
Abstract
Zika virus (ZIKV) is an arbovirus belonging to the genus Flavivirus (Flaviviridae). ZIKV infection is associated with alterations in various organs, including the liver, lungs, and kidneys. Studies on the influence of posttranscriptional control on viral infections have demonstrated that microRNAs (miRNAs) interfere with different stages of the replicative cycle of several viruses and may influence the disease outcome. To shed light on ZIKV-induced regulation of host miRNA-processing machinery in the above organs, we analyzed the expression of genes encoding key proteins of the miRNA pathway in different ZIKV-infected continuous primate cell lineages (HepG2, A549, and MA104) by reverse-transcription quantitative polymerase chain reaction (RT-qPCR). Expression of the genes encoding the miRNA-related proteins DGCR8, Ago1, and Ago3 in HepG2 cells and Drosha, Dicer, Ago2, and Ago3 in A549 and MA104 cells was significantly altered in the presence of ZIKV. Our results suggest that ZIKV modulates miRNA levels during infection in liver, lung, and kidney cells, which may be an additional mechanism of host cell subversion in these organs.
Introduction
R
The majority of ZIKV infections are asymptomatic; however, when symptomatic, the condition is clinically characterized by a mild and self-limiting febrile disease accompanied by headache, pruritic maculopapular exanthema, nonpurulent conjunctival hyperemia, arthralgia, joint edema, and myalgia, with spontaneous disappearance of the symptoms after 3–7 days (4,13,15,30,38). However, by the end of 2015, Brazil confirmed the first deaths caused by ZIKV and the association of ZIKV infection with severe neurological disorders besides other complications involving organs such as the liver, lungs, and kidneys—where significant histopathological changes were observed in fatal adult cases (2).
microRNAs (miRNAs or miRs) are small, noncoding RNA sequences that range in size from 19 to 25 nucleotides and act as posttranscriptional gene regulators by binding to messenger RNAs (mRNAs). Through complete or incomplete base pairing, miRNAs regulate the gene product mainly in a negative manner by inhibiting the translation process via binding to the 3′ untranslated region (3′UTR) of the mRNA or leading to its degradation (3,14,31). Studies on the influence of posttranscriptional control on viral infections have demonstrated that miRNAs interfere with different stages of the replication cycle of several viruses. For instance, studies on dengue virus serotype 2 (DENV-2) showed that overexpression of miR-133a decreased the number of virus-infected cells and the number of viral RNA copies produced, although the mechanism by which miR-133a altered viral replication was not clarified (7). A study on microglial cells infected with Japanese encephalitis virus (JEV) showed a decrease in miR-34c-5p expression, which reduced proinflammatory cytokine production—interestingly, miR-34c-5p was highly expressed in microglial cells infected by West Nile virus (WNV), suggesting that the modulation pattern of certain miRNAs may be linked to specific viral pathogenesis (22). Insertion of multiple miRNA target sequences into a neurotropic flavivirus genome followed by infection of mice brains led to inhibition of the pathogenicity of this virus in vivo (33).
A recent study on ZIKV replication in human astrocytes demonstrated that the virus induces global downregulation of miRNAs with only a small subset being upregulated, including miR-30e-3p, miR-30e-5p, and miR-17-5p, concomitantly to deregulation of numerous processes known to be involved in flavivirus infection, including genes involved in the unfolded protein response pathway (21). Regarding the insect vector, 17 host miRNAs were found to be modulated by ZIKV infection in Aedes aegypti mosquitoes—many of them already reported to have their expression altered by DENV and WNV infection in mosquitoes (28). Besides, several flaviviruses—including ZIKV—are known to produce miRNA-like small RNAs and thereby regulate host gene expression (36).
In this study, we evaluated the mRNA expression profiles of key genes in the miRNA-silencing pathway (Drosha, DGCR8, Dicer, and Argonaute family members) in HepG2, A549, and MA104 cells experimentally infected with ZIKV to shed light on virus-induced regulation of host miRNA-processing machinery in the liver, lungs, and kidneys, respectively.
Materials and Methods
Viral sample
The viral isolate BeH818308 was provided by the Section of Arbovirology and Hemorrhagic Fevers of the Evandro Chagas Institute. The virus was obtained from a fatal case of a patient from the State of Maranhão (Brazil) infected with ZIKV and was isolated from several organs in culture of Aedes albopictus cells (clone C6/36) (2,12).
The viral stock was prepared in Vero cells (kidney epithelial cells from the African green monkey Chlorocebus aethiops) after three passages.
Maintenance of cell cultures
HepG2 cells originated from a human hepatocellular carcinoma and are widely used to study the in vitro effects of flavivirus infection. For example, studies on the components of the apoptosis pathway, the production of cytokines, and the role of miRNAs in viral infections have been performed using HepG2 cells (24,26,34). A549 cells are derived from a human lung carcinoma and have been used in proteomic analysis during DENV infection and for the analysis of interferon (IFN) production during viral infections, among others (8,17,25). MA104 cells are healthy adherent epithelial fetal kidney cells from African green monkey and are widely used in rotavirus research (35). These cell types were carefully selected after Azevedo et al. (2) had observed alterations in several tissues by histopathology of biological materials obtained from a patient from the State of Maranhão (Brazil) who died due to ZIKV disease. Intense histopathological alterations, such as necrosis, intense inflammation, and hemorrhages, were observed in the liver, lungs, and kidneys of the patient (2).
Cell culture was performed using Dulbecco's modified Eagle's medium (DMEM) supplemented with 5% fetal bovine serum (FBS) and maintained at 37°C with 5% CO2, as described elsewhere (34).
Viral infection
Infection of the cells was performed in six-well plates when the cell monolayers exhibited more than 70% confluency. At the time of infection, the medium was discarded, and the cells were washed twice with phosphate-buffered saline (PBS). Infection was performed using a multiplicity of infection of 0.5 viral genome copy/cell and the cells were incubated for 1 h at 37°C with 5% CO2 for viral adsorption. Two wells were used as negative controls (mock) and contained only culture medium without the virus. After viral adsorption, the culture medium was supplemented with 2% FBS. The plate was incubated at 37°C with 5% CO2 for 4 days, and the cells of one well were collected every 24 h for extraction of viral RNA and cellular mRNAs.
RNA extraction
Both viral RNA and cellular mRNAs were extracted from the samples of infected cells using the PureLink Viral RNA/DNA Mini Kit (Life Technologies) following the manufacturer's protocol.
Quantification of viral RNA
Viral RNA was quantified based on an external curve generated with concentrations determined by serial dilution using the probe and primers described by Lanciotti et al. (23); the probe and primers bind to the E protein coding region with concentrations of 200 and 300 nM, respectively. Quantification was performed on the qPCR 7500 Fast Real-Time PCR System (Promega) using the TaqMan One-Step RT-PCR Master Mix Reagents Kit (Life Technologies) according to the manufacturer's protocol.
Determination of viral titer
Viral titer was determined by plaque assay in Vero cells grown in 24-well plates. On confluency, cells were washed with PBS and incubated with 10-fold serial dilutions of supernatants from HepG2, A549, or MA104 cells infected with ZIKV for 1, 2, 3, or 4 days. After 1 h at 37°C, the medium containing nonadsorbed virus particles was replaced by a semisolid medium (1.6% carboxymethylcellulose in DMEM) supplemented with 2% FBS and cells were incubated for another 48 h at 37°C. Subsequently, cells were simultaneously fixed and stained with a solution composed of 0.1% crystal violet, 30% ethanol, and 20% formalin in PBS to allow for plaque counting.
Detection of target gene mRNA expression
The mRNA expression levels of the selected genes were quantified with the SuperScript® III Platinum® SYBR® Green One-Step qRT-PCR Kit using the 2−ΔΔCT method with the qPCR 7500 Fast Real-Time PCR System (Promega) according to the manufacturer's instruction. The primers used in the mRNA screening of the proteins involved in the miRNA pathway are listed in Table 1. The cellular β-actin gene was used as an endogenous control to ensure the RNA integrity of the extracted samples.
F, forward; R, reverse.
Statistical analyses
The statistical analyses and the graphs were generated on the Prism 6 software (GraphPad). Statistical analyses were performed by one-way ANOVA using the Dunnett's multiple comparison test. Significance was considered when p ≤ 0.05. Biological triplicates were performed at each stage of this study and all results presented as mean ± range.
Results
Viral load and titer in ZIKV-infected cells
The viral RNA quantification showed variable concentrations depending on the cell type and collection time as follows: quantification of ZIKV RNA copies in HepG2 cells during the 4 days of infection showed higher viral production at 2 days postinfection (dpi), the highest copy number of ZIKV RNA in A549 cells occurred at 4 dpi, and the highest copy number of ZIKV RNA in MA104 cells during the 4 days of infection was observed at 3 dpi (Fig. 1A). Regarding viral titer, the peak was observed at 2 dpi for all cell types (Fig. 1B).
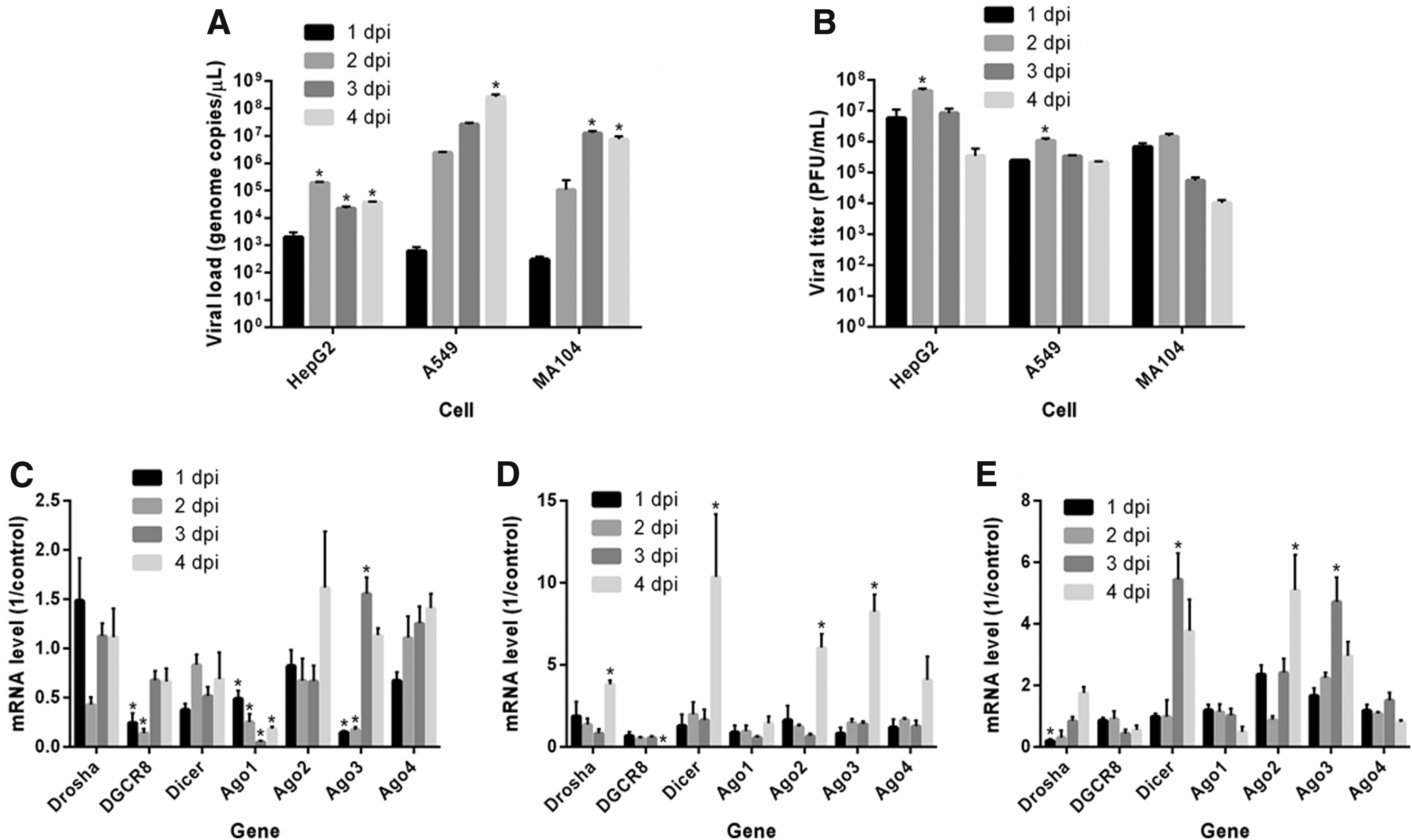
Quantification of viral load and titer and analysis of miRNA-related gene expression in ZIKV-infected cells. HepG2, A549, and MA104 cells were infected with ZIKV and harvested every 24 h for up to 4 days to quantify viral genome by RT-qPCR
Expression of miRNA-related genes in ZIKV-infected cells
The expression profiles of mRNAs encoding key miRNA pathway proteins (Drosha, DGCR8, Dicer, and Ago1–4) in cells infected with ZIKV were determined by RT-qPCR.
In HepG2 cells, the variation in the Drosha, Dicer, Ago2, and Ago4 mRNA expression levels did not significantly differ during the 4 days of ZIKV infection. Conversely, the DGCR8, Ago1, and Ago3 mRNA expression levels differed over the 4 days of infection, with the highest expression levels observed at 3 and 4 dpi for DGCR8, 1 dpi for Ago1, and 3 dpi for Ago3. Compared with the uninfected cells (mock), DGCR8 expression was decreased at 1 and 2 dpi. A decrease in Ago1 expression was observed during the 4 days of infection, whereas Ago3 expression decreased at 1 and 2 dpi and increased at 3 dpi (Fig. 1C).
In A549 cells, Ago1 and Ago4 mRNA expression levels did not significantly differ over the 4 days of ZIKV infection. However, significant differences were found compared with other mRNAs, with DGCR8 showing lower expression level, while Drosha, Dicer, Ago2, and Ago3 showed higher expression levels at 4 dpi. When the expression levels were compared between the infected and uninfected cells, an increase in Drosha, Dicer, Ago2, and Ago3 expression was observed at 4 dpi, with no DGCR8 expression observed at this time point (Fig. 1D).
In MA104 cells, the gene expression profiles of the miRNA pathway proteins showed no significant differences for DGCR8, Ago1, and Ago4 during the 4 days of ZIKV infection, whereas Drosha, Dicer, Ago2, and Ago3 mRNA expression differed significantly. The highest Drosha and Ago2 expression levels were observed at 4 dpi, whereas the highest Dicer and Ago3 expression levels were observed at 3 dpi. Compared with the control, low Drosha expression was observed at 1 dpi, an increase in Dicer and Ago3 expression was observed at 3 dpi, and an increase in Ago2 expression was observed at 4 dpi (Fig. 1E).
Discussion
ZIKV infection in HepG2 cells led to a decrease in DGCR8, Ago1, and Ago3 expression compared with the control cells. A study with yellow fever virus (YFV)-infected HepG2 cells conducted by Holanda et al. (16) showed that the virus modulated the expression of almost all mRNAs of the proteins of the miRNA pathway (Drosha, DGCR8, Dicer, Ago2, Ago2, and Ago4) except for the Ago3 mRNA, suggesting that viral infection played an important role in the regulation of proteins that contributed to viral replication and modulated the immune response of infected cells. Studies on hepatitis B virus (HBV) infection of HepG2 cells found that more than 30 miRNAs related to innate immunity via Toll-like receptors were downregulated during viral infection (20). In addition, the pattern of upregulation and downregulation of miRNAs was modified according to the level of cellular damage (39). A study on DGCR8 expression showed a significant increase in the mRNA levels of proteins involved in the miRNA pathway in HepG2 cells compared with healthy controls (19). Our findings suggest that ZIKV infection significantly interferes with DGCR8 mRNA expression in cells, which can lead to a decrease in Drosha-DGCR8 microprocessor assembly since DGCR8 is an important Drosha cofactor.
During DENV-4 infection, the Drosha, Dicer, and DGCR8 proteins were downregulated in A549 cells, where they were assumed to play important roles in viral replication and viral pathogenesis (6). However, the present study observed that the Drosha, Dicer, Ago2, and Ago3 mRNA levels were upregulated in this cell lineage during ZIKV infection at 4 dpi, which coincided with the increase in the viral load at 3 and 4 dpi. A computational prediction study described several miRNAs in the DENV genome that would play roles in viral replication and pathogenesis (27). WNV was also found to modulate the expression of an miRNA derived from a highly conserved sequence located at the end of the 3′-UTR in the viral genome, shown to be essential for WNV replication in C6/36 cells (18).
Few studies on the miRNA profiles in MA104 cells have been conducted to date. Our study showed that ZIKV infection modulated the expression of four mRNAs of the miRNA pathway (Drosha, Dicer, Ago2, and Ago3); these alterations overlapped with the peak detection of viral RNA. In a study with MA104 cells infected with rotavirus, which is a segmented double-stranded RNA virus, most of the miRNAs studied were downregulated during infection (41). In other studies, viral infection was shown to downregulate the expression of miRNAs and to induce a consequent decrease in IFN-I, which is an important protein component of the antiviral immune response to HBV, thereby contributing to the progression of HBV replication (37,40). Furthermore, infection by WNV was shown to induce the expression of multiple miRNAs, including the Hs_154 miRNA that inhibits antiapoptotic proteins and is therefore critical for the apoptotic process of infected cells and for viral pathogenesis (32). However, more studies are still needed to properly associate the miRNA-related proteins with the apoptotic process in the cells.
Regarding the insect vector, recent studies were contradictory when addressing gene expression changes in response to ZIKV infection in A. aegypti mosquitoes, not allowing to determine yet whether the expression of miRNA-related genes such as Dicer and Ago1–4 is altered or not on viral infection in insect cells (1,11).
It is difficult to discern a priori whether upregulation of a specific miRNA represents part of the host innate immune response and therefore might inhibit viral replication, is specifically induced by the virus and therefore might enhance virus replication or is simply irrelevant (9). As previously shown, in some contexts, components of the animal RNA interference (RNAi) pathway do not play an antiviral role—on the contrary, miRNAs and the RNAi apparatus have been co-opted to negatively regulate toxic effectors of the protein-based IFN response (29).
In this study, differential expression profiles of key genes of the miRNA-silencing pathway were found among three distinct primate cell types experimentally infected with ZIKV, suggesting that viral infection is able to affect miRNA biogenesis in liver, lung, and kidney cells. The alterations in the expression of miRNA-related proteins coincided with the peak expression of viral RNA in all cells tested, which may suggest a modulator effect of ZIKV that favors viral replication and control of immune responses. Viral modulation of the miRNA-silencing pathway may play a critical role in mediating viral pathogenesis and therefore may represent novel targets for therapeutic interventions.
Footnotes
Acknowledgments
We thank the Postgraduate Program in Biology of Infectious and Parasitic Agents (BAIP/UFPA) and the Evandro Chagas Institute (IEC) for their support to this study. This work was supported by Pro-Rectory of Research and Postgraduate Studies (PROPESP) of Federal University of Pará (UFPA), and by grants from the National Council for Scientific and Technological Development (CNPq–439971/2016-0; 303999/2016-0; 440405/2016-5) and Coordination of Improvement of Higher Level Personnel (CAPES–Zika Fast-Track). Those who provided funding had no role in study design, data collection and analysis, decision to publish, or preparation of the article.
Author Disclosure Statement
No competing financial interests exist.