
Abstract
The development of an effective preventative hepatitis C virus (HCV) vaccine will reside, in part, in its ability to elicit neutralizing antibodies (NAbs). We previously reported a genotype 1a HCV virus like particle (VLP) vaccine that produced HCV specific NAb and T cell responses that were substantially enhanced by Toll-like receptor 2 (TLR2) agonists. We have now produced a quadrivalent genotype 1a/1b/2a/3a HCV VLP vaccine and tested the ability of two TLR2 agonists, R4Pam2Cys and E8Pam2Cys, to stimulate the production of NAb. We now show that our vaccine with R4Pam2Cys or E8Pam2Cys produces strong antibody and NAb responses in vaccinated mice after just two doses. Total antibody titers were higher in mice inoculated with vaccine plus E8Pam2Cys compared to HCV VLPs alone. However, the TLR2 agonists did not result in stronger NAb responses compared to vaccine without adjuvant. Such a vaccine could provide a substantial addition to the overall goal to eliminate HCV.
Introduction
H
HCV virus-like particles (VLPs) offer an important approach for the development of a protective vaccine (8,13,24). We have previously reported a human liver cell-derived genotype 1a HCV VLP and characterized the biochemical and biophysical properties and morphology of the VLPs and importantly demonstrated the production of NAb and T cell responses to HCV (8,13). Similarly, a genotype 3a HCV VLP vaccine has also been shown to produce humoral and cellular immune responses (24). To further advance our vaccine approach to cover multiple HCV genotypes we produced a quadrivalent vaccine and established methods for large-scale vaccine production (12). The quadrivalent vaccine includes antigens from four common HCV genotypes: 1a/1b/2a/3a and is expected to produce broader cross-NAb responses and thereby be more effective than a single antigen.
We had previously shown that combining our genotype 1a HCV VLPs with the Toll-like receptor 2 (TLR2) lipopeptide agonists R4Pam2Cys or E8Pam2Cys as adjuvants resulted in strong antibody responses after just two doses of a monovalent HCV VLP vaccine (8). The TLR2 lipopeptides carry either a net positive (R4Pam2Cys) or negative (E8Pam2Cys) charge, binding to protein antigens by electrostatic interactions and improving their ability to produce antibody and T cell responses (9). Therefore, we wanted to determine whether the TLR2 agonists also worked well with our quadrivalent vaccine (12).
In this report we show that our quadrivalent vaccine produces strong antibody and NAb responses when combined with novel TLR2 agonists as adjuvants. Our results are encouraging for the development of a protective vaccine for HCV.
Materials and Methods
The construction of recombinant adeno-encoding HCV structural protein core E1 and E2 and the large-scale production, purification, concentration, and characterization of our quadrivalent HCV vaccine composed of genotypes; 1a, 1b, 2a, and 3a VLPs have been reported previously (8,12,13).
All animal experiments used 8- to 12-week-old BALB/c mice that were housed in the Doherty Resource Facility under specific pathogen-free conditions. Mice were obtained from the Animal House facility, Department of Microbiology and Immunology, The University of Melbourne. Animal experiments were performed according to local ethics committee approval. Groups of five age- and weight-matched BALB/c mice were immunized subcutaneously on each side of the base of tail with 80 μg (50 μL per dose) of quadrivalent VLP alone, quadrivalent VLP combined with the TLR2 agonists R4Pam2Cys or E8Pam2Cys (8), or PBS alone (control). The purified HCV VLPs were quantified for total protein (HCV core, E1 and E2) using a Bradford assay (12). Five nanomoles of each TLR2 lipopeptide adjuvant were used per dose of vaccine administered to each mouse. This equated to 9.9 μg for R4Pam2Cys and 16.2 μg for E8Pam2Cys per dose of vaccine. Two weeks later the mice received a second equivalent dose of quadrivalent vaccine. Animals were sacrificed 1 week after the final immunization, and blood was collected for serum preparation.
Sera were tested by ELISA to determine genotype specific antibody responses. Flat bottom 96-well polyvinyl plates were coated with purified HCV VLPs (20 μg/mL) in carbonate coating buffer (100 mM Na2CO3 and NaHCO3, pH 9.6) overnight at 4°C. The plates were then blocked with 100 μL of BSA (10 mg/mL) in PBS and incubated for 2 h at room temperature before washing four times with PBST (PBS containing v/v 0.05% Tween-20; Sigma Aldrich, Milwaukee). Serial dilutions of sera obtained from immunized mice were added to wells and incubated in a humidified atmosphere overnight. After washing, bound antibody was detected using horseradish peroxidase-conjugated rabbit anti-mouse IgG antibodies (Dako, Glostrup, Denmark). Following washing with 3,3′,5,5′-Tetramethylbenzidine (TMB), Liquid Substrate solution (Mabtech) was added to wells for 10–15 min. The reaction was stopped by addition of 50 μL of 0.16 M H2SO4. Absorbances were determined on a Labsystems Multiskan Multisoft plate reader at 450 nm. Titers of antibody are expressed as the reciprocal of the highest dilution of serum required to achieve an optical density of 0.2.
Neutralization assays using an HCV infectious cell culture system (HCVcc) using genotype 2a virus have been described previously (6,17). These were performed by mixing HCVcc virus with an equal amount of serially diluted immune serum. Each experiment was performed in triplicate. The virus/serum mixture was incubated for 1 h at 37°C before addition to Huh7.5 cells seeded 24 h earlier at 30,000 cells/well in 48-well plates for 4 h. Cells were washed at least four times and replenished with fresh DMF10NEAA and incubated for a further 48–72 h. Luciferase activity was measured in clarified lysates using Renilla luciferase substrate (Promega) and a FLUOstar OPTIMA microplate reader fitted with luminescence optics (BMG Life Technologies, Germany). The data shown are the mean from at least two independent experiments. Mab24 was used as a positive control for inhibition of HCVcc entry. This monoclonal antibody is a murine NAb that recognizes a linear epitope in the modified recombinant E2661 protein that lacks the HVR1, HVR2, and igVR regions (3).
Statistical analysis was performed using the PRISM 5.0 software (GraphPad). In all cases the mean ± standard deviation of the mean (SD) is shown unless otherwise stated. p values for statistical analysis were calculated using a one-way analysis of variance. Differences were considered statistically significant when p values were less than 0.05 (p < 0.05) with a 95% confidence level.
Results and Discussion
Mice in all vaccination groups developed genotype specific antibody responses (Fig. 1A: a–d). Mice inoculated with quadrivalent VLPs in E8Pam2Cys gave the highest genotype specific antibody titers. These were not significantly different than the titers observed with VLPs in R4Pam2Cys but were significantly higher than VLP alone (Table 1). The antibody response to the vaccine without adjuvant was still strong and comparable to vaccine supplemented with R4Pam2Cys. The strongest responses were detected against HCV genotypes 1a and 1b VLPs, while responses to genotype 2a and 3a VLPs were generally slightly lower across all groups. The geometric mean titer for anti-HCV VLP antibody was highest in mice receiving the quadrivalent HCV VLPs in E8Pam2Cys compared to HCV VLPs in R4Pam2Cys and HCV VLPs in PBS. These findings were consistent with our previous report showing strong immunogenicity of a monovalent genotype 1a vaccine combined with these TLR2 agonists (8).
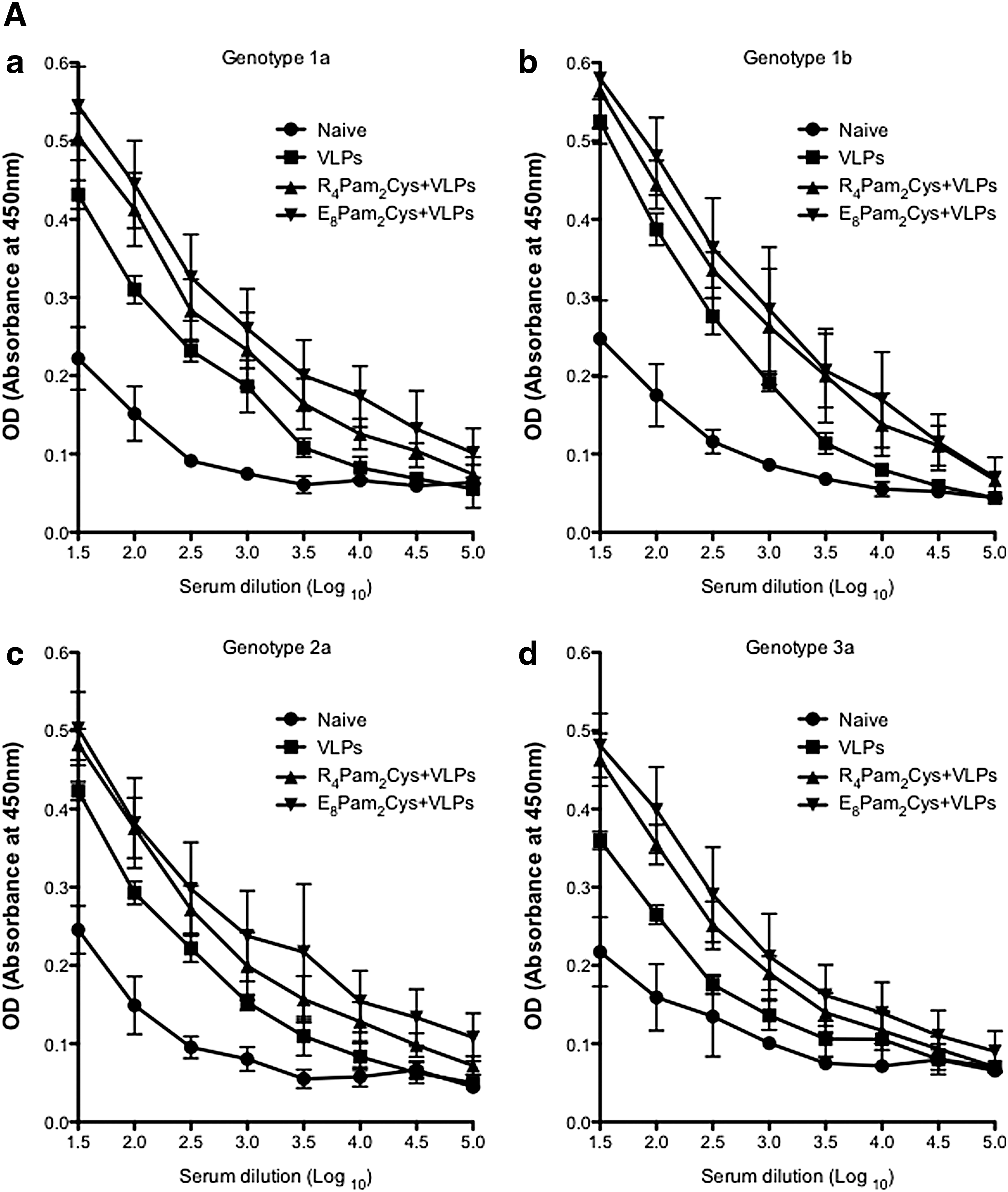
(A) Genotype-specific antibody responses elicited by immunization with quadrivalent vaccines. BALB/c mice (n = 5/group) were immunized subcutaneously at the base of the tail with PBS alone (control) or 80 μg of quadrivalent VLP at a ratio of 1:1:1:1 or quadrivalent VLP combined with 5 nmol of R4Pam2Cys or E8Pam2Cys in a final volume of 50 μL/dose. Two weeks later the mice received a second equivalent dose of quadrivalent vaccine. Genotype specific antibody in sera prepared from blood taken on day 21 was determined by ELISA using genotype 1a (
BALB/c mice (n = 5/group) were immunized subcutaneously at the base of the tail with PBS alone (control) or 80 μg of quadrivalent VLP at a ratio of 1:1:1:1 or quadrivalent VLP combined with 5 nmol of R4Pam2Cys or E8Pam2Cys in a final volume of 50 μL/dose. Two weeks later the mice received a second equivalent dose of quadrivalent vaccine. Genotype specific antibody in sera prepared from blood taken on day 21 was determined by ELISA using genotype 1a (A), 1b (B), 2 (C), or 3a (D) HCV VLPs as coating antigens. Values represent the geometric mean antibody titer (log10) and 95% confidence interval (shown in brackets) from 5 individual mice per treatment group using individual HCV genotypes; Gt1a, Gt1b, Gt2a, and Gt3a as coating antigens. In all cases, a significant difference (p < 0.05) was observed between treatment groups and naive mice. The geometric mean antibody titers were also significantly higher in mice immunized with VLPs plus E8Pam2Cys for all genotypes (p < 0.05).
HCV, hepatitis C virus; VLP, virus-like particle.
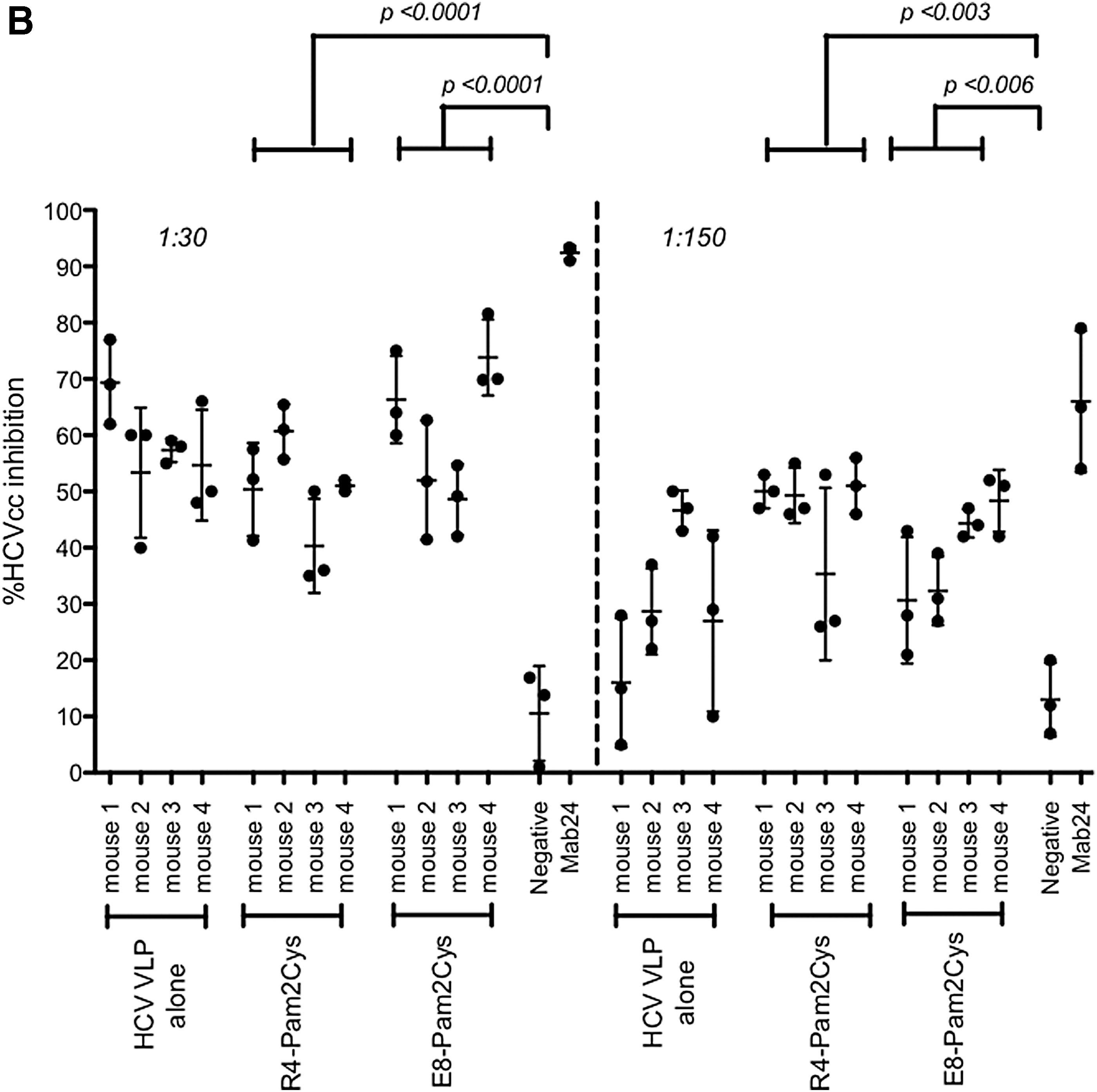
We then determined NAb responses to the vaccine using cell culture derived genotype 2a HCV as described in Ref. (37). We compared NAb responses in mice inoculated with quadrivalent vaccine combined with the TLR2 agonists R4Pam2Cys or E8Pam2Cys. Immune sera were tested at increasing dilutions (Fig. 1B). Sera (diluted 1:30) from mice inoculated with quadrivalent VLPs in R4Pam2Cys inhibited HCVcc entry by 50.6% (±SD 9.3%) compared to 60.2% (±SD 8.4%) with VLPs with E8Pam2Cys (p = 0.02), 10.6% (±SD 8.4%) (p < 0.0001) for negative control sera, and 92.4% (±SD 1.3%) with the monoclonal antibody Mab24 (Fig. 1B). Neutralization of Jc1 HCV was maintained at a serum dilution of 1:150. Sera from mice inoculated with quadrivalent VLPs in R4Pam2Cys inhibited HCVcc entry by 46.4% (±SD 9.9%) compared to 38.9% (±SD 9.1%) (p = 0.04) with VLPs with E8Pam2Cys, 13.0% (±SD 10%) (p = 0.005) for negative control sera, and 66.0% (±SD 12.5%) with the monoclonal antibody Mab24 (Fig. 1B). In contrast to the total antibody responses the NAb responses in mice vaccinated with HCV VLPs alone were not significantly lower than in mice vaccinated with adjuvanted HCV VLPs. These findings were consistent with our previous report showing strong immunogenicity of a monovalent genotype 1a vaccine combined with these TLR2 agonists (8).
There are estimated to be a large number of individuals who are unaware that they are infected with HCV, leaving a large residual pool of chronically infected individuals who will act as a persistent source for ongoing HCV transmission (35). An effective preventative vaccine could therefore have a significant impact on HCV prevalence. We have shown that our quadrivalent vaccine results in strong antibody and NAb responses when combined with the TLR2 agonists R4Pam2Cys or E8Pam2Cys. However, the vaccine was found to be strongly immunogenic after just two doses, even in the absence of adjuvant.
Although a vaccine derived from a single genotype can elicit NAb, the inclusion of antigens of a number of different genotypes could be expected to produce cross-NAb responses and thereby be more effective than a single antigen. NAbs act by recognizing native E1E2 heterodimers (16), and so an effective preventative vaccine should express native E1E2 structures. The significance of these regions on the viral envelope is further highlighted by the ability of passive immunization with polyclonal and monoclonal antibodies directed to these regions to protect human liver chimeric uPA/SCID mice against challenge with human serum derived HCV (27). In addition, immunization of macaques with a soluble cE2 subunit vaccine has also been shown to induce the production of broad cross NAb (25). These findings are highly significant for HCV vaccine design because they reinforce the need to present both E1 and E2 proteins in the correct structural organization as they would be on virions or HCV VLPs.
The E2 protein contains the major conformational neutralizing antigenic regions (antigenic domains A–E or the AR3 and AR4 regions) organized into distinct clusters of overlapping epitopes (16,20,21). Crystallographic structural analyses of the E2 core domain in complex with key NAbs have helped to define both a neutralizing and a non-neutralizing face to the E2 core antigenic surface (22,23). It has also been possible to map the binding of human monoclonal antibodies (HuMAbs) to these critical regions of E2 with the most potent antibodies binding to domains B and D of the protein (14). These studies reinforce the complex nature of how neutralizing epitopes are presented on the surface of HCV and suggest that these important antigenic regions need to be closely reproduced in a vaccine. A VLP based vaccine should be able to deliver these complex epitopes in the correct conformation.
Peptide epitope antigens can be made more immunogenic when covalently attached to Pam2Cys to target their delivery through TLR2 to dendritic cells (19). This results in the induction of robust antibody and cell-mediated immune responses (4,10,19,36). We have previously shown that R4Pam2Cys and E8Pam2Cys enhance the immunogenicity of genotype 1a HCV VLPs, with the improved overall antibody responses compared to VLPs in alum. This study shows that a similar enhancement in antibody responses can be achieved with a quadrivalent HCV VLP vaccine with the TLR2 agonist E8Pam2Cys, although this was not significantly greater than quadrivalent HCV VLP in R4Pam2Cys or quadrivalent HCV VLP vaccine alone. Studies of the ability of our quadrivalent HCV VLP vaccine to produce cross-NAb responses are currently ongoing using a large animal model.
HCV VLPs elicit both NAb and cellular immune responses (5,8,13,24). Furthermore, as HCV-specific NAbs recognize tertiary or quaternary structures (16), the repetitive and ordered particulate structure of HCV VLPs make them an attractive vaccine candidate (5,8,15). HCV VLPs are also able to present conformational epitopes in their native state. The development of cross-NAbs to epitopes on the surface of HCV that develop in the course of natural infection provides further encouragement for the development of a neutralizing HCV vaccine (26). An HCV VLP based vaccine would fulfill the requirement of delivering critical conformational neutralizing epitopes, and our report provides further data for the development of a preventative quadrivalent HCV VLP vaccine.
Footnotes
Acknowledgments
This work was supported by the National Health and Medical Research Council (NHMRC) of Australia, grant numbers 1060436 and 1126379. J.T. is supported by an NHMRC Practitioner Fellowship, number 106043.
This work was also supported by Australia-India Biotechnology Research Fund (BF040005), Department of Innovation and Industry, Australian Commonwealth Government.
We wish to thank Prof. Saumitra Das for having provided the cDNA for the production of the HCV genotype 3a VLP vaccine.
Author Disclosure Statement
No competing financial interests exist.