
Abstract
Over the past decade, metabolic dysfunction has been re-examined as an area of interest in a variety of atopic, malignant, and autoimmune conditions. The unique immune and, specifically, lymphocyte metabolic dysfunctions in disease are starting to be more clearly classified. Exploring the lymphocyte metabolic profiles of these diseases in the current literature, we characterize these diseases into two distinct metabolic groups. The influence of viral infection on immune metabolic dysfunction is explored. During an acute or chronic infection, there is a varied rapid shift in nutrients available to immune cells. This article explores the effect of these changes in nutrient availability and the resulting outcome on immune cell function. From this, a new hypothesis is proposed that these two groups of conditions, characterized by two unique immune profiles, are caused by either acute or chronic viral infections. The physiology behind this is explored, which proposes a new step between a healthy immune system and that of disease, in a unique working model.
Introduction
T
In their resting state, lymphocytes maintain their modest energy demand with oxidative phosphorylation. This is the metabolic process in which cells use enzymes to oxidize nutrients, thereby releasing energy that is used to produce adenosine triphosphate. On activation, lymphocytes meet the increased energy demands by switching to predominantly aerobic glycolysis. Aerobic glycolysis is the transformation of glucose to lactate in the presence of oxygen. Glycolysis is less energy efficient, but it creates energy at a much quicker rate. Failure to increase glucose consumption by lymphocytes reduces cell proliferation. However, after activation, a later significant drop of the glucose level within the extracellular space forces the activated T cells to enter a more dormant state of activity. This is to spare energy as a survival mechanism to counteract the glucose insufficiency (27). Conversely at times of high glucose availability, studies have shown over-activated proinflammatory T lymphocytes (12). It is clear that controlling lymphocyte nutrient availability can have a dramatic effect on the host's immune response and function.
Distinct T cell subsets display unique metabolic characteristics with regard to their energy demands at rest and on activation. Regulatory T cells (Tregs) are a subpopulation of cells that modulate the immune system, have a role in suppressing activated cells, and maintain a resting state once an infection is cleared. Memory T cells, and also Tregs, are known to have smaller metabolic demands, similar to the profile of resting T cells, and rely predominantly on oxidative phosphorylation. T effector cells have a much higher glucose demand, and increased glucose availability promotes lymphocyte differentiation to this subtype of cells. T helper (effector) cells, Th17 and Th1, which have been implicated in a number of autoimmune and malignant conditions, have a high glycolytic capacity and can efficiently increase their glycolytic rates on addition of glucose (7). It has been found that different metabolic environments can control T lymphocyte cell fate and differentiation (4).
Lymphocyte Metabolism and Disease: A Brief Overview
In the tumor microenvironment, lymphocytes have been found to display similar characteristics in many different types of cancer. Warburg first described how cancer cells uniquely produce their energy predominantly through aerobic glycolysis even in the presence of oxygen, which is known as the Warburg effect (22). Because of the Warburg effect, and thus the high glucose consumption by cancer cells, the tumor microenvironment has been found to have a reduced extracellular concentration of glucose (8). As a consequence of this, tumor-infiltrating activated T lymphocytes are forced to decrease aerobic glycolysis rates and, therefore, lose their useful effector functions (10). Owing to the relative low glucose availability for immune cells, they have dysfunctional activation due to utilization of other energy sources for activation. This allows for prolonged activation and the cells are thought to enter a more state of dormant “anergy,” and consequently this prevents a useful response against the malignant cells (27). This proinflammatory state is maintained by the deregulation of Tregs due to the shift of balance between regulatory and effector lymphocytes, itself supported by the metabolic shift (3).
Immune cells in autoimmune and atopic conditions have been found to have similar metabolic changes. Many conditions also have T cells that display upregulated aerobic glycolysis and remain in a state of prolonged activation. However, there are stark differences between different autoimmune conditions. Lymphocytes in rheumatoid arthritis (RA) show an “anti-Warburg” effect, wherein they have upregulated aerobic glycolysis but have a low glucose demand similar to the metabolic picture of dysfunctional lymphocytes seen in cancer (25). Once activated in lupus, T lymphocytes, have a dysfunctional state of increased oxidative phosphorylation to maintain their energy demands and do not upregulate aerobic glycolysis (21). Although thyroid hormones increase the aerobic glycolysis rates of lymphocytes in Graves' disease initially, the maximal level of the mediators of aerobic glycolysis has been found to be reduced (23), again suggesting a similar picture to that of RA.
As we have reviewed, at times of hyperglycemia, T lymphocytes are activated with an associated proinflammatory state, as seen in type 1 diabetes (6). In a similar manner, in multiple sclerosis (MS) patients, it has been shown that CD4+ T cells, the main subset of lymphocytes involved in MS pathogenesis, have downregulated oxidative phosphorylation and greatly increased rates of aerobic glycolysis and glucose demand (5). A similar picture is also seen in inflammatory bowel disease patients, wherein local T cells have a significantly increased glucose uptake (7). Asthma T lymphocytes have also been found to have upregulated aerobic glycolysis with high glycolytic activity, which promotes inappropriate lymphocyte activation (15). The authors of the study found that blocking aerobic glycolysis inhibited T cell activation and the development of asthma.
Building the Hypothesis
The link between infection, inflammation, autoimmune disease, and cancer has been widely explored and the inflammatory hypothesis is now widely accepted. However, the step between a healthy immune system and that of inflammation and disease has yet to be fully established. Authors exploring the metabolic changes to lymphocytes that occur in autoimmune and malignant disease suggest that there must be a rapid metabolic shift, triggering the lymphocyte energy utilization changes seen in disease. Conditions causing a gradual shift in metabolism, such as obesity, do not trigger these rapid changes, or cause these diseases directly. In an otherwise healthy individual, the only condition that sets off a rapid ongoing change in local metabolism is infection, which often causes a rapid glucose and metabolic shift. To support this hypothesis, we first need to review the effects viral infection has on immune cell metabolism, subsequently a striking pattern emerges.
In acute infection, many viruses induce high rates of aerobic glycolysis in infected cells, which induces a rapid intracellular glucose flux to support viral spread (20,26). In chronic viral infections, ongoing infection and latent infection can continue to induce aerobic glycolysis in infected cells; this has been seen in investigations of cells in hepatitis C infection among others (17,18). This maintains a high glucose demand in infected cells long after the initial infection. HIV infection has been found to target glucose metabolism directly in CD4+ T cells. The resulting changes in the metabolic status of host immune cells are thought to contribute to the systemic metabolic changes and inflammation seen in HIV-infected individuals (16).
It has been widely discussed how viral infections may trigger inflammatory, autoimmune, and malignant conditions, but the pathway that explains how this may occur has remained elusive. Reviewing the patterns seen in disease and infection, I can propose that there are three distinct lymphocyte outcomes after a viral infection, which further classifies lymphocyte metabolic changes and thus these diseases into two distinct groups (Fig. 1). First, after an acute viral infection, ideally the infection is cleared, and the host's immune cells are deactivated by Tregs. Once glucose levels normalize and the lymphocytes are deactivated, the immune system then returns to a normal resting state.
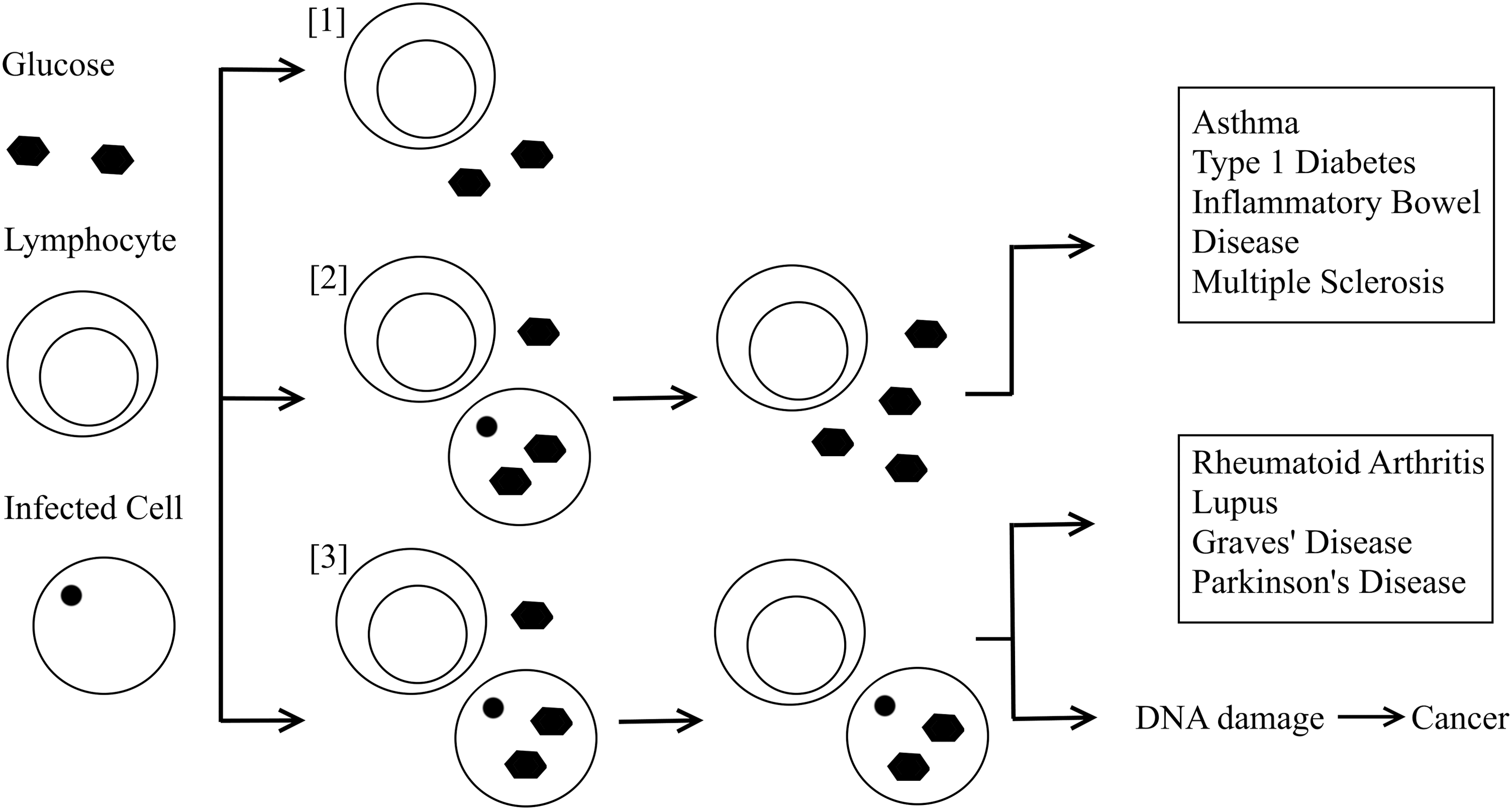
The proposed pathway of the pathological series of events during viral infection. [1] Both lymphocytes and infected host cells utilize glucose, the infected cells are cleared, and the immune system returns to a normal resting state. [2] The infected cells are cleared after persisting acute infection. This causes a shift of glucose available in the extracellular space. This causes lymphocytes to enter a state of hyperactivation. [3] During chronic viral infection, local lymphocytes are initially activated. Owing to the persisting infection, there is reduced glucose available to lymphocytes, so they are forced to utilize other energy sources with dysfunctional activation.
The second outcome I propose is that in an acute viral infection, the infection is cleared after persisting. After this, there is another glucose flux once the virus is cleared, as the previously infected cells now have a reduced glucose uptake, resulting in a sudden increase of glucose available to lymphocytes to metabolize in the extracellular space. This increases glucose availability to local lymphocytes, therefore, they enter a hyperactive state with a high rate of aerobic glycolysis, as we have already discussed. This pattern is seen in many diseases such as MS, asthma, inflammatory bowel disease, and type 1 diabetes. This may be why, with asthma and some other autoimmune conditions, it has been proposed that individuals are at increased risk of disease when they have not previously been exposed to microbial antigens. Consequently, their immune systems mount a more prolonged immune response and cannot quickly clear the infection with a memory T cell response (11). Also this could be why there is an increased risk of these conditions in patients with diabetes, and thus an increased baseline glucose level. We also know conditions of hyperglycemia promote reductions in Tregs and, therefore, promote the ongoing hyperactive T lymphocyte proinflammatory state (12,19). These phenomena are also reported to promote reductions in Treg populations, which prevent clearing of the proinflammatory state once glucose levels normalize (1). This would suggest that these conditions are triggered by an acute viral infection, which has been widely suggested in the literature.
Third, in a local state of chronic viral infection, there is ongoing low glucose availability to the activated lymphocytes, due to the increased glucose demand of infected cells, and those with latent infection. This leads the immune cells to try to gain energy through other means, as seen in RA, lupus, and Graves' disease, to utilize other energy sources, which maintains them in an active dysfunctional state against the infected cells. This model would suggest that these autoimmune conditions are caused by chronic viral infections. It is worth noting that they have all been linked with the Epstein Barr virus as a potential causative factor (14), which itself is known to upregulate glycolysis in infected cells (24). This type of lymphocyte metabolism is also seen in chronic infections associated with cancer (9). It is likely that DNA damage, and this associated immune dysfunction from chronic infection, triggers the shift from inflammation to malignancy.
Neurodegenerative conditions could also be accumulated into this hypothesis. Similar to lupus patient lymphocyte metabolism, in Parkinson's disease, lymphocytes also display increased oxygen uptake and reduced aerobic glycolysis. In Alzheimer's disease, lymphocytes also show metabolic derangement; however, the evidence is not yet sufficient to further contemplate this (2).
Factors that normalize lymphocyte metabolism and improve healthy immune cell invasion, such as a further acute infection and exercise, have been found to improve outcomes in all of these diseases. By reviewing the current literature base exploring metabolic changes in the immune system in disease and health, we can clearly see a pattern emerging. Linking these pieces together, I have proposed a new step in the pathway between a healthy immune response and that of disease, which encompasses a wide variety of diseases of unknown etiology.
Discussion
The current literature base, along with the extensive work on cell metabolism over the past century, supports this hypothesis, which starts to put together the pieces of the inflammatory pathway of disease. There is an obvious lack of data in a number of conditions that would further strengthen this understanding. Only a relatively small number of atopic and autoimmune conditions along with viral infections have been discussed in this article. Lymphocyte metabolism has not been specifically investigated in a number of autoimmune and atopic conditions, which would further help to develop this hypothesis. A number of mediators in this pathway are already being targeted, particularly in the field of oncology, and this is a promising area for future progress across a number of diseases and specialties.
Footnotes
Author Disclosure Statement
No competing financial interests exist.