
Abstract
Microparticles (MPs) are vesicles that are released by budding from plasma membrane of living cells. Recently, the role of MPs in antiviral activity has been proposed. We investigated quantity and anti-influenza activity of MPs from human alveolar epithelial cells A549, human bronchial epithelial cells BEAS-2B, human colon adenocarcinoma cells HT-29, and the human lung fibroblast cells MRC-5. MPs were found from all four cell lines. However, anti-influenza activity against an H1N1 influenza virus was found only from MPs of A549 and BEAS-2B. BEAS-2B cell differentiation did not increase MP release. Methyl-β-cyclodextrin (MβCD) increased MP release and anti-influenza activity in HT-29 and A549. MP release increased after calcium ionophore A23187 treatment in three cell lines but only in HT-29 after forskolin treatment. These findings provide in vitro data supporting the role of MPs as an innate defense against influenza virus and as an approach to enhance the defense.
Introduction
C
MPs have been shown to play a role in intercellular exchange of biological signals and information (19). Another role of MPs in antiviral activity has been proposed. It has been reported that the vesicles secreted by human tracheobronchial epithelial cells have a neutralizing effect on human influenza A virus (IAV) (14), and our recent data have shown anti-influenza activity of MPs isolated from human BAL (29). In the initial step of infection, IAV hemagglutinin (HA) protein binds to sialic acid residues on the respiratory epithelial cell, triggering endocytosis of the virion into the host cell. Since MPs express surface molecules specific to their parental cells, it is possible that MPs from the respiratory epithelial cells contain IAV receptors that can act as a decoy receptor to prevent the binding of viral HA on cell surface. However, data of MPs released from cell lines are limited.
Generation and release of MPs occur during different biological processes, including stimulation with high shear stress as present in arteries with a severe stenosis, cellular differentiation, senescence, apoptotic cell breakdown, or exposed to proinflammatory, prothrombotic, or proapoptotic substances (19). It is generally assumed that MPs form when the asymmetrical distribution of lipids between the inner and outer leaflets of a plasma membrane is lost (6).
In resting cells, phosphatidylserine (PS) is located almost exclusively in the inner leaflet of the plasma membrane (30). The transbilayer lipid redistribution is under the control of three translocase proteins: flippase, an inward-directed PS-specific pump; floppase, an outward-directed PS-specific pump; and a lipid scramblase, promoting unspecific bidirectional redistribution across the bilayer (34). A significant increase of cytosolic Ca2+ accompanying cell stimulation may lead to the collapse of the membrane asymmetry by activation of calcium-dependent enzymes such as scramblase and floppase (21). Increase in cytosolic Ca2+ also promotes disassembly of the cytoskeleton through protein-degrading enzymes, for example, calcium-activated calpains (13).
In vitro release of MPs can be induced by various compounds, for instance, calcium ionophore A23187, a mobile ion carrier that is used for increasing intracellular Ca2+ levels (8); and forskolin, a herbal extract commonly used as a specific activator of the cyclic AMP (cAMP) (11,33). High intracellular levels of cAMP can lead to increased intracellular Ca2+ levels (16,17). Moreover, cyclodextrins (CDs), nonreducing cyclic glucose oligosaccharides, are used to selectively extract membrane cholesterol, the major lipid constituent of the plasma membrane of mammalian cells (20). The disturbances of lipid rafts by removing cholesterol from cell membranes by methyl-β-cyclodextrin (MβCD), a derivative of CDs, have been suggested to induce MP release (25). However, data of MP induction by calcium ionophore, MβCD, A23187, and forskolin in other cells are still limited.
In this study, we hypothesized that MPs may relate to mucosal defense against viral infection, since the mucosa is a membrane that consists of layers of epithelial cells. Thus, we selected three epithelial cell lines, including human lung alveolar epithelial cell line A549, transformed bronchial epithelial cell line BEAS-2B, human colon adenocarcinoma cells line HT-29, and nonepithelial cells, the human lung fibroblast cell line MRC-5 to determine the differences in MP release and anti-influenza activity. We also challenged the cells with various concentrations of MβCD, calcium ionophore A23187, and forskolin to assess their ability to induce MP release.
Materials and Methods
Cell lines
Human lung alveolar epithelial cell line A549 (lung adenocarcinoma, ATCC CCL-185) was cultured in Dulbecco's modified Eagle's medium (DMEM; Sigma) supplemented with heat-inactivated 10% fetal bovine serum (FBS; Gibco); transformed bronchial epithelial cell line BEAS-2B (human bronchial epithelium, ATCC CRL-9609) was cultured in bronchial epithelial growth medium (BEGM; Lonza); human colon adenocarcinoma cell line HT-29 (colon adenocarcinoma, ATCC HTB-38) was cultured in McCoy's 5A media (Gibco) supplemented with 10% heat-inactivated FBS; and the human lung fibroblast cell line MRC-5 was cultured in minimum essential medium (MEM; Gibco) supplemented with 2 mM sodium pyruvate, 0.1 mM nonessential amino acids, and heat-inactivated 10% FBS.
Viral strains
The influenza viruses used in this study were the 2009 pandemic H1N1 (pdmH1N1) influenza virus A/Thailand/MVCU-013/2009 propagated in Madin–Darby canine kidney (MDCK) cells and a reverse genetic virus containing the monobasic cleavage site, HA gene from a highly pathogenic avian influenza A/Thailand/1(KAN-1)/2004 (H5N1) virus, and seven genes from A/Puerto Rico/8/34 (H1N1) virus (rVac-H5) (28).
Air–Liquid interface culture
Cells were cultured on 12 mm polyester Transwell inserts with a pore size of 0.4 μm (Corning). BEAS-2B cells were plated at 100,000 cells per insert in growth factor-supplemented medium (BEGM) (Lonza Walkersville, Inc.). When confluent (∼3 days), medium from apical and basal chambers was removed and differentiation medium (BEDM) was added to the basal chamber only. Medium was replaced every 48 h. Differentiated cell was tested after an incubation period of 21 days (27).
Lectin staining
Cells were trypsinized and cell suspensions were then blocked for nonspecific binding with 3% bovine serum albumin (Sigma) in 1× phosphate-buffered saline (PBS) for 30 min. After discarding blocking solution, the cell suspensions were incubated with 10 μg of FITC-conjugated Maackia amurensis lectin MAA I or Sambucus nigra lectin SNA (Vector Laboratories) in blocking solution for 30 min at room temperature, then washed three times with PBS, and finally, stained cells were fixed with 1% paraformaldehyde (PFA) in 1× PBS. Flow cytometry measurements were performed immediately using an FACSCalibur instrument and CellQuest software (Becton Dickinson).
MP collection
Cells were maintained at 37°C in 5% CO2 before being seeded onto 12-well plates at a density of 5 × 105 cells per well. After overnight incubation, culture supernatant was removed and 1 mL of fresh media with or without various concentrations of testing compound, including MβCD (Acros Organics), calcium ionophore A23187 (Sigma), and forskolin (Sigma), were added before incubating at 37°C in 5% CO2. Culture supernatant was collected and cell debris was removed by centrifugation two times at 200 g for 5 min. MPs were isolated from culture supernatant by centrifugation at 20,000 g for 120 min at 4°C.
Apoptotic body staining
For apoptotic bodies using as positive control, HT-29 cells were treated with hydrogen peroxide in serum-free medium for 24 h. The cell culture supernatants were collected and centrifuged two times at 200 g for 10 min to remove cells and cell debris. MP samples and supernatant from the HT-29 cells were stained with 3 μL of annexin V-FITC in annexin V buffer in the dark at room temperature for 15 min and subsequently stained with 1 μg/mL of propidium iodide (PI) in the dark at room temperature for 15 min. The stained samples were analyzed immediately using an FACSCalibur instrument and CellQuest software (Becton Dickinson).
Flow cytometry and MP analysis
According to the property of MPs that express PS on their surface, we stained MPs with annexin V, a specific marker for PS. Isolated MPs resuspended in 100 μL of culture medium were stained with 3 μL annexin V-FITC (BD Pharmingen) in a tube that has 20,000 particles of 1.34 μM latex beads (SPHERO™ Fluorescent Particles), diluted with twofold annexin V-FITC binding buffer (BD Pharmingen), and incubated for 15 min in the dark at room temperature. Flow cytometry measurements were performed immediately using an FACSCalibur instrument and CellQuest software (Becton Dickinson). MPs were defined as those smaller than the latex bead and positively stained for annexin V. Absolute MP count per microliter was calculated by using the formula (number of MPs counted/number of beads counted) χ (total number of beads in tube/volume of tested MPs).
Hemagglutination inhibition assay
MP samples were twofold serially diluted in 1 × PBS. Each diluted MP was mixed with four hemagglutination U/25 μL of influenza virus A/Thailand/MVCU-13/2009 (H1N1) or rVac-H5 virus in a 96-well U-shaped microtiter plate. The virus–MP mixture was incubated at room temperature for 30 min. Then, 50 μL of 0.5% goose red blood cells was added to the virus–MP mixture and was incubated at 4°C for 30 min. HI titers were read as reciprocal of the highest dilution with complete inhibited hemagglutination. Each MP sample was tested in duplicate.
Microneutralization assay
MP samples were twofold serially diluted in MEM plus 1 mM TPCK-treated trypsin, and then, 50 μL of each MP dilution was mixed with an equal volume of the test virus at a concentration of 25 TCID50/50 μL. The virus–serum mixture was incubated at 37°C for an hour. A one hundred microliter of the mixture was inoculated onto an MDCK cell in a 96-well tissue culture plate. After an overnight incubation at 37°C, the tissue culture plate was washed and fixed with 80% cold acetone in 1× PBS. The assays were run in duplicate. The reaction plate was tested by ELISA to determine the amount of influenza nucleoprotein produced in the infected MDCK cells using an antiviral nucleoprotein monoclonal antibody (Milipore) as the primary antibody, a peroxidase-conjugated antibody (Dako) as secondary antibody, and TMB (KPL, Inc., Gaithersburg, MD) as the chromogenic substrate. The reaction plate was read under a spectrophotometer at the wavelengths of 450 and 630 nm. The neutralizing (NT) titer was defined as the reciprocal of the highest MP dilution that reduces more than 50% of the amount of viral nucleoprotein in the reaction wells compared with the virus control wells.
Statistical analysis
Data are presented as means and SEMs. Groups were compared using the Student t test and one-way ANOVA with Tukey's multiple comparisons test. p < 0.05 was considered significant.
Results
MPs from A549, BEAS-2B, HT-29, and MRC-5 cells and their anti-influenza activity
We investigated the difference in MPs released from epithelial cells among alveolar epithelial cells A549, bronchial epithelial cells BEAS-2B, colonic epithelial cells HT-29, and the difference between epithelial cells and lung fibroblast cells MRC-5. MP samples were quantified and assessed for anti-influenza activity. MP samples were analyzed by flow cytometry as shown in Figure 1A; on a cytogram of forward scatter versus side scatter (SSC), the population that was smaller than 1.34 μM latex beads (R3) was gated (R1). In Figure 1B, only events included within gate R1 were further identified by their SSC/annexin FITC. The population of MPs was determined as the annexin-positive population that was absent after degradation of MPs by adding 0.1% Triton X-100 nonionic detergent that solubilizes cell membrane but does not disrupt protein aggregates (1) as shown in Figure 1C.
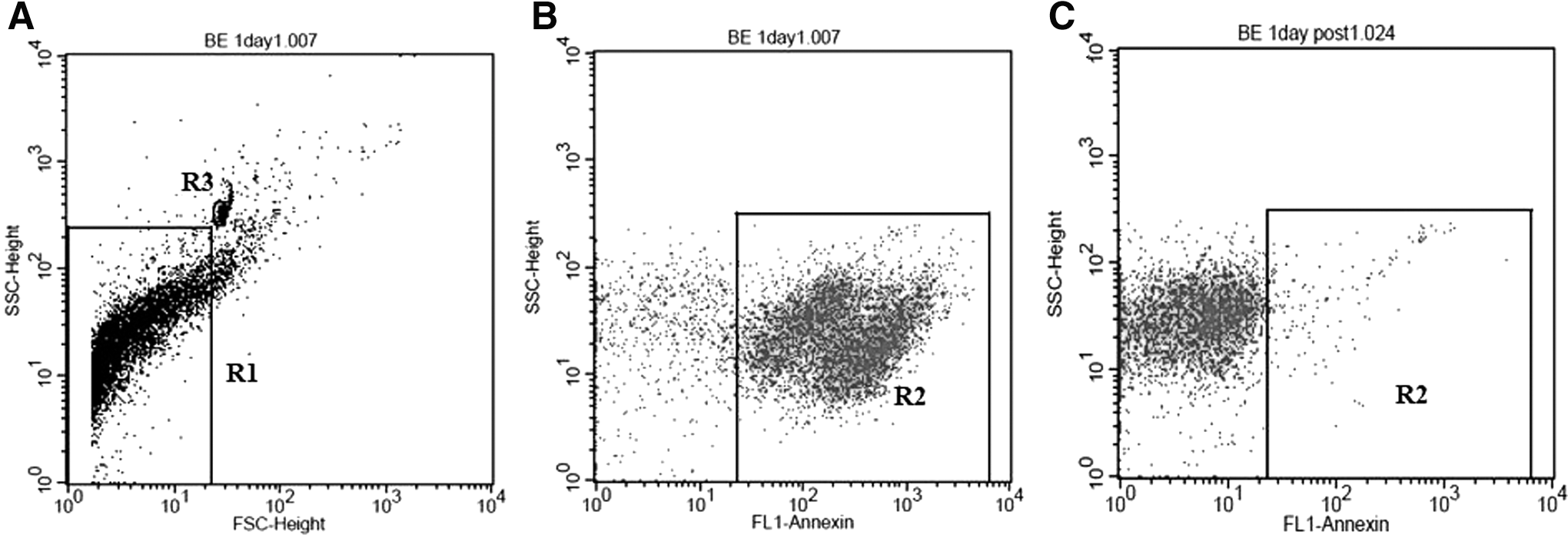
Flow cytometry analysis of MPs from culture supernatant of cell culture. In region R1
We showed that MPs were found in cell culture supernatant of A549, BEAS-2B, HT-29, and MRC-5 cells as shown in Figure 2. Among these cells, BEAS-2B cells showed the highest MP release followed by HT-29, A549, and MRC-5 cells. MPs from BEAS-2B and A549 cells had a maximum level at 30 min and 3 h, respectively, and persisted up to 24 h. However, MPs from HT-29 and MRC-5 cells continuously increased from 0 to 24 h. MPs from cell culture supernatant of BEAS-2B and A549 cells also had anti-influenza activity against H1N1 virus. However, anti-influenza activity was not found in MPs from HT-29 and MRC-5 cells (Table 1). BEAS-2B cell differentiation induced by culturing at an air–liquid interface did not increase MP release and anti-influenza activity compared with normal BEAS-2B cells as shown in Figure 2 and Table 1, respectively. Anti-influenza activity against H5N1 virus was also found in MPs from BEAS-2B and HT-29 cells (Table 1).
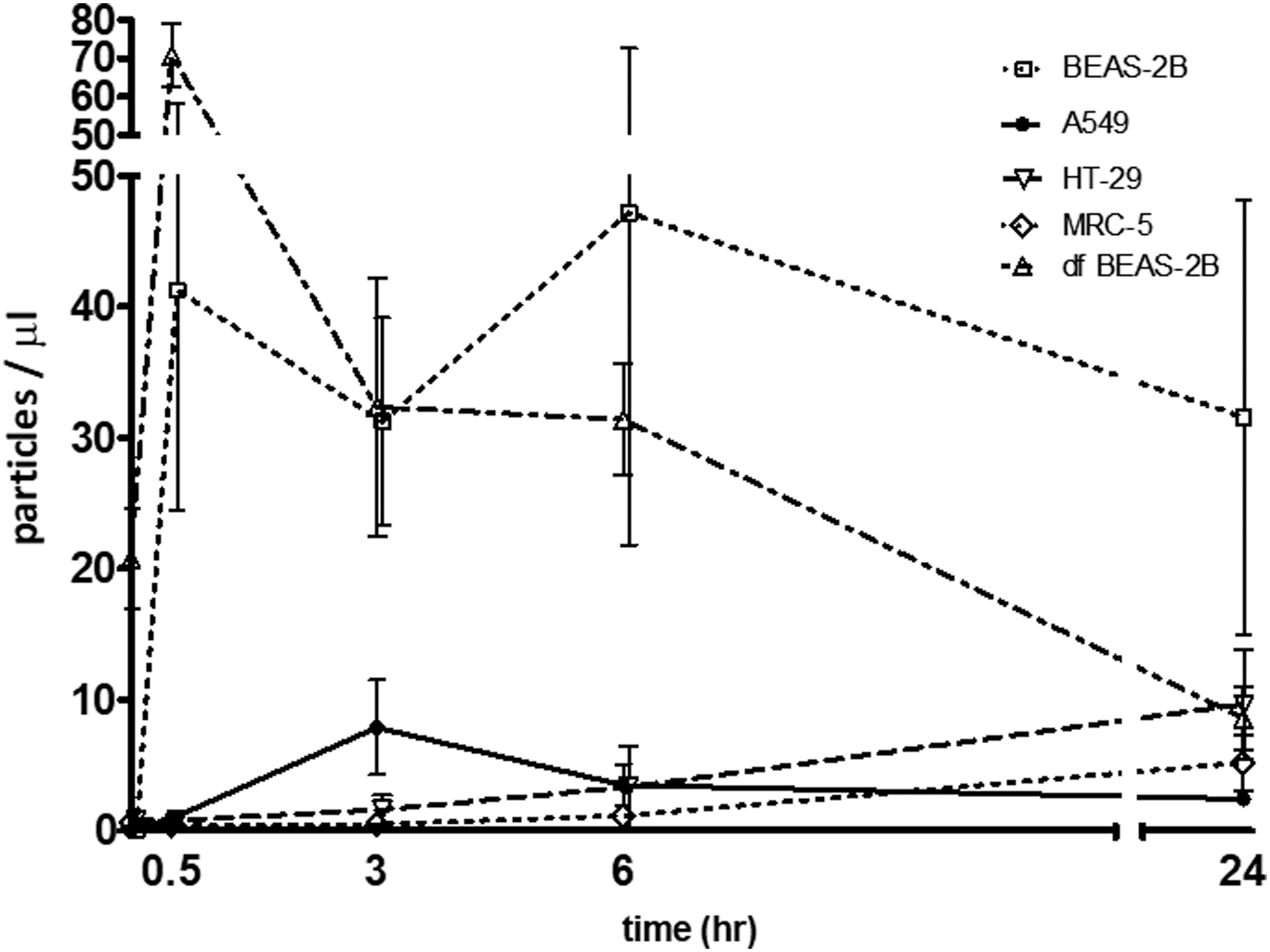
MPs released from A549, HT-29, MRC-5, BEAS-2B, and differentiated BEAS-2B cells were cultured on 12-well plate for 24 h. Then, culture supernatant was removed and 1 mL of fresh media was added before incubating at 37°C, in 5% CO2. Culture supernatants were collected at 0, 30 min, 3, 6, and 24 h. MPs were quantified by flow cytometry. The experiments were performed in triplicate. Bars and errors represent mean ± SEM.
Results are representative of three experiments.
MP, microparticle.
The sialic acid expression on the membrane of BEAS-2B, A549, HT-29, and MRC-5 cells
We next sought to investigate the expression levels of sialic acid on BEAS-2B, A549, HT-29, and MRC-5 cells. The cells were stained with FITC-conjugated lectin MAA I or SNA, specific toward α2,3- or α2,6-linked sialic acid, respectively. The results showed that compared with BEAS-2B, A549 cell has lower fluorescence intensity in MAA I and has similar level of fluorescence intensity in SNA, whereas HT-29 and MRC-5 have lower fluorescence intensity than BEAS-2B in both MAA I and SNA (Fig. 3). We further normalized MP concentration to 10 MP/μL and performed HI assay to confirm that the difference in sialic acid expression correlated with anti-influenza activity. The HI titer result indicated that MPs from BEAS-2B and A549 cells had anti-influenza activity against H1N1 influenza virus. However, only MPs from BEAS-2B had anti-influenza activity against H5N1 influenza virus (Table 2). The discrepancy result of anti-influenza activity against H5N1 between BEAS-2B and HT-29 in Tables 1 and 2 suggested that the inhibition activity from MPs correlated to the levels of (2–3) linked sialic acid expression on MP surface from both cell types.
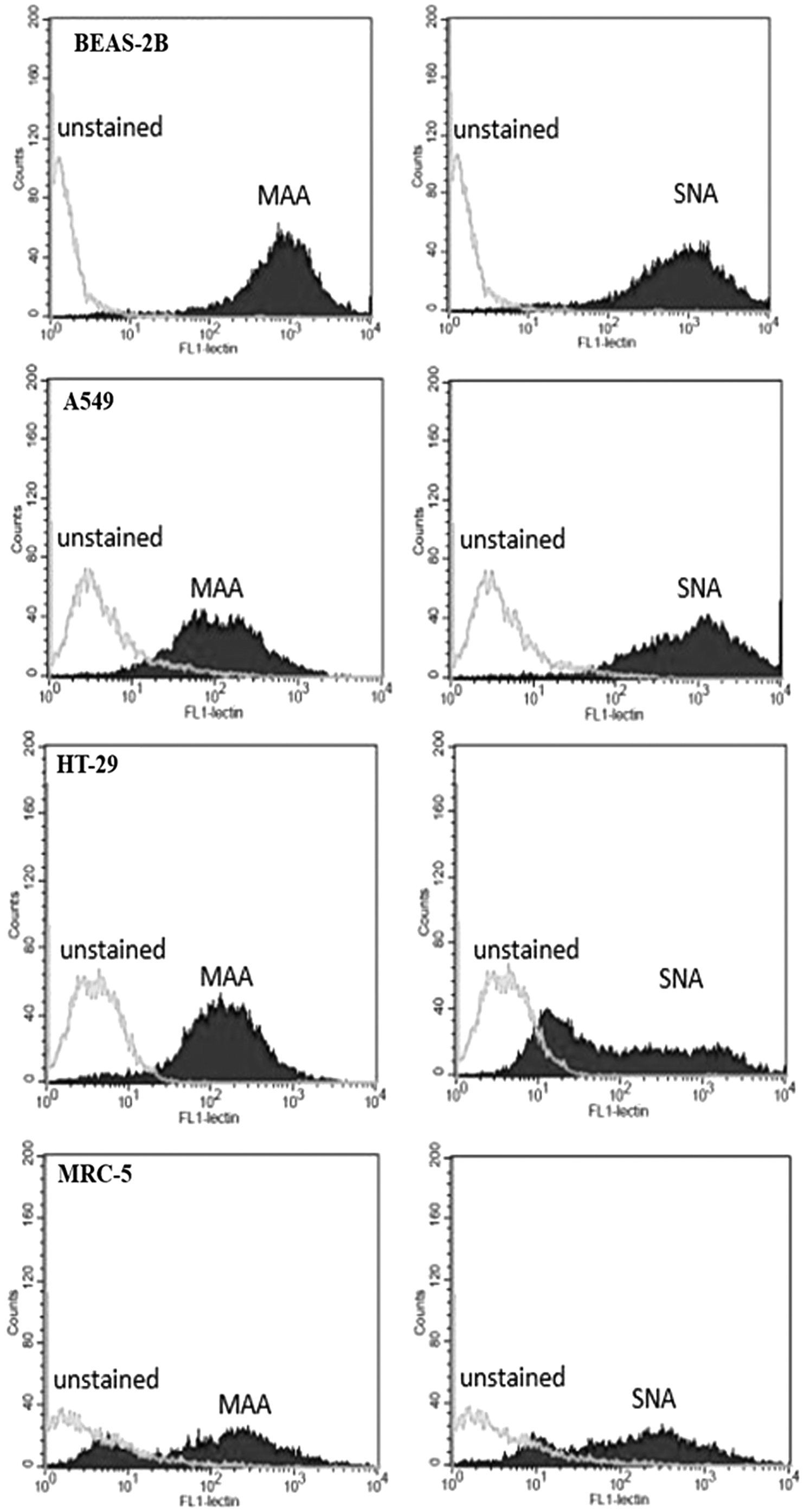
Flow cytometry histogram profile of BEAS-2B, A549, HT-29, and MRC-5 cells that were unstained or stained with 1 μg of FITC-conjugated lectin MAA I (left panel) or SNA (right panel) for 30 min at room temperature.
Results are representative of three experiments.
MP induction by MβCD and calcium ionophore A23187
To enhance innate mucosal immune defense through MP induction, epithelial cells should be the target cells. Thus, we chose only epithelial cells for investigation of the compounds that might increase MP release. The MPs from the cells that were exposed to various concentrations of MβCD and calcium ionophore A23187 for 1 h were quantified and assessed for anti-influenza activity. After being exposed to MβCD for 1 h, MP release was significantly increased in HT-29 and A549 cells but not in BEAS-2B cells (Fig. 4). Concordantly, anti-influenza activity by the HI and NT assays of MPs from HT-29 and A549 treated with MβCD were increased compared with that of cell control (Fig. 5). In contrast, after being exposed to calcium ionophore A23187 for 1 h, BEAS-2B showed a marked increase in MP release, whereas only a modest increase was observed in A459 and HT-29 (Fig. 6).
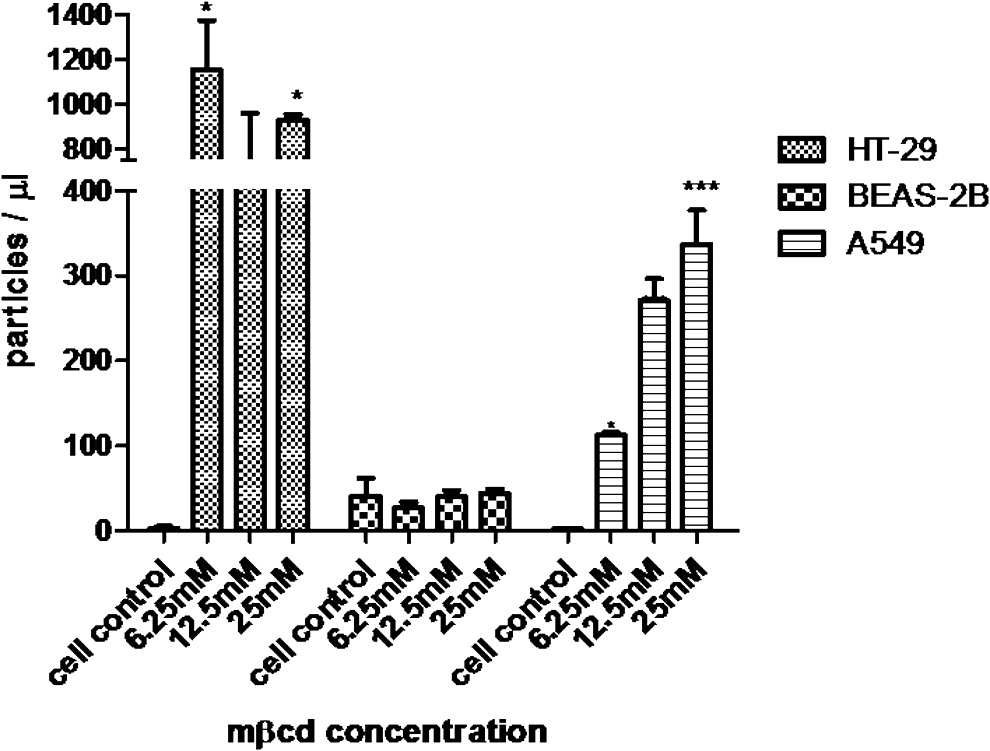
MP release from HT-29, BEAS-2B, and A549 after MβCD treatment. Cells were cultured on 12-well plate for 24 h. Then, culture supernatant was replaced with 1 mL of media in the presence of MβCD 6.25, 12.5, and 25 mM before incubating at 37°C, in 5% CO2. Culture supernatant was collected at 60 min after incubation. MPs were quantified by flow cytometry. Bar graphs represent the mean ± SEM for three experiments. Statistical significance was determined by one-way ANOVA with Tukey's test for multiple comparisons (*p < 0.05 and ***p < 0.00). MβCD, methyl-β-cyclodextrin.
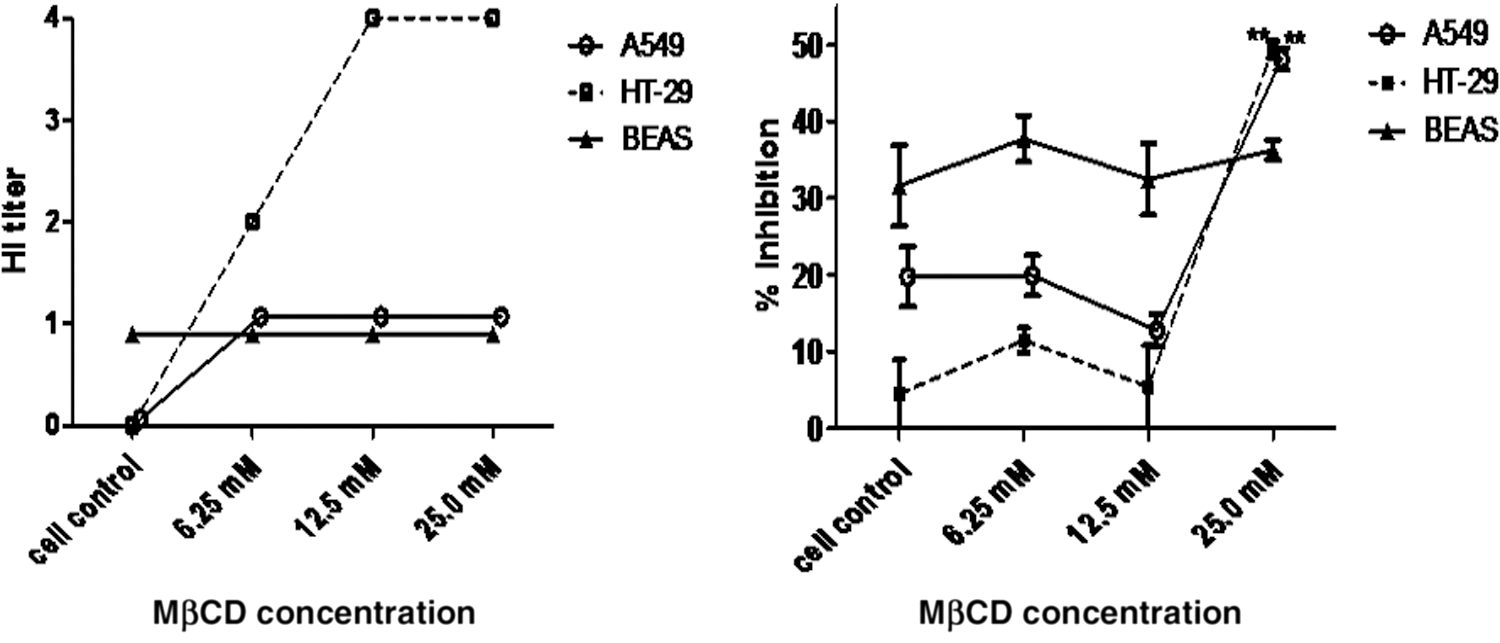
Neutralization and HI activity against H1N1 influenza virus strain A/Thailand/MVCU-13/2009 of MP release from A549, HT29, and BESA-2B cells after treatment with MβCD 6.25, 12.5, and 25 mM for 1 h. Graphs represent the mean ± SEM of triplicate results. Statistical significance was determined by one-way ANOVA with Tukey's test for multiple comparisons (**p < 0.01).
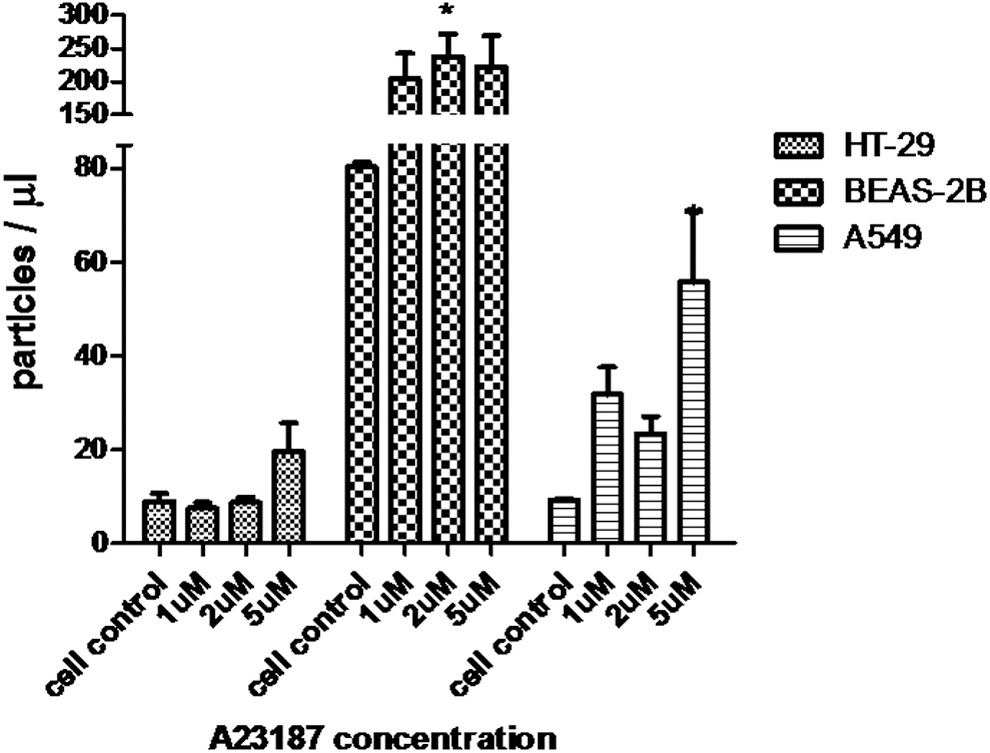
MP release from HT-29, BEAS-2B, and A549 after calcium ionophore A23187 treatment. Cells were cultured on 12-well plate for 24 h. Then, culture supernatant was replaced with 1 mL of media in the presence of calcium ionophore A23187 1, 2, and 5 μM before incubating at 37°C, in 5% CO2. Culture supernatant was collected at 60 min after incubation. MPs were quantified by flow cytometry. Bar graphs represent the mean ± SEM for three experiments. Statistical significance was determined by one-way ANOVA with Tukey's test for multiple comparisons (*p < 0.05).
MP induction by forskolin, salbutamol, and aminophylline
Among HT-29, BEAS-2B, and A549 cells, forskolin treatment increased MP release and anti-influenza activity only in HT-29 cells (Fig. 7 and Table 3). Aminophylline and salbutamol are bronchodilator drugs that increase intracellular levels of cAMP through different mechanisms (9,12). We compared the ability of MP induction between forskolin with salbutamol and aminophylline in A549 cells. MP samples from treated cells were quantified and assessed for anti-influenza activity. The results showed that there was no difference in either MPs released (Fig. 8) or HI titer against influenza virus (A/Thailand/MVCU-13/2009) (data not shown).
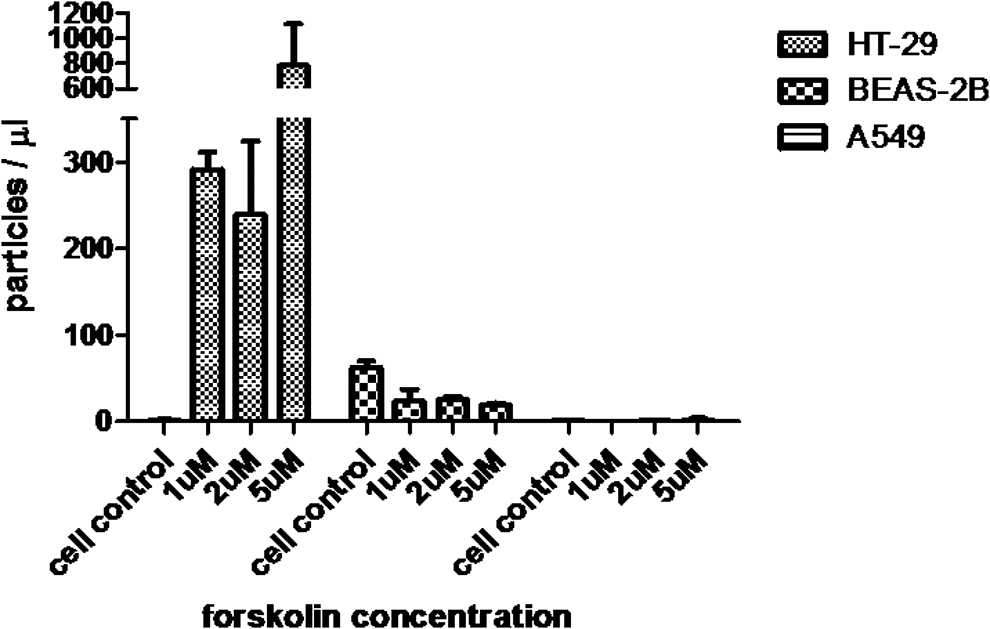
MP release from HT-29, BEAS-2B, and A549 after forskolin treatment. Cells were cultured on 12-well plate for 24 h. Then, culture supernatant was replaced with 1 mL of media in the presence of forskolin 1, 2, and 5 μM before incubating at 37°C, in 5% CO2. Culture supernatant was collected at 60 min after incubation. MPs were quantified by flow cytometry. Bar graphs represent the mean ± SEM for three experiments.
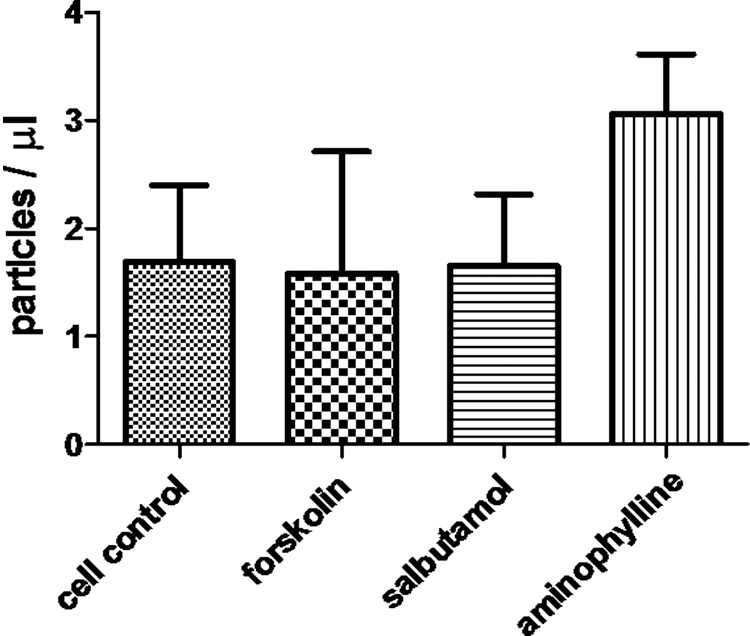
MP release from A549 after forskolin, salbutamol, and aminophylline treatment. Cells were cultured on 12-well plate for 24 h. Then, culture supernatant was replaced with 1 mL of media in the presence of forskolin, salbutamol, and aminophylline 1, 1, and 10 μM, respectively, before incubating at 37°C, in 5% CO2. Culture supernatant was collected at 60 min after incubation. MPs were quantified by flow cytometry. Bar graphs represent the mean ± SEM for three experiments.
Results are representative of three experiments.
Verification of MP preparation
To investigate whether MP fraction contains apoptotic bodies, we double stained the MP samples with annexin V and PI. MP population in gate R2 of Figure 1B was further observed for PI intensity on histogram plot compared with the apoptotic body control obtained from the supernatant of H2O2-treated cells. The result showed the PI peak of apoptotic bodies in supernatant of H2O2-treated cells (Fig. 9A), which was absent in MP population of A549 and HT-29 cells cultured in normal condition (Fig. 9B, C) and treated with MβCD (Fig. 9D, E). These indicated that the MP samples did not contain a significant amount of apoptotic bodies, and the annexin V staining in MP preparation really represented MPs. To clarify whether there was anti-influenza activity of exosomes from cell lines, the MP pellets and supernatants after centrifugation at 20,000 g for 2 h were tested for anti-influenza activity. The HI activity was only present in the MP pellets of cell treated with 25 mM MβCD for 1 h (Table 4).
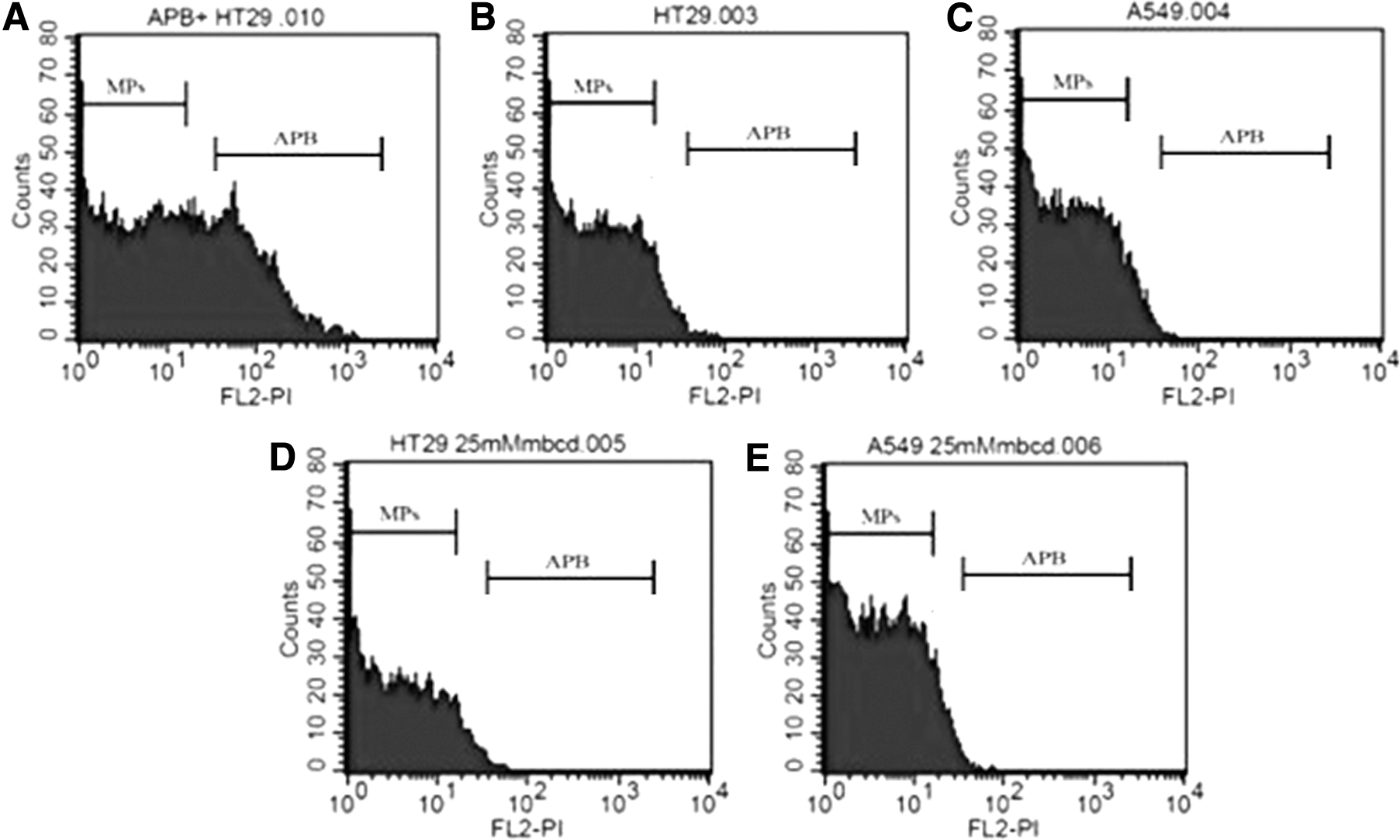
Verification of MP preparation. To distinguish MPs from apoptotic bodies, the annexin V-positive population of MP samples from A549 and HT-29 cells that culture in normal condition
MP sample obtained from HT-29 and A549 cells that were cultured in normal condition or treated with 25 mM MβCD for 1 h.
MβCD, methyl-β-cyclodextrin.
Discussion
We have recently found anti-influenza activity from MPs isolated from human BAL (29). Thus, in an effort to provide in vitro data supporting the role of MPs as an innate defense against influenza virus, we investigated quantity and anti-influenza activity of MPs from culture supernatant of A549, BEAS-2B, HT-29, and MRC-5 cells by performing flow cytometry and HI assay. In this study, MPs were isolated from culture supernatant by centrifugation at 20,000 g for 2 h. At this centrifugation speed, MPs will be precipitated while exosomes that require higher speed (19) were still suspended in culture supernatant and no anti-influenza activity was found from culture supernatants after the centrifugation. Moreover, our previous publication did not observe a significant amount of 50 ± 100 nm particles; range of exosome size in human BAL by electron microscopy and the exosome fraction by ultracentrifugation did not show anti-influenza activity (29). It suggests that epithelial cells released exosomes in low level and exosomes were not an important part of the anti-influenza activity. Since annexin V can stain both the MPs and apoptotic bodies, it is possible that the fraction of MPs using FACS contain apoptotic bodies. However, we did not observe apoptotic bodies from the MP sample isolated from cells that were cultured in normal condition and treated with MβCD.
We demonstrated that all four tested cell lines released MPs in normal culture condition in different quantities. BEAS-2B showed the highest MP release and anti-influenza activity, but anti-influenza activity could only be detected in MPs from BEAS-2B and A549 cells. BEAS-2B did not increase MP release after inducing cell differentiation by culturing at an air–liquid interface. Conversely, the level of MPs from differentiated BEAS-2B cells decreased at 3, 6, and 24 h. It may be because the differentiated BEAS-2B cells produce a mucin (32) and secreted MPs may be adsorbed by mucin, contributing to the decrease of MP level.
The levels of sialic acid expression on each cell line partially explained the difference in the levels of anti-influenza activity. BEAS-2B cells strongly expressed both α2,6- and α2,3-linked sialic acids and their MP showed antiviral activity against both H1N1 and H5N1, which is known to bind α2,6- and α2,3-linked sialic acids, respectively. In contrast, A549 cells strongly expressed only α2,6-linked sialic acid and showed antiviral activity only against H1N1 virus, whereas MRC-5 cells expressed little of both sialic acids and showed no antiviral activity against both types of viruses. Although HT-29 cells showed weak staining for both types of sialic acids, the signal of α2,3-linked sialic acid was higher than that of α2,6-linked sialic acid and their MPs showed some activity against H5N1 virus. The comparable levels of antiviral activity against H1N1 virus between MPs from BEAS-2B and A549 cells are surprising since the level of MPs in supernatant of BEAS-2B cells was much higher and both cells expressed similar levels of α2,6-linked sialic acid. Other factors may contribute to the relative antiviral activity between MPs from the two cell lines. Alternatively, it is possible that the quantification of MPs by annexin V staining may not include all MPs as there were reports of annexin V negative MPs (3) and different levels of annexin V-negative MPs from different cell types (23).
We demonstrated that MβCD, forskolin, and calcium ionophore A23187 can increase MP release and anti-influenza activity especially in HT-29 cells. It may be a specific property of HT-29 cells that allow efficient stimulation of MP release, even though it releases low level of MPs in a normal culture condition.
MβCD was the best MP inducer in our experiments. The increase in MP release by MβCD may be related to its ability to remove cholesterol from cellular membranes (18). MβCD has been used in pharmaceutical products for many years because it can interact with drug molecules to form inclusion complexes and modify the drug's properties (26). Thus, administration of drugs containing MβCD through the respiratory tract may enhance innate immunity against influenza infection. It was previously shown that inhalation of aerosolized 75 mM MβCD solutions was nontoxic in mice as assessed by the study of BAL, lung, kidney histology, blood urea, and bronchial responsiveness to methacholine (5). MβCD is also used in nasal formulations of commercial pharmaceutical products (15). However, further in vivo investigations are needed to confirm the effect of MβCD on MP induction and innate mucosal defense.
Conclusion
MPs were found to be released from various cell lines at various rates. The MPs showed anti-influenza activity, the levels of which could be partially explained by the levels of surface sialic acid. MβCD efficiently activated MP release from some cell lines and might be a candidate for treatment of mucosal viral infection by enhancing innate mucosal defense through MP induction.
Footnotes
Acknowledgments
This study was supported by a research grant from the National Research Council of Thailand. S.J. was supported by Siriraj Graduate Scholarship, Faculty of Medicine Siriraj Hospital, Mahidol University. C.B. was supported by the research assistant program, Faculty of Medicine Siriraj Hospital, Mahidol University, and P.A. was supported by a grant (IRN 60 W0002) from Thailand Research Fund.
Compliance with Ethical Standards
Research involving human participants and/or animals: No part of this study was performed with human participants or animals by any of the authors.
Author Disclosure Statement
The authors declare no conflicts of interest.