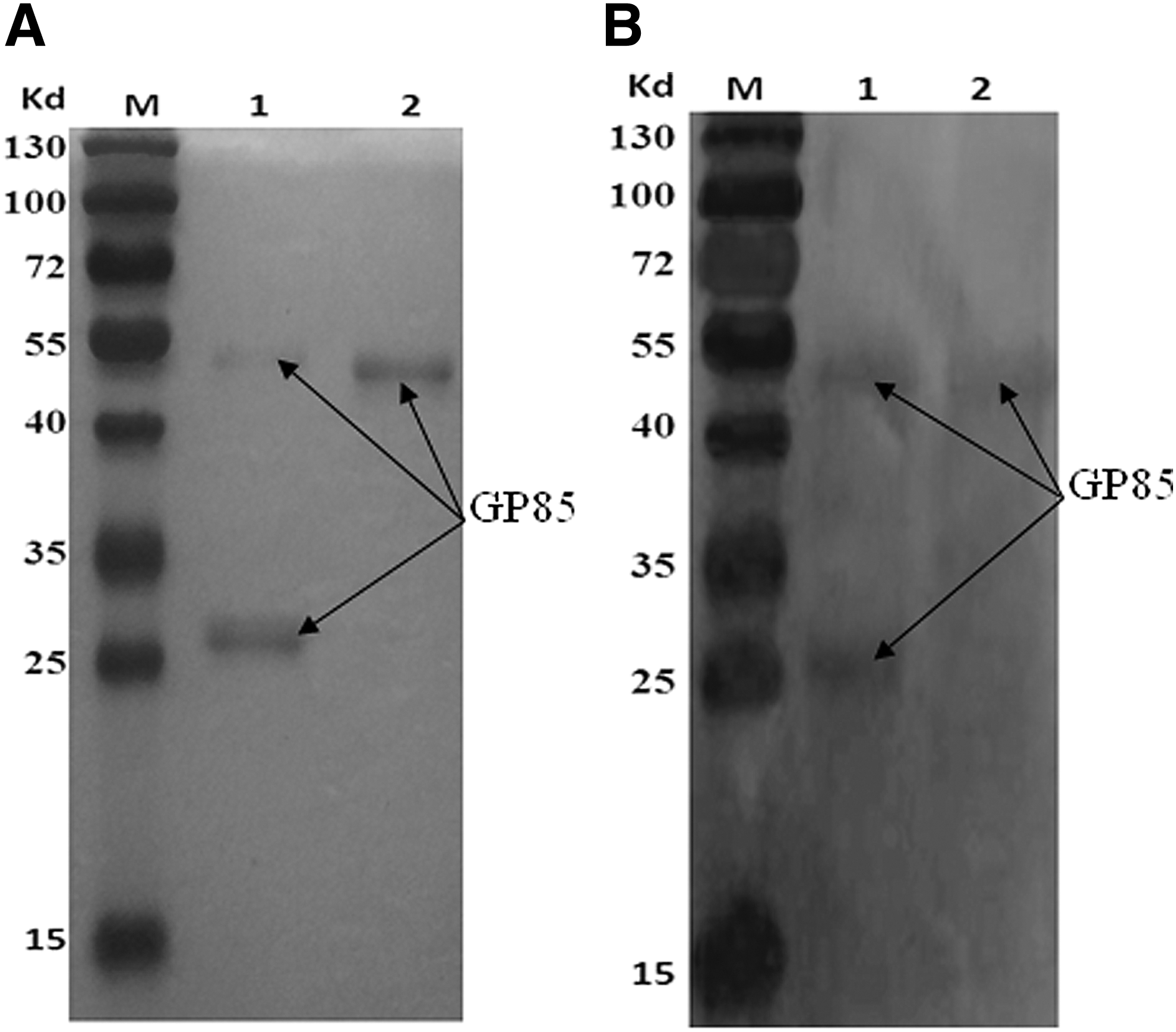
Identification of glycosylation
Western blot analysis of the secretory recombinant gp85 protein
To confirm the glycosylation of the secretory recombinant protein, the purified protein products were digested with Endo-H enzyme, and their specificity was analyzed by western blot mediated with ALV-J gp85-specific MAb (JE9). The results showed that both the protein band of 50 kDa and the protein band of 30 kDa could specially combine with gp85 MAb (JE9) (Fig. 3B, lane 1 and lane 2). These suggested that both the secretory recombinant protein in the cells' supernatant and the digested protein contained gp85 envelope protein of ALV-J.
Antibody responses induced by the secretory recombinant ALV-J gp85 protein combining with CpG-ODN adjuvant
ALV-J antibody titers were dramatically increased
The dynamical changes of serological antibody titers against ALV-J after vaccination are shown in Figure 4. Compared with those in the control group and in CpG group, the serum antibody titers in gp85 group and in gp85+CpG group were dramatically increased from 14 to 77 dpfi (p < 0.05), and booster inoculation could further improve the antibody titers. Meanwhile, the antibody titers in gp85+CpG group increased more rapidly and were higher than those in gp85 group from 21 to 77 dpfi.
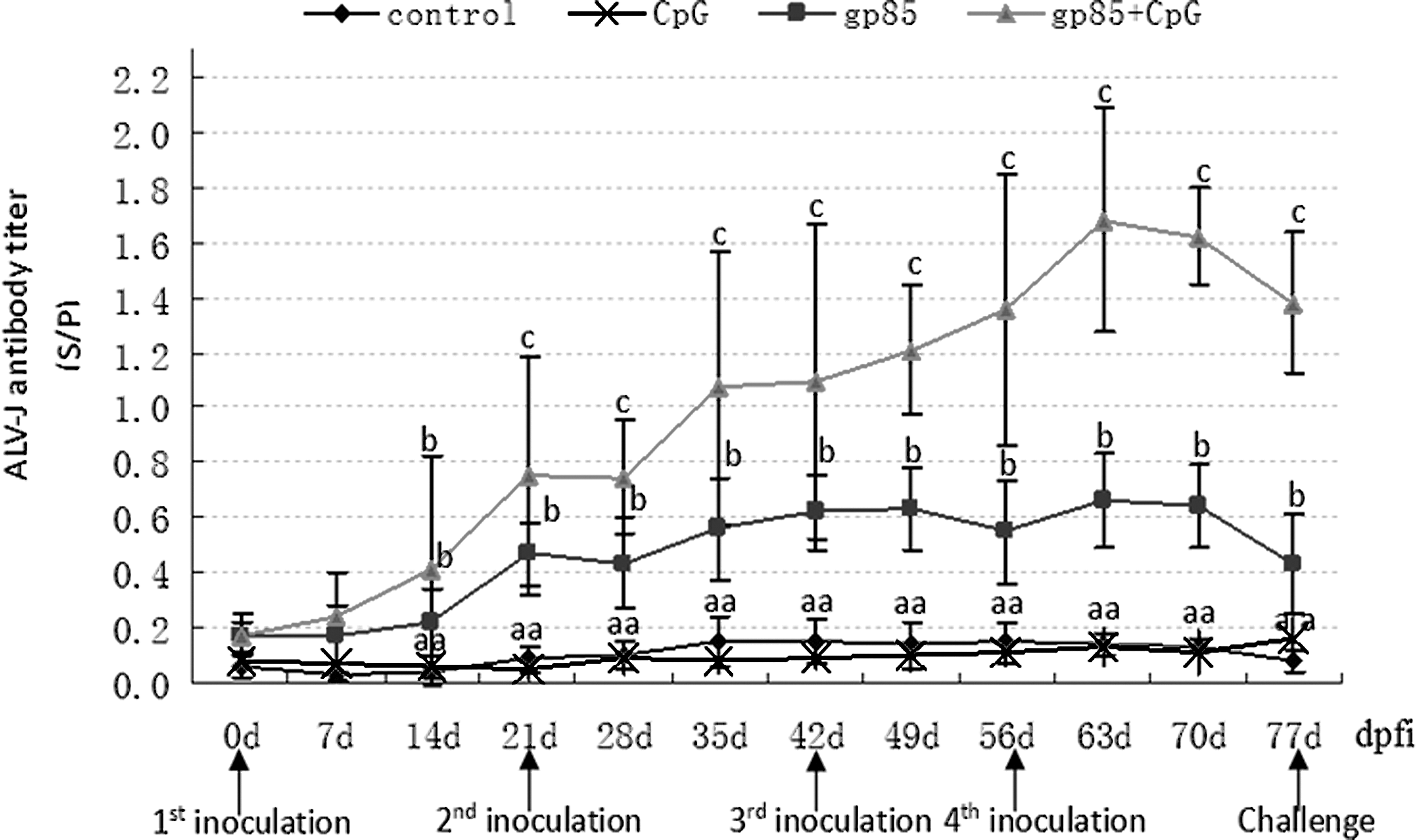
Dynamic change of ALV-J antibody in the chickens vaccinated with gp85 recombinant proteins expressed using Pichia pastoris combining with CpG-ODN adjuvant. dpfi, days post first inoculation; serum antibodies were detected with the IDEXX ELISA ALV-J Antibody Test Kits, and booster inoculations were performed at 21, 42, and 56 dpfi. The number of chickens in each group was 13. The bar with different letters means statistically significant difference (p < 0.05), and with same letters or no letters means no significant difference (p > 0.05) between groups.
Antibody-positive ratios were dramatically increased
The antibody-positive ratios of the vaccinated chickens are shown in Table 1. The antibody-positive ratios in gp85 group were increasing from 8% (1/13) on 28 dpfi to 69% (9/13) on 63 dpfi, and in gp85+CpG group were increasing from 23% (3/13) on 14 dpfi to 100% (13/13) on 49 dpfi and kept 100% until 77 dpfi.
The positive critical value (S/P ≥ 0.6) was obtained according to the ALV-J Antibody ELISA Kit (IDEXX).
ALV-J, J subgroup of avian leukosis virus; dpfi, days post first inoculation; S/P, sample to positive.
Cellular immune responses induced by the secretory recombinant ALV-J gp85 protein combining with CpG-ODN adjuvant
Spleen lymphocyte proliferation response was dramatically increased
To evaluate effects of the secretory recombinant protein on lymphocyte immune responses, the spleen lymphocyte proliferation responses in the inoculated chickens on 56 dpfi were detected and the results are shown in Figure 5A. Compared with those in the control group, the SI of lymphocyte proliferation responses to ConA or rgp85 protein in other three inoculated groups were dramatically increased (p < 0.05), and the SI in the gp85+CpG group was increased higher than those in gp85 group and in CpG group. These data suggested that the secretory recombinant gp85 protein plus CpG-ODN adjuvant could improve lymphocyte proliferation responses in the inoculated chickens.
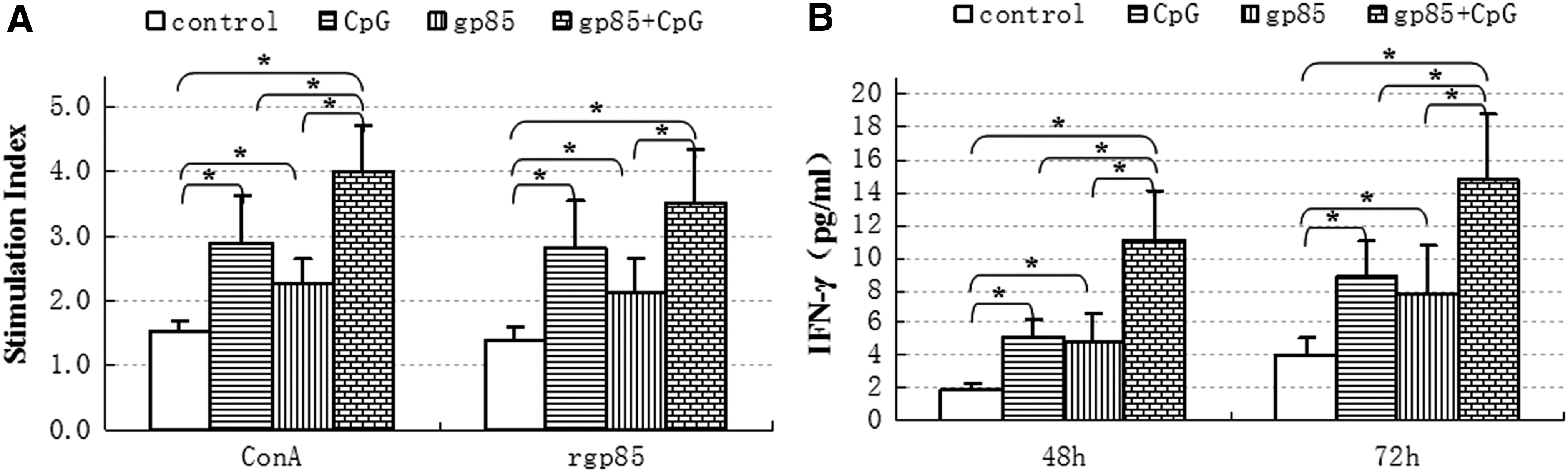
Spleen lymphocyte proliferation responses (
IFN-γ in spleen lymphocytes were dramatically increased
To analyze the effects of the secretory recombinant protein on cytokine responses, IFN-γ in the splenocytes of the inoculated chickens was detected 48 and 72 h post incubation and the results are shown in Figure 5B. Compared with those in the control group, IFN-γ in CpG group, in gp85 group, and in gp85+CpG group was dramatically increased (p < 0.05), and in gp85+CpG group was increased more than those in gp85 group and in CpG group (p < 0.05). These results suggested that the secretory recombinant gp85 protein plus CpG-ODN adjuvant could induce IFN-γ response in the inoculated chickens' splenocytes.
Immunoprotection of inoculation with the secretory gp85 protein plus CpG-ODN adjuvant against ALV-J challenge
Immunoprotection against the viremia caused by ALV-J challenge
Some studies showed that ALV-J could cause persistent viremia in most of the infected chickens (8,9) and the protection of inoculation with the secretory protein against viremia was evaluated in this study. The results in Table 2 shows that the viremia-positive ratio of the chickens was 61.5% (8/13) in CpG group, 46.2% (6/13) in gp85 group, and was 0% (0/13) in gp85+CpG group; they were significantly lower than 92.3% (12/13) in the control group. These suggested that the inoculation with gp85 protein plus CpG-ODN adjuvant could protect all of the chickens against ALV-J infection.
Different superscript letters on the data mean significant difference between groups p < 0.05; the same letters mean no significant difference between groups p > 0.05.
BW, body weight; NW, net weight; DI, development indices.
Immunoprotection against the immunosuppressive lesions caused by ALV-J challenge
ALV-J could cause growth retardation and immune organs' atrophy in the infected chickens (9,25) and so the immunoprotection of inoculation with the secretory protein against these lesions was tested in this study. The body weight and the DI of mainly immune organs in different groups on 3 weeks post challenge (wpc) are statistically analyzed in Table 2. The results shows that the body weight and the DI in the inoculated groups are higher than those in the control group, especially in gp85+CpG group these indices were also clearly higher than those in the CpG group (p < 0.05). These results suggested that the inoculation with the secretory gp85 protein plus CpG-ODN adjuvant could decrease the immunosuppressive lesions caused by ALV-J infection.
The neutralization of antisera in vitro
To clarify the effect of antisera induced by the secretory recombinant gp85 protein, the neutralizing test was performed using DF1 cells. DF1 cell is a selected chicken fibroblast cell line, where the exogenous ALVs, such as ALV-J or ALV-A, can replicate and grow, but the endogenous ALVs cannot (8,9). In this test, ALV-J antibody in the antisera obtained from the inoculated chickens was detected using the IDEXX ALV-J Antibody ELISA Kit, and the titers (S/P value) were 1.62 in the gp85+CpG group, 0.65 in gp85 group, and 0.11in control group, respectively. The growth curves of the neutralized ALV-J viruses in DF1 cells are described from 1 to 5 dpi in Figure 6. In virus wells with antisera in the control group, the viruses were detected in the DF1 cell supernatant from 3 to 5 dpi and the viral titers (S/P value) also increased with the increase of incubating days. In virus wells with antisera in gp85+CpG group, none of the three diluted titers could be detected for ALV-J virus on 5 dpi. However, in the virus wells with antisera in gp85 group, after neutralization using different dilutions, the viruses could be detected only on 5 dpi (Fig. 5, blue line). The results suggested that the antisera with higher ALV-J antibody titers could better neutralize ALV-J virus in vitro.
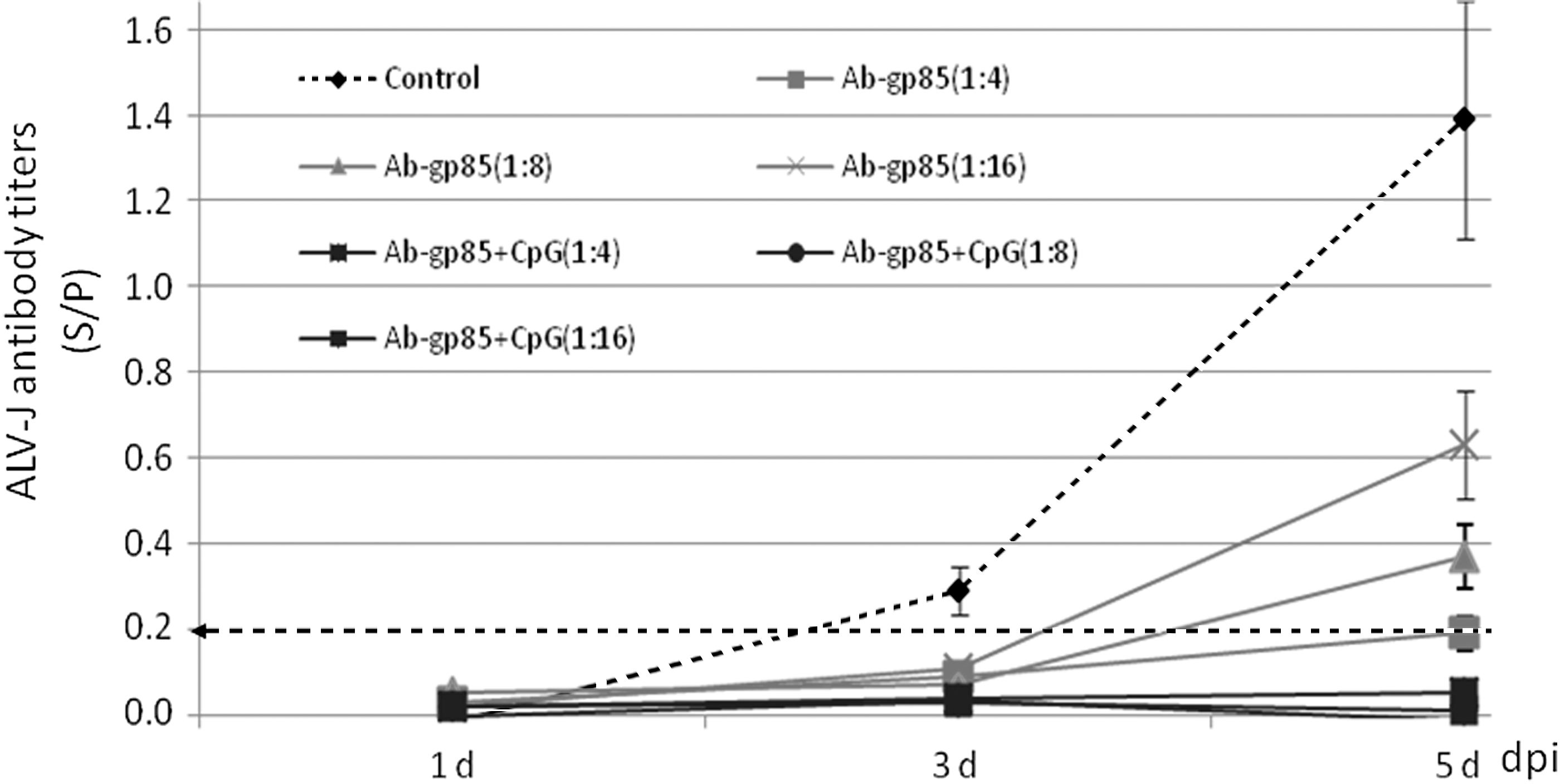
Neutralization of the antisera from different inoculated chickens against 102.625 TCID50 of ALV-J-NX0101 strain. the dashed line with arrow indicates the positive critical value determined using the IDEXX ALV P27 ELISA Test Kits; Ab-gp85+CpG (black line) indicates the antisera from some chickens in gp85+CpG group (Ab titers: S/P = 1.62); Ab-gp85 (light gray line) indicates the antisera from some chickens in gp85 group (Ab titers: S/P = 0.65); Control (dashed line) indicates the antisera from control chickens (Ab titers: S/P = 0.11). S/P, sample to positive.
IgG antibody in antisera could specially combine with ALV-J virus
To confirm that IgG antibody in antisera from the inoculated chickens could combine with ALV-J virus, IFA was performed in this study. The antisera from the gp85-inoculated chickens were used as the first antibody in this IFA and the FITC-labeled goat anti-chicken IgG antibody was used as the second antibody. The green fluorescent signals in the cytoplasm or nucleus of virus-infected cells could confirm that the antisera contained IgG neutralizing antibody. The results in Figure 7 show that the antisera from the chickens in gp85 group or in gp85+CpG group contained IgG neutralizing antibody; whereas the antisera in control group did not. On the basis of these data, the IgG antibody induced by the secretory gp85 protein expressed in P. pastoris could combine the growing ALV-J strain.
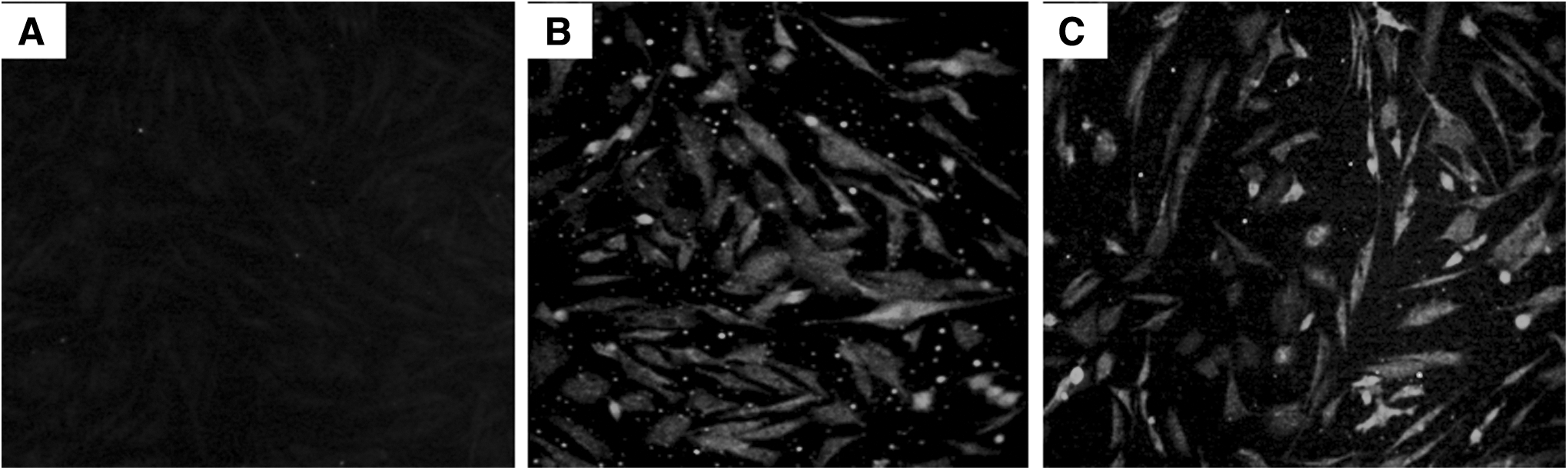
IFA mediated with the antisera from the inoculated chickens.
Discussions
The preparation of vaccine antigen is critical in the development and application of subunit vaccine against ALV infection. In our previous study, we reported that the subunit vaccine antigen of ALV-A/J prepared using E. coli combining with CpG-ODN adjuvant could induce the inoculated chickens to produce maternal antibody to protect their offspring against ALV-A/J early infection (7,32). But it is still difficult to prepare the subunit vaccine antigen of ALV-J using E. coli in its clinical application due to the complicated operation and unstable expression, etc. So it is important and necessary to find new methods to prepare sufficient protein antigens for further developing ALV-J subunit vaccine.
Some studies reported that the recombinant proteins expressed using P. pastoris could induce broadly neutralizing antibodies and marginal cell-mediated immune responses as vaccine antigens (3,27). Because the advantages of secretory expressions are that P. pastoris can secrete very little native proteins (11), thus the vast majority of the total protein in the medium was target proteins, the recombinant gp85 protein. This significantly simplifies the purification process and subsequent usage of the protein for vaccination purposes (22). In this study, the secretory recombinant expressed plasmid pPIC9-gp85, used for P. pastoris expression system, was designed. The gp85 gene of ALV-J was inserted into the specified sites of AOX1 promoter in pPIC9 vector and the recombinant vector pPIC9-gp85 was integrated into the genome of host yeast GS115 cells. After culture for five passages, the recombinant vector pPIC9-gp85 is rather stably expressed in GS115 cells under the induction of 0.5% methanol. In addition, the gp85 recombinant protein was also secreted into cultural medium of cells by the effects of its upstream α-factor secretion signal gene (21). The results showed that the recombinant gp85 protein reached about 30% of the total protein content in the supernatant, a sufficient purity as vaccine antigen.
To analyze the structure characters of the secretory recombinant gp85 protein, the expressed products were further concentrated, and dialyzed and purified, the recombinant protein reached above 95% of the total protein content (Fig. 3A, lane 2), and could specifically combine with MAb (JE9) against gp85 protein (Fig. 3B, lane 2). The expressed recombinant gp85 protein with ∼50 kDa of size could be digested into gp85 protein with ∼30 kDa of size with endo-β-N-acetylglucosaminidase H. These results confirmed that the secretory recombinant gp85 protein expressed in P. pastoris was modified by N-acetyl glycosylation. The compared result showed that the molecular weight of the N-acetyl-glycosylated gp85 protein was bigger than that of the recombinant gp85 protein expressed using E. coli cells (7) (data not shown). Obviously, due to the glycosylation, the secretory gp85 protein may have different molecular structure and bioactivity with the latter. So the immunogenicity or immunoprotection of the secretory gp85 protein needs further research.
To evaluate the immunoprotection of the secretory recombinant gp85 protein as subunit vaccine antigen, 7-day-old chickens were inoculated with the prepared gp85 protein combining with CpG-ODN adjuvant, the serological antibody responses and the cellular immune responses were detected, and the immunoprotection against viremia and the immunosuppressive lesions caused by ALV-J challenge was analyzed. The results showed that the antibody titers and the antibody-positive ratios were increased after inoculation, and the spleen lymphocyte proliferation responses to ConA or rgp85 protein were improved, and the induced IFN-γ from the splenocytes of inoculated chickens was clearly increased. These improved humeral and cellular immune responses suggested that the secretory recombinant gp85 protein had better immunogenicity than the recombinant gp85 protein prepared using E. coli cells in our previous study(7). More importantly, the viremia and the immunosuppressive lesions caused by ALV-J challenge in gp85+CpG group were dramatically decreased. These results suggested that the expressed secretory gp85 protein combining with CpG-ODN adjuvant could induce the inoculated chickens to produce better immunoprotection against ALV-J infection and the secretory gp85 protein was very potent to be used as vaccine antigen as like other recombinant protein expressed in Pichia pastoris (18,20).
In this study, the improved immunoprotective responses induced by the secretory recombinant gp85 protein plus CpG-ODN adjuvant may result not only from the prepared secretory gp85 protein from P. pastoris, but also from the immunoenhancement of CpG-ODN as adjuvant. Because CpG-ODN, one of toll-like receptor agonists, has been reported as an effective immune-enhancing adjuvant and it can activate B- and/or T-lymphocytes and improve the immunoprotective responses combined with vaccines in many studies (7,24). In this study, CpG-ODN could enhance the antibody responses and the cellular immune responses induced by the secretory gp85 protein; meanwhile, the inoculation of CpG-ODN alone could also partly protect the chickens against ALV-J challenge, and this finding deserves further attention.
Additionally, some subunit vaccines with many times of inoculations can better activate the memory lymphocytes and can further improve the antibody responses, which had been confirmed by the study (7). In this study, four times of inoculations were performed to improve the antibody responses to the secreted gp85 protein vaccine. The results showed that the secreted gp85 protein with three times of booster inoculations could induce the inoculated chickens to produce higher antibody responses (Fig. 4 and Table 1) and better immunoprotection against the viremia and immunosuppressive lesions caused by ALV-J challenge (Table 2) in deed. This can supply a scientific data for their application in clinics in the future.
To further clarify the immunoprotective mechanism of the inoculation with the secretory gp85 protein against ALV-J infection, the antisera with different ALV-J antibody diluters were collected to neutralize the virus in DF1 cells. The results suggested that the antisera with different ALV-J antibody diluters had different inhibition on the replication of ALV-J strain in cells. In gp85+CpG group, the antiserum (S/P = 1.62) with three diluted titer could completely neutralize 102.625 TCID50 of ALV-J strain; meanwhile, in gp85 group, the antiserum (S/P = 0.65) with three diluted titers could only neutralize part of the virus. The result of IFA further confirmed that IgG antibody in the antisera could directly combine with ALV-J virus in DF1 cells. These can partly clarify the difference of protection between gp85+CpG group and gp85 group. Of course, little has been known about the effects of other components, such as IFN-γ, in the splenocytes of the inoculated chickens on ALV-J virus, and the protective mechanism of the secretory gp85 protein combining with CpG-ODN adjuvant may need further research in the future.
In summary, this study successfully prepared the secretory recombinant ALV-J gp85 protein using P. pastoris and the N-acetyl-glycosylated gp85 protein as subunit vaccine antigen combining with CpG-ODN adjuvant, which could induce the inoculated chickens to produce sufficient immunoprotective responses to completely protect against ALV-J infection. This study first reported the immunoprotection of the secretory recombinant gp85 protein combining with CpG-ODN adjuvant against ALV-J infection in chickens.
Footnotes
Acknowledgments
This study was funded by the National Key Research and Development Program of China (No. 2016YFD0500800) and by the earmarked fund for the protection for the State Forest and Wild Animals (No. 2130211) and by Funds of Shandong “Double Tops” Program.
The authors would like to thank Prof. Zhao Xiaomin for providing the plasmids for P. pastoris expression.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.