
Abstract
CD137 is a costimulatory molecule transiently expressed on activated T cells after mitogen or antigen stimulation that can be exploited for isolating antigen-specific T cells as reported in mouse models. By utilizing an antiporcine CD137 monoclonal antibody (mAb, clone 3B9) developed in our laboratory, we isolated virus-specific CD8β T cells from peripheral blood of pigs experimentally infected with different porcine reproductive and respiratory syndrome virus (PRRSV) strains. Similar to mouse, porcine CD8β T cells also express CD137 transiently upon Concavalin A stimulation while the unstimulated cells did not. Most frequently, virus-specific CD8β T cells were isolated at low levels from peripheral blood of pigs experimentally infected with PRRSV strains VR2385, NADC20, and MN184B at 49 and 63 days postinfection. The results suggest that porcine CD137-specific mAb is a useful tool for isolating virus-specific CD8 T cells from peripheral blood and tissues of pigs after in vitro stimulation with viral antigen.
Introduction
CD137
Porcine reproductive and respiratory syndrome is currently endemic worldwide, causing significant economic losses to the global pig industry (3,6). In addition to neutralizing antibody responses, the cell-mediated immunity including CD8-specific cytolytic T cell (CTL) responses are necessary for effective porcine reproductive and respiratory syndrome virus (PRRSV) protective immunity (4). In pigs, SLA class I-restricted CTLs can be identified by CD8α and CD8β heterodimers on the surface of cells (2). In addition to the SLA class I-restricted CD8 T cells, CD8α was also found to be expressed in a subpopulation of CD4 T-helper cells, natural killer cells, and T cell receptor (TCR)-γδ T cells (2). Owing to the exclusive expression of CD8β in SLA class I-restricted CD8 T cells, these cells can be identified in the peripheral blood of pigs by specific staining with an anti-CD8β antibody. The specific detection of PRRSV-specific CD8 T cells in peripheral blood cell populations is the first step that will aid in their isolation, amplification, and further characterization (1). In this study, we developed a mAb (3B9) specific to porcine CD137 and characterized it as a tool for purifying virus-specific CD8 T cells in the peripheral blood of pigs experimentally infected with genetically divergent strains of PRRSV.
Materials and Methods
mAb (Cl. 3B9) specific to porcine CD137 was generated by mouse hybridoma technology as previously described (8). The mAb 3B9 specificity was confirmed by immunofluorescence assay of mammalian cell lines (PK-15 and 293T cells) over-expressing porcine CD137 protein. In brief, the cells were transfected with constructs encoding the CD137 gene using lipofectamine LTX reagent (Thermo Fisher Scientific). Two days post-transfection, cells were fixed with 4% paraformaldehyde and permeabilized with 0.1% Triton X-100 if necessary and stained with mAb clone 3B9 (1:350 dilution) for overnight at 4°C. After extensive washings with phosphate buffered saline (PBS)-Tween20 (0.1%), cells were stained with Alexafluor 488 or 594-conjugated antimouse IgG (1:300 dilution; Thermo Fisher Scientific) for 45 min at 37°C. After extensive washings with PBS-Tween20, images were acquired with a fluorescent microscope. Cells transfected with empty vector served as negative control.
For the animal study, the experimental procedures in the study protocol were thoroughly reviewed and approved by Virginia Tech Institutional Animal Care and Use Committee (IACUC), protocol number 13-110-CVM. The pigs were handled humanely throughout the study and treated with appropriate care according to the guidelines set forth by American Veterinary Medical Association (AVMA). A total of 31, 4-week-old, PRRSV-negative pigs were divided into four treatment groups (n = 7 or 8 in each group). Pigs in each group were inoculated either with Dulbecco's modified Eagle's medium or with each of the PRRSV strains VR2385 (lineage 5), NADC20 (lineage 9), or MN184B (lineage 1; 1 × 105.2 tissue culture infectivity dose50 [TCID50]/2 mL each pig). All pigs received booster virus inoculation of the same dose at 42 and 56 days after the primary infection. Serum and heparinized blood samples were collected weekly for analyzing antibody, viremia, and peripheral blood mononuclear cells (PBMCs) isolation, respectively. PBMCs were isolated by density gradient centrifugation; aliquots were made at a concentration of 2 × 107 cells/mL and subsequently stored in liquid nitrogen until further analysis.
CD8β T cells were enriched by magnet-activated cell sorting (MACS). After thawing and overnight resting, PBMCs were stained with 0.1 μg CD8β antibody (PG164A; Washington State University Antibody Service Center) per million cells in staining buffer (cold PBS, 0.5% bovine serum albumin, and 2 mM EDTA) for 30 min on ice. Stained cells were positively selected after incubation with antimouse IgG microbeads (Miltenyi Biotec) and passing through LD magnetic columns (Miltenyi Biotec). Cells bound to column were collected as CD8β POS fraction and the cells passed through the column were collected as CD8β NEG fraction.
Autologous monocyte-derived dendritic cells (MoDCs) were prepared by incubating plate-adherent monocytes (derived from PBMCs prepared at 42 and 56 days postinfection [DPI]) with porcine IFN-α1 (20 ng/mL; IBI Scientific) and porcine GM-CSF (20 ng/mL; Thermo Fisher Scientific) and loaded with ultraviolet [UV]-irradiated PRRSV-infected pulmonary alveolar macrophage for 5 h.
CD8β POS T cells were either left unstimulated or stimulated with antigen-loaded MoDCs at a ratio of 10:1 for 18–20 h. As a positive control, they were stimulated with Concavalin A (10 μg/mL) in the presence of MoDCs for 24 h. CD8β POS T cells were stained with CD8β mAb, PG164A (0.1 μg/million cells), and biotin-conjugated CD137 mAb 3B9 (0.1 μg/million cells) for 30 min on ice and subsequently stained with PE-conjugated antimouse IgG2a and PerCP/5.5-conjugated streptavidin. By fluorescence-activated cell sorting (FACS) analysis, CD8β POSCD137POS T cells were sorted out of 100,000 counts of CD8β POS T cells per sample. Dead cells were excluded from the isolation by DAPI staining.
Results and Discussion
A specific mAb clone 3B9 was found to specifically stain the full-length CD137 variant 2 as well as the CD137 extracellular domain as detected by immunofluorescence analyses (Fig. 1A). The 3B9 mAb did not stain the cells transfected with empty vector control plasmid (Fig. 1A). Since the CD137 variant 2 (lacking a domain in the extracellular region of the CD137) is the shortest transmembrane form (13), the 3B9 mAb is expected to bind both CD137 variants 1 and 2 on porcine T cells.
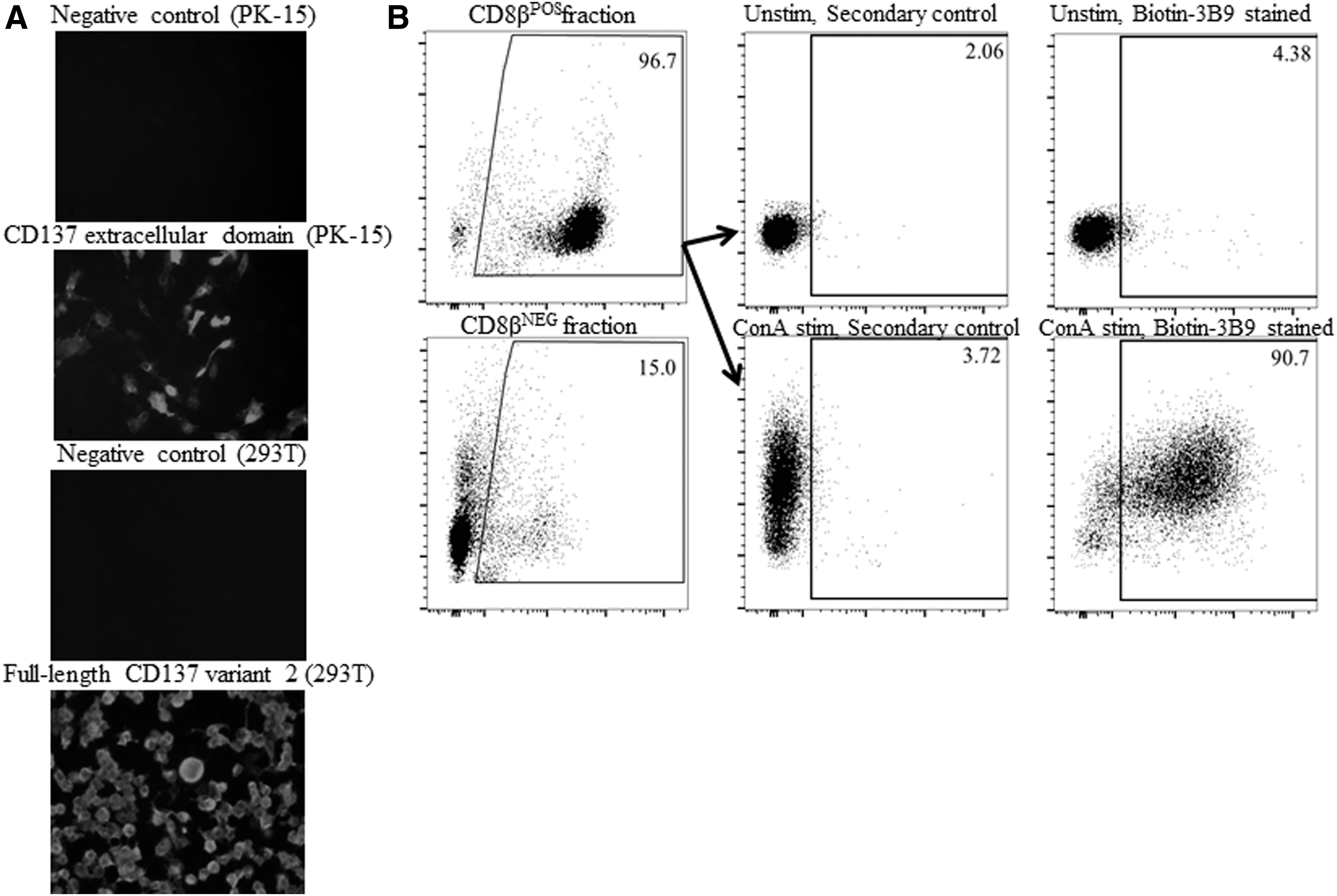
Characterization of a mouse mAb specific to porcine CD137.
When the mouse T cells were stimulated by Concavalin A or anti-TCR mAbs, they expressed CD137 in an inducible manner while the unstimulated resting T cells failed to upregulate CD137 (9). We examined whether porcine T cells express CD137 in the presence or absence of mitogen stimulation. CD8β T cells were enriched from PBMCs by MACS and the purity of the enriched CD8β T cells was found to be at least 95% as examined by flow cytometry analysis (Fig. 1B, left panels). When CD8β T cells were stimulated with Concavalin A in the presence of MoDCs, they strongly upregulated CD137 on more than 90% cells while the unstimulated resting T cells showed background staining for CD137 (Fig. 1B, right panels). Consistent with mouse CD137 expression on CD8 T cells (9), our result suggests that porcine CD8 T cells also upregulated CD137 upon stimulation with mitogen, and possibly with antigen through TCR stimulation, while the resting unstimulated T cells did not express CD137. Therefore, the highly specific staining of porcine CD137 mAb 3B9 could possibly be employed for purifying the antigen-activated CD8 T cells specific to PRRSV.
To isolate the CD8β T cells specific to PRRSV, pigs were experimentally infected with three different genetically diverse strains of PRRSV, VR2385 (lineage 5), NADC20 (lineage 9), and MN184B (lineage 1) (7). All pigs were found to be seroconverted by 14 DPI (Fig. 2A). Consistent with this, pigs infected with all three strains reached peak viremia titers between 7 and 14 DPI (Fig. 2B). Pigs in control group did not show seroconversion until the end of the study (data not shown). Moreover, at 21 DPI, the viremic titers observed in pigs were significantly different between treatment groups (two-way analysis of variance [ANOVA], Tukey's multiple comparisons test, p ≤ 0.05) (Fig. 2B). Nevertheless, the viremia in pigs of all three treatment groups reduced to barely detectable levels by 28 DPI.
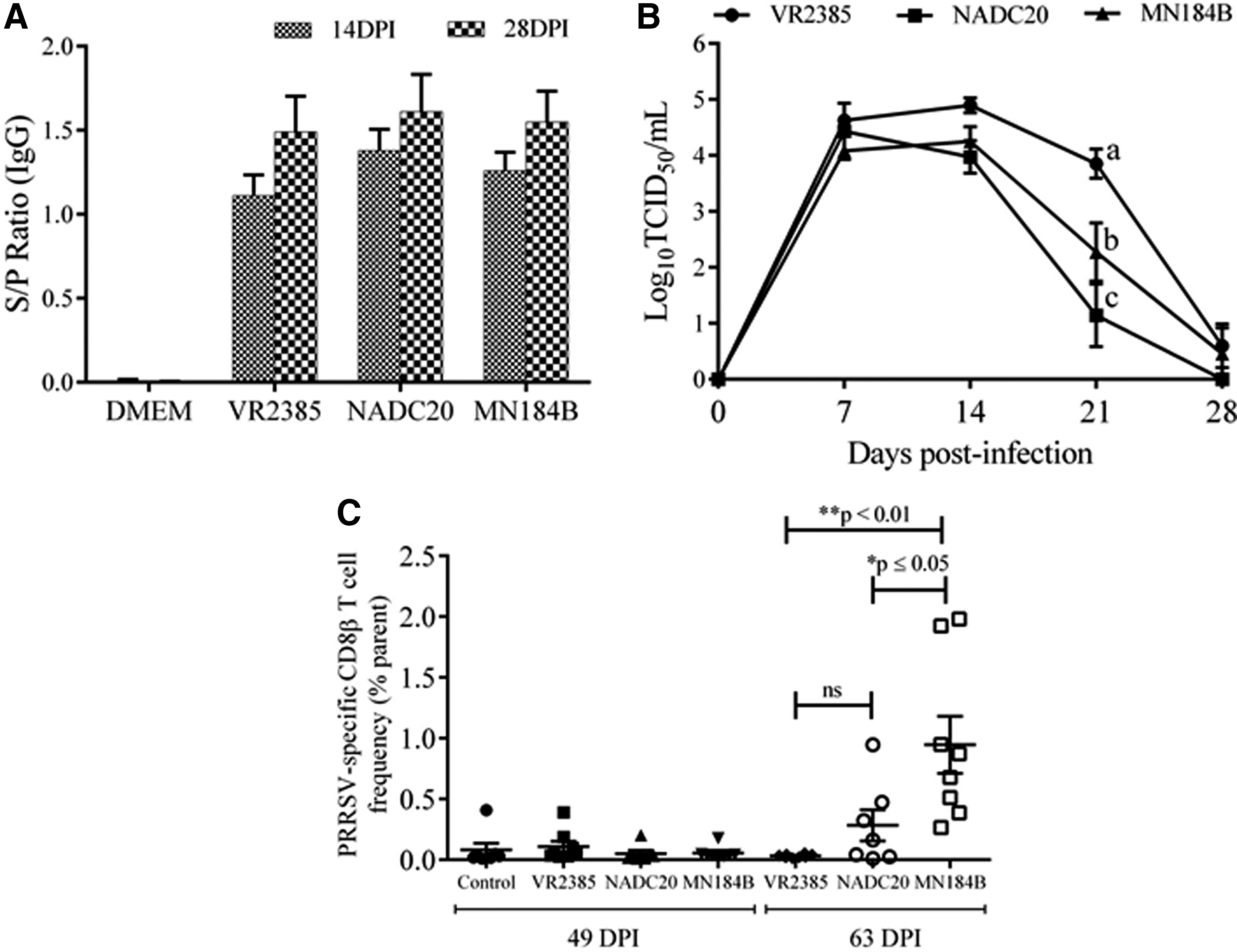
Viremic status, seroconversion in pigs experimentally infected with PRRSV strains and the frequencies of antigen-specific CD8β T cells isolated from infected pigs.
Total CD8β T cells were enriched from PBMCs prepared from peripheral blood of PRRSV-infected pigs at 49 and 63 DPI. These time points were selected for isolating CD8β T cells since the previous study showed that the PRRSV-specific T cell frequencies were found to be gradually increasing in peripheral blood of pigs after the viremic period (5). The CD8β T cells were enriched by MACS and in vitro stimulated with PRRSV antigen-loaded DCs. The virus strain-specific CD8β T cells were isolated by porcine CD137 staining followed by FACS. As shown in Figure 2C, the frequencies of PRRSV-specific CD8β T cells are <0.5% of parental population in most pigs at 49 DPI for all three virus strains examined. At 63 DPI, PRRSV-specific CD8β T cells were rarely isolated (<0.05%) from peripheral blood population of pigs infected with VR2385 (Fig. 2C).
Our results are consistent with a previous report suggesting that IFN-γ-specific T cell responses are weak in the peripheral blood of pigs during first 13 weeks post-PRRSV infection, which was predominantly contributed by CD4posCD8α pos T populations (5). The CD4neg CD8α/β pos T cell populations were rarely found in PRRSV-infected pigs during persistent phase of PRRSV infection (5). However, pigs infected with NADC20 and MN184B strain showed higher number of virus-specific CD8β T cells in a proportion of pigs at 63 DPI (Fig. 2C). Particularly, the pigs infected with MN184B strain showed significantly higher frequencies of CD8β T cells than those infected with VR2385 strain (one-way ANOVA, Tukey's multiple comparison test, p ≤ 0.05) (Fig. 2C). In conclusion, PRRSV-specific CD8β T cells can be isolated by CD137-specific staining and by FACS. The frequencies of PRRSV-specific CD8β T cells isolated by CD137 staining are generally at low levels in pigs infected with different PRRSV strains. Isolation of virus-specific CD8β T cells from peripheral blood of PRRSV-infected pigs by CD137 staining is the first crucial step that will enable us to establish SLA class I-restricted CD8 T cell clones specific to a particular PRRSV strain to characterize T cell epitopes in vitro. In general, the porcine CD137 mAb may be a useful tool for isolation and characterization of virus-specific T cells in pigs to improve our understanding of T cell immune responses against virus pathogens including PRRSV.
Footnotes
Acknowledgments
We thank Ms. Melissa Makris for her technical assistance in flow cytometry data acquisition and analyses. We also thank Karen Hall, Sarah Kenley, Laura Thomas, Julie Holdridge, Kimberly Allen, and Catherine Caldwell for their timely support in the pig study. This study was funded by a grant from USDA National Institute of Food and Agriculture (AFRI grant No. 2013-01131).
Author Disclosure Statement
No competing financial interests exist.