
Abstract
Signaling through interleukin (IL)-7 is essential and required for development, differentiation, proliferation, and homeostasis of T cells. However, the role of IL-7 in regulation of CD4+ T cells in chronic viral infections was not fully elucidated. Thus, the aim of the current study was to investigate the immunomodulatory activity of IL-7 to T follicular helper (Tfh) cells and its contribution to pathogenesis of chronic HCV, hepatitis C virus (HCV) infection. A total of 47 patients with chronic hepatitis C and 19 normal controls were enrolled. Serum IL-7 and proportion of Tfh cells was measured. The regulatory function of IL-7 to Tfh cells was also investigated in CD4+ T cells and CD4+ T/HCVcc-infected Huh7.5 cell cocultured system. Serum IL-7 concentration was significantly downregulated in patients with chronic hepatitis C, and was negatively correlated with HCV RNA level. Tfh frequency and Tfh-associated cytokines (IL-21 and IL-6) were also reduced in chronic HCV-infected patients. Moreover, recombinant IL-7 stimulation elevated proportion of Tfh cells and IL-21/IL-6 secretion in both HCV-specific and nonspecific manners. Furthermore, IL-7-treated CD4+ T cells exhibited elevated antiviral activities without killing infected hepatocytes, which presented as inhibition of HCV RNA, induction of antiviral proteins, and promotion of cytokine production (especially IL-21) in cocultured system. This process might be dependent on IL-6 secretion. The current data revealed that IL-7 regulated HCV-specific and nonspecific activated Tfh cells, which might contribute to viral clearance. IL-7 could be a potential therapeutic agent for the treatment of chronic hepatitis C.
Introduction
C
Interleukin (IL)-7 is essential and required for the development, differentiation, proliferation, and homeostasis of T cells, whereas deficiencies in IL-7 signaling lead to severe immunodeficiency (16,22). It demonstrated an increased serum IL-7 concentration in human immunodeficiency virus (HIV)-1 infection (25), however, Zhong et al. indicated comparable IL-7 expression in the serum in patients with chronic hepatitis B virus (HBV) infection (33), indicating an altered availability of IL-7 at various sites and different viral infections. IL-7 may influence both the generation and maintenance of T follicular helper (Tfh) cells. IL-7 plays a vital role in the generation of Tfh cells and germinal center B cells in lymph nodes in response to influenza and rabies vaccination (14,27). Exogenous IL-7 could enhance immature T cells to differentiate to Tfh cells, and this effect was specific for Tfh cells without affecting other T helper cells (27). However, controversy remains as it was reported that IL-7 signaling pathway suppressed Tfh-associated Bcl-6 mRNA expression through activation of signal transducers and activators of transcription (STAT)-5 (17). Thus, the definitive mechanism of IL-7 in the biology of Tfh still needs further elucidation in chronic viral infections.
Our previous study suggested a downregulation of serum IL-7 in patients with chronic hepatitis C (9). A more recent study also revealed an immunomodulatory role of Tfh cells in chronic HCV infection (34). Thus, we hypothesized that IL-7 also regulates Tfh function in patients with chronic hepatitis C. To test this possibility, CD4+ T cells, purified from patients with chronic HCV infection, were stimulated with recombinant IL-7. Changes in proportion and function of Tfh cells were assessed.
Materials and Methods
Subjects
The study protocol was approved by the Ethics Committee of The Affiliated Hospital to Changchun University of Chinese Medicine. Written informed consent was obtained from each enrolled subject. A total of 47 patients with chronic hepatitis C, as well as 19 healthy individuals with matched average age and sex ratio were enrolled in the current study. All patients were positive for both anti-HCV and HCV RNA for at least 6 months, and were hospitalized or followed-up in The Affiliated Hospital to Changchun University of Chinese Medicine between January 2016 and July 2016. Patients who were positive for other hepatitis virus or HIV infection, or accompanied by advanced liver diseases (decompensated liver cirrhosis, liver failure, and hepatocellular carcinoma) were excluded. No patients received antiviral or immunomodulatory therapies before baseline sampling. The clinical characteristics of enrolled subjects are shown in Table 1.
ALT, alanine aminotransferase; HCV, hepatitis C virus; NC, normal control.
Virological assessment
HCV RNA was quantitated using a commercial Real-Time Polymerase Chain Reaction (PCR) Kit (Da'an Biotech, Guangzhou, Guangdong Province, China), with detection limit threshold of 100 copies/mL.
Peripheral blood mononuclear cell isolation and CD4+ T cell purification
Peripheral blood mononuclear cells (PBMCs) were isolated by density gradient centrifugation using Ficoll-Hypaque (Solarbio, Beijing, China). CD4+ T cells were purified using the Human CD4+ T Cell Isolation Kit (Miltenyi Biotech, Bergisch Gladbach, Germany) according to instructions of the manufacturer. The purity of enriched CD4+ T cells was more than 95% by flow cytometry analysis.
HCV-infected Huh7.5 cells in vitro
Infectious HCV in cell culture (HCVcc) system was obtained from Zhonghaichao Biotech (Kunming, Yunnan Province, China). HCVcc copy, which was generated by pFL-J6/JFH plasmid (HCV genotype 2a) transfection, was quantified using the Real-time PCR-Fluorescence Quantitative Detection Kit (Da'an Gene, Guangzhou, Guangdong Province, China) with detection limitation of 2 log10 copies/mL. Huh7.5 cells were seeded into 24-well plates, and 107 copies of HCVcc were added for a 5-h incubation. Infected Huh7.5 cells were washed five times with RPMI 1640, and were cultured in RPMI 1640 supplemented with 10% fetal bovine serum (FBS), penicillin (100 U/mL), and streptomycin (0.1 mg/mL) at 37°C under 5% CO2 condition. Infected Huh7.5 cells were cultured for 48 h, and HCV RNA level in Huh7.5 cells was measured to confirm the infection.
Cell culture
Purified 106 CD4+ T cells were seeded into 24-well plates, and were cultured in RPMI 1640 supplemented with FBS, penicillin (100 U/mL), and streptomycin (0.1 mg/mL) at 37°C under 5% CO2 condition. Cells were stimulated with HCV Th epitope NS3 1248-1261 [sequence: GYKVLVLNPSVAAT (7); final concentration: 20 μg/mL] or anti-CD3/CD28 (eBioscience, San Diego, CA; final concentration: 1 μg/mL) in the presence or absence of recombinant IL-7 (PeproTech, Rocky Hill, NJ; final concentration: 1 ng/mL) for 12 h. In some experiments, 5 × 105 of purified CD4+ T cells were cocultured with 2.5 × 106 of HCVcc-infected Huh7.5 cells with HCV Th epitope NS3 1248-1261 (20 μg/mL) in the presence of or absence of anti-IL-6 neutralizing antibody (InvivoGen, San Diego, CA; final concentration: 10 ng/mL) stimulation for 48 h. Purified CD4+ T cells were also stimulated with IL-7 (1 ng/mL) for 12 h before coculture. Cells and supernatants were harvested for further experiments.
Enzyme-linked immunosorbent assay
Levels of relative cytokine production in the serum or supernatants were measured using the commercial ELISA Kits (eBioscience) according to instructions of the manufacturer.
Flow cytometry
Cells were transferred to FACS tubes, and were stained with anti-CD3-PerCP (BD Bioscience, San Jose, CA), anti-CD4-APC (eBioscience), anti-CXCR5-PE (eBioscience), and anti-ICOS-FITC (eBioscience) for a 30-min incubation at 4°C in the dark. Isotype control antibodies were used to separate positive and negative cells in the PerCP, APC, PE, and FITC fluorescence channels. In some experiments, cells were stained with Annexin V-FITC (Beyotime Biotech, Wuhan, Hubei Province, China) and propidium iodide (PI; Beyotime Biotech) for apoptosis and necrosis analyses. Samples were analyzed using FACSCalibur analyzer (BD Bioscience). Acquisitions were performed using CellQuest Pro software (BD Bioscience), and analyses were performed using FlowJo Version 8.7.2 for Windows (Tree Star, Ashland, OR).
Western blot
Cells were lysed on ice for 15 min in lysis buffer, and the supernatants were harvested by centrifugation at 12,000 g for 10 min at 4°C. Total proteins were separated on SDS-PAGE using Mini-PROTEAN 3 electrophoresis cell systems (Bio-Rad, Hercules, CA), and was electroblotted onto a PVDF membrane. The membrane was soaked in phosphate-buffered saline containing 5% non-fat milk and 0.05% Tween 20, and then incubated overnight in the presence of rabbit anti-phosphorylated STAT-5 (phospho Y694) (1:1,000 dilution), rabbit anti-STAT-5 (1:1,000 dilution), or anti-GAPDH (1:2,000 dilution) (Abcam, Cambridge, MA). Membrane was washed five times after incubation, and horseradish peroxidase-conjugated goat anti-rabbit antibody IgG (Abcam; 1:2,000 dilution) was added for an additional 2-h incubation. Antigen–antibody complexes were visualized by enhanced chemiluminescence (Western Blotting Luminol Reagent, Santa Cruz, CA).
Real-time PCR
Total RNA was isolated using the RNeasy Mini Kit (Qiagen, Hilden, Germany) according to instructions of the manufacturer. cDNA was synthesized using PrimeScript RT Master Mix (TaKaRa, Dalian, Liaoning Province, China) with random hexamers. Real-time PCR was performed using SYBR Premix Ex Taq (TaKaRa). The relative gene expression was quantified by ΔΔCT method using 7500 System Sequence Detection software (Applied Biosystems, Foster, CA). The sequences of primers were shown as following: Myxovirus resistance A (MxA) forward: 5′-ACA ATC AGC CTG GTG GTG GTC-3′, MxA reverse: 5′-CCT CCC CTA CAG TTT CCT CTC C-3; Oligoadenylate synthetase (OAS) forward: 5′-AGG TGG TAA AGG GTG GCT-3′, OAS reverse: 5′-TGC TTG ACT AGG CGG ATG-3′; β-actin forward: 5′-CAC GAA ACT ACC TTC AAC TCC-3′, β-actin reverse: 5′-CAT ACT CCT GCT TGC TGA TC-3′ (34).
Lactate dehydrogenase assay
Lactate dehydrogenase (LDH) expression in the supernatants was measured using the LDH Cytotoxicity Assay Kit (Beyotime Biotech) according to instructions of the manufacturer. LDH expression in HCVcc-infected Huh7.5 cells was defined as low-level control, whereas LDH expression in Triton X-100-treated HCVcc-infected Huh7.5 cells was defined as high-level control. The resulting value corresponding to percentage of cell death was calculated in the following equation: (experimental value−low control)/(high control−low control) × 100%.
Statistical analyses
All data were analyzed using SPSS Version 19.0 for Windows (Chicago, IL). Shapiro–Wilk test was used for normal distribution assay. Student's t-test was used for the comparison between two groups. Paired t-test was used for the comparison before and posttreatment. SNK-q test was used for the comparison among groups. Pearson correlation analysis was used for correlation analysis. A p value of less than 0.05 was considered as a significant difference.
Results
Downregulation of serum IL-7 in patients with chronic hepatitis C was negatively correlated with HCV RNA
We previously showed reduced expression of IL-7 in the serum of chronic HCV-infected patients using Luminex Multiplexing Tests (9). Herein, we measured serum IL-7 level using the ELISA Kit. Consistent with our previous results, IL-7 expression was significantly downregulated in the serum from patients with chronic hepatitis C (26.90 ± 13.98 pg/mL) in comparison with that from normal controls (NCs) (35.84 ± 20.57 pg/mL; p = 0.045, Fig. 1A). There was a negative correlation between serum IL-7 concentration and HCV RNA level (r = −0.341; p = 0.019, Fig. 1B). However, IL-7 level did not remarkably correlate with alanine aminotransferase level in the serum (r = −0.201; p = 0.176, Fig. 1C).
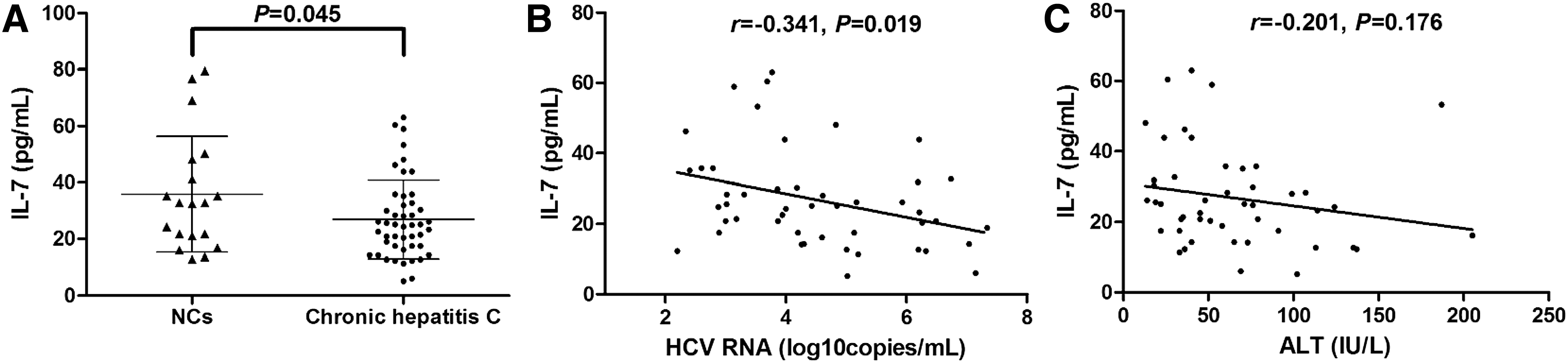
IL-7 expression in patients with chronic hepatitis C and its correlation with virological and biochemical index.
The percentage of Tfh cells and IL-21/IL-6 production were reduced in patients with chronic hepatitis C
We then analyzed the circulating Tfh cells in PBMCs isolated from NCs and patients with chronic hepatitis. The representative flow cytometry determinations of Tfh cells are shown (Fig. 2A). Activated Tfh cells were defined as CD3+CD4+CXCR5+ICOS+, whereas resting Tfh cells were defined as CD3+CD4+CXCR5+ICOS−. Total Tfh cells were defined as CD3+CD4+CXCR5+. (Fig. 2A). Activated Tfh cells within CD4+ T cells was notably decreased in patients with chronic hepatitis C (0.89% ± 0.12%) in comparison with that in NCs (0.74% ± 0.14%; p = 0.0003, Fig. 2B). Moreover, resting Tfh cells within CD4+ T cells was also reduced in chronic HCV-infected patients (14.52% ± 3.56%) when compared with that in NC (5.58% ± 1.85%; p < 0.0001, Fig. 2C). Thus, total Tfh cells was decreased in chronic hepatitis C than in NCs (15.40% ± 3.59% vs. 6.32% ± 1.86%; p < 0.0001, Fig. 2D). Moreover, serum IL-21 and IL-6 expressions, which were important for Tfh cell proliferation and differentiation, were measured by ELISA. As shown in Figure 2E and F, both serum concentrations corresponding to IL-21 and IL-6 were significantly reduced in chronic hepatitis C patients when compared with NCs (IL-21: 123.5 ± 8.90 pg/mL vs. 105.6 ± 15.39 pg/mL; p < 0.0001, Fig. 2E; IL-6: 108.2 ± 42.53 pg/mL vs. 65.25 ± 13.03 pg/mL; p < 0.0001, Fig. 2F). However, there were no remarkable correlations between Tfh cell proportion or IL-21/IL-6 secretion and viral replication/liver inflammation (p > 0.05).
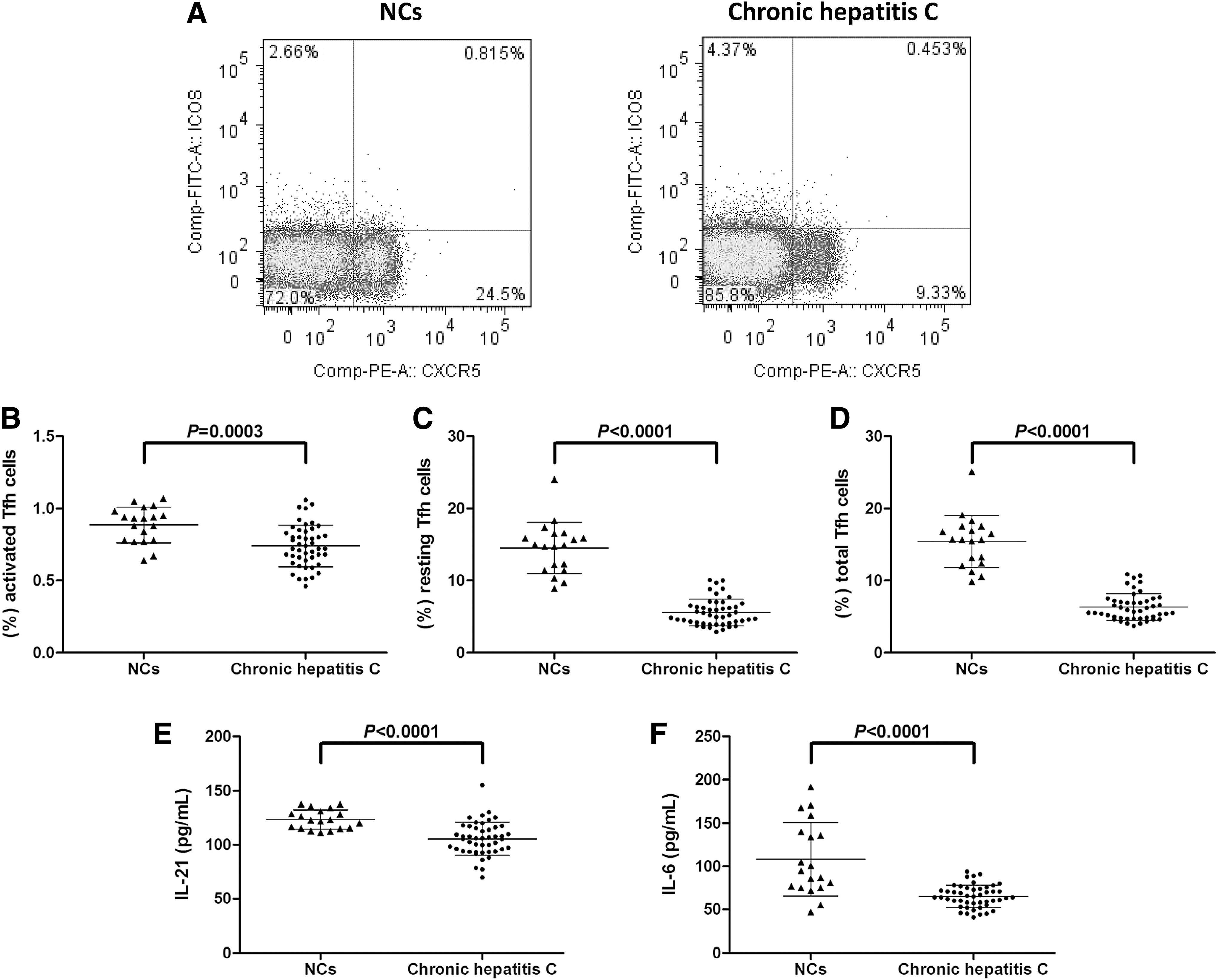
The proportion of Tfh cells and related cytokine expressions in patients with chronic hepatitis C.
Recombinant IL-7 stimulation elevated activated Tfh cell proportion and IL-21/IL-6 production in both HCV-specific and nonspecific manners
Purified CD4+ T cells from 14 patients with chronic hepatitis C were stimulated with either HCV Th peptide or anti-CD3/CD28 in the presence or absence of recombinant IL-7 for 12 h. IL-7 induced elevated expression of phosphorylated STAT-5 (pSTAT-5), confirming the functional activation through IL-7 signaling in CD4+ T cells (Fig. 3A). IL-7 stimulation increased the proportion of activated Tfh cells within CD4+ T cells in both HCV Th peptide (0.66% ± 0.14% vs. 0.93% ± 0.19%; p = 0.0006, Fig. 3B) and anti-CD3/CD28 (0.70% ± 0.16% vs. 0.89% ± 0.11%; p = 0.0039, Fig. 3B) stimulation. Moreover, both IL-21 and IL-6 expressions in the supernatants were also significantly elevated in response to IL-7 stimulation in both HCV-specific (IL-21: 80.97 ± 11.51 pg/mL vs. 107.4 ± 12.33 pg/mL; p < 0.0001, Fig. 3C; IL-6: 36.43 ± 12.01 pg/mL vs. 62.21 ± 25.37 pg/mL; p = 0.0059, Fig. 3D) and nonspecific (IL-21, 84.00 ±12.55 pg/mL vs. 109.2 ± 10.76 pg/mL; p = 0.0002, Fig. 3C; IL-6: 30.11 ± 8.91 pg/mL vs. 52.21 ± 24.62 pg/mL; p = 0.0071, Fig. 3D) manners. Moreover, it was well accepted that cell death could account for the reported differences between untreated and IL-7-treated cells. Thus, CD4+ T cells purified from seven chronic hepatitis C patients were stained with Annexin V and PI for apoptosis and necrosis analyses in response to IL-7 stimulation. Representative Annexin V/PI-stained cells, under HCV Th peptide stimulation with or without IL-7 treatment, are shown in Figure 3E. Annexin V+PI− was defined as apoptosis, whereas Annexin V+PI+ was necrosis. There were no remarkable differences in either apoptosis (3.74% ± 0.49% vs. 4.11% ± 0.48%; p = 0.103, Fig. 3F) or necrosis (5.32% ± 0.85% vs. 5.79% ± 0.67%; p = 0.095, Fig. 3F) in the presence or absence of IL-7 stimulation.
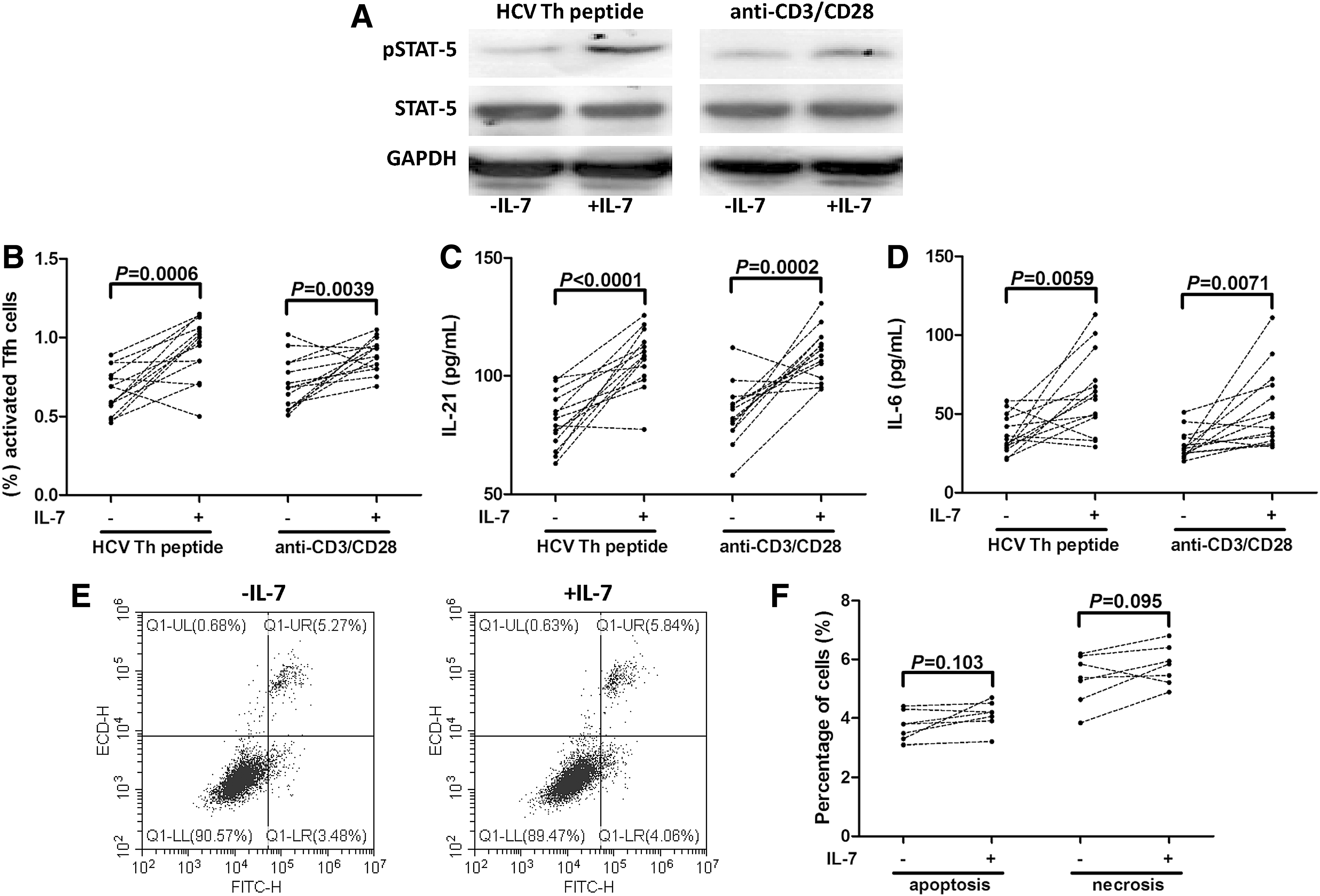
IL-7 elevated Tfh cell frequency and cytokine production of CD4+ T cells in both HCV-specific and nonspecific manners. Purified CD4+ T cells from 14 patients with chronic hepatitis C were stimulated with either HCV Th peptide or anti-CD3/CD28 in the presence or absence of recombinant IL-7 for 12 h.
IL-7 did not directly inhibit HCV replication in HCVcc-infected Huh7.5 cells
It was reported that IL-7 was also a hepatically expressed protein, which directly regulated immune cell responses (15,26). Thus, we measured IL-7 expression in the cultured supernatants from normal and HCVcc-infected Huh7.5 cells. IL-7 level revealed significant reduction in the supernatants from HCVcc-infected Huh7.5 cells than that from normal Huh7.5 cells (60.80 ± 9.83 pg/mL vs. 281.4 ± 83.94 pg/mL; p = 0.0004, Fig. 4A). Moreover, HCVcc-infected Huh7.5 cells were also stimulated with recombinant IL-7 for 48 h, and HCV RNA was measured using real-time PCR. There was no remarkable difference in HCV RNA level between HCVcc-infected Huh7.5 cells with and without IL-7 stimulation (Cells: 4.76 ± 2.11 log10 copies/mL vs. 3.78 ± 0.99 log10 copies/mL; p = 0.376; Supernatants: 4.68 ± 1.06 log10 copies/mL vs. 4.18 ± 0.89 log10 copies/mL; p = 0.441; Fig. 4B). All experiments were performed independently for five times.
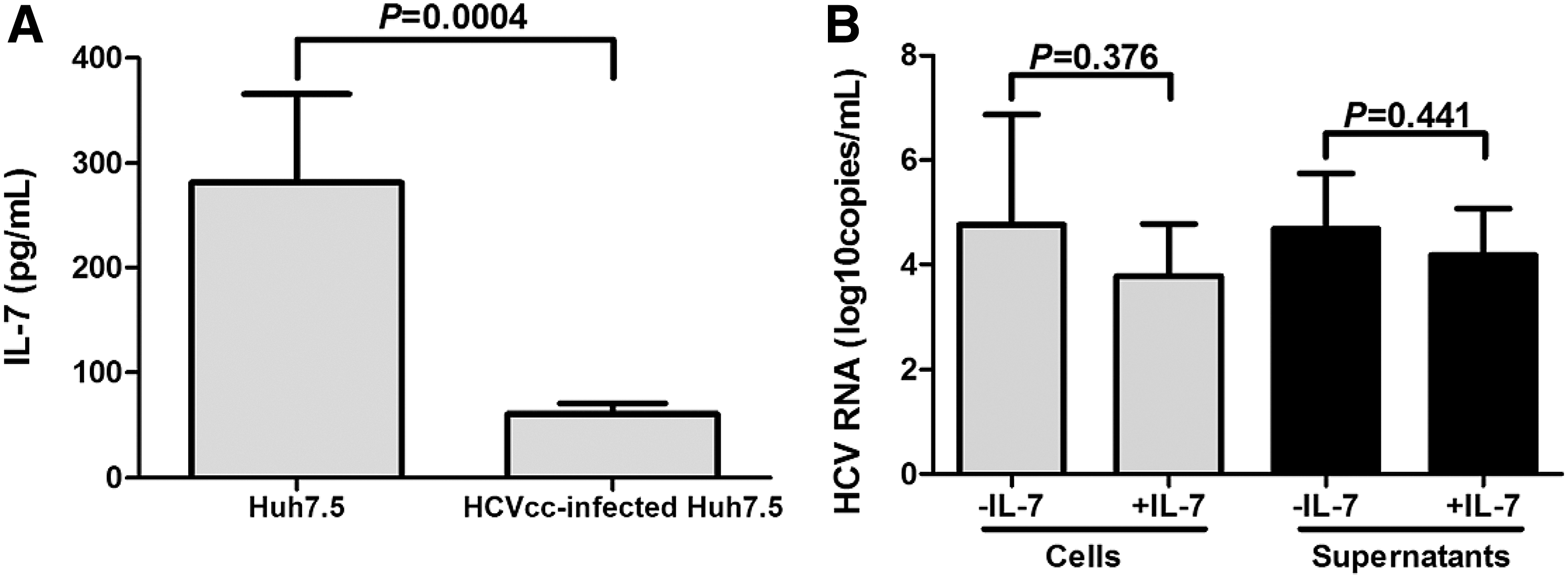
IL-7 did not directly inhibit HCV replication in vitro.
IL-7 elevated noncytolytic activities of HCV-specific CD4+ T cells, whereas anti-IL-6 blocked this process
A previous study demonstrated that IL-7 mediated multiple effects to overcome chronic viral infection, and these processes were dependent on IL-6 production, but not IL-21 secretion (21). Thus, we investigated the involvement of IL-6 in regulation of Tfh function by IL-7. Purified CD4+ T cells from 15 patients with chronic hepatitis C were stimulated with recombinant IL-7 for 12 h. Cells were washed twice, and cocultured with HCVcc-infected Huh7.5 cells for a 48-h incubation with HCV Th peptide stimulation in the presence or absence of anti-IL-6 neutralizing antibody. Cells and supernatants were harvested for further analysis. HCV RNA replication was significantly downregulated in IL-7-stimulated CD4+ T cell coculture system when compared with unstimulated CD4+ T cell coculture system (4.71 ± 1.51 log10 copies/mL vs. 5.86 ± 0.91 log10 copies/mL; p = 0.017, Fig. 5A). Anti-IL-6 antibody treatment did not affect the HCV RNA level (5.72 ± 1.11 log10 copies/mL; p = 0.697, Fig. 5A). There was a consistent trend that anti-IL-6 antibody eliminated IL-7-induced inhibition of viral replication; however, this difference failed to achieve remarkable difference (5.54 ± 1.08 log10 copies/mL; p = 0.093, Fig. 5A). However, little cell deaths were observed in coculture systems with no statistical differences among groups (p = 0.486, Fig. 5B). Moreover, IL-7-treated CD4+ T cell coculture system induced more than 10-fold increase of mRNA expressions of antiviral proteins (MxA and OAS; p < 0.0001, Fig. 5C, D). Anti-IL-6 antibody treatment did not decrease MxA and OAS mRNA expressions in cocultured cells (p = 0.602 and 0.129; Fig. 5C, D), but significantly reduced IL-7-induced antiviral protein elevation (p < 0.0001, Fig. 5C, D).
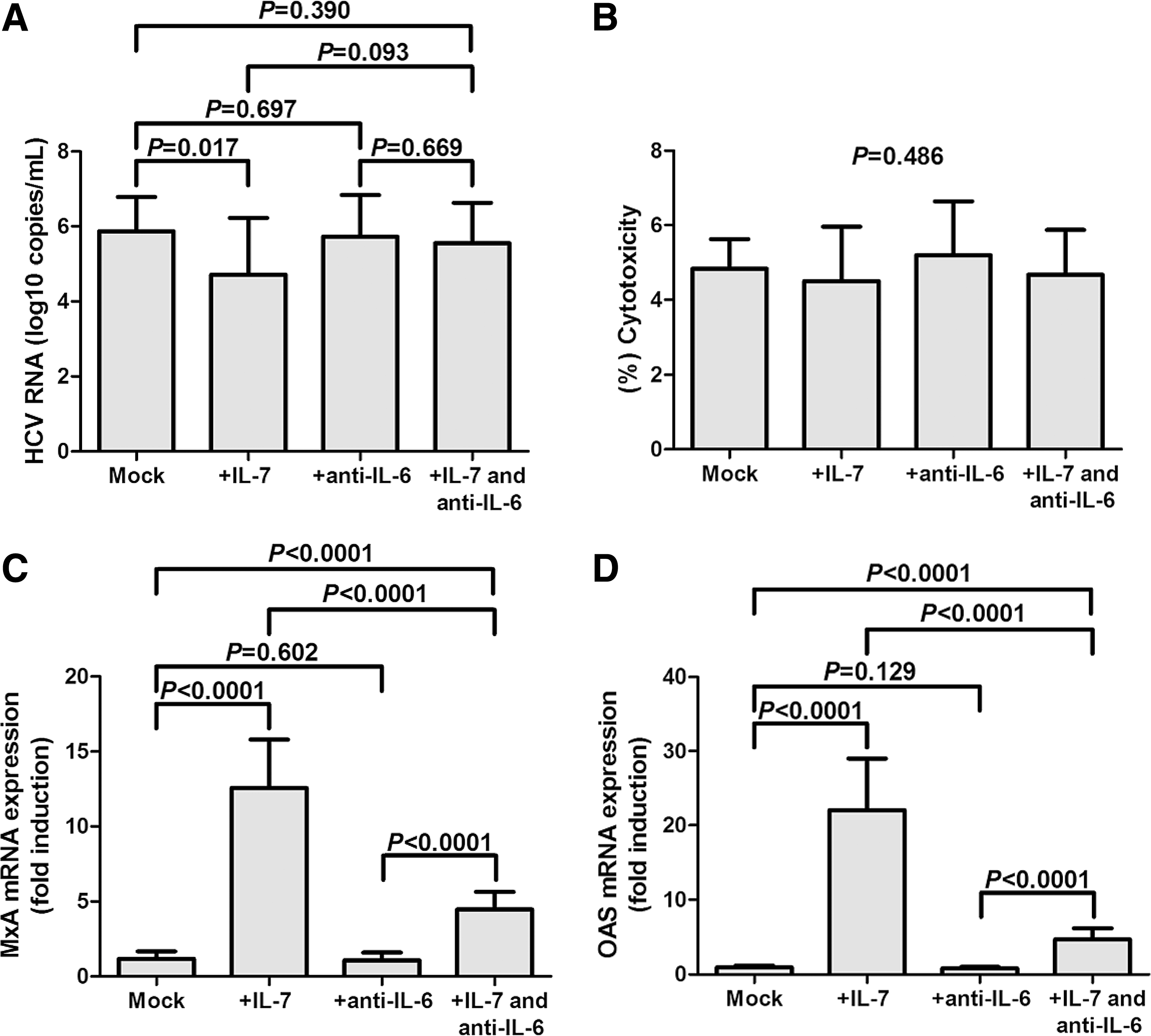
IL-7 promoted antiviral protein mRNA expression in CD4+ T cells and HCVcc-infected Huh7.5 cell coculture system, and this process might be dependent on IL-6 secretion. Purified CD4+ T cells from 15 patients with chronic hepatitis C were stimulated with recombinant IL-7 for 12 h, and then cocultured with HCVcc-infected Huh7.5 cells for a 48-h incubation with HCV Th peptide stimulation in the presence or absence of anti-IL-6 neutralizing antibody.
The levels of relative cytokines, including interferon-γ (IFN-γ), IL-4, IL-17, IL-21, and IL-22, in the cocultured supernatants were then measured. There were significant elevations of all the above cytokines in the supernatants from IL-7-treated CD4+ T cell coculture system in comparison with untreated CD4+ T cell coculture system (p < 0.05, Fig. 6A–E). Anti-IL-6 antibody treatment slightly downregulated IL-17 and IL-22 production (p = 0.05 and 0.046, respectively; Fig. 6C, E), remarkably reduced IL-21 secretion (p < 0.0001, Fig. 6D), but did not affect IFN-γ and IL-4 expressions (p = 0.330 and 0.158, respectively; Fig. 6A, B). Moreover, anti-IL-6 antibody also eliminated IL-7-induced elevation of all tested cytokines (p < 0.01, Fig. 6A–E).
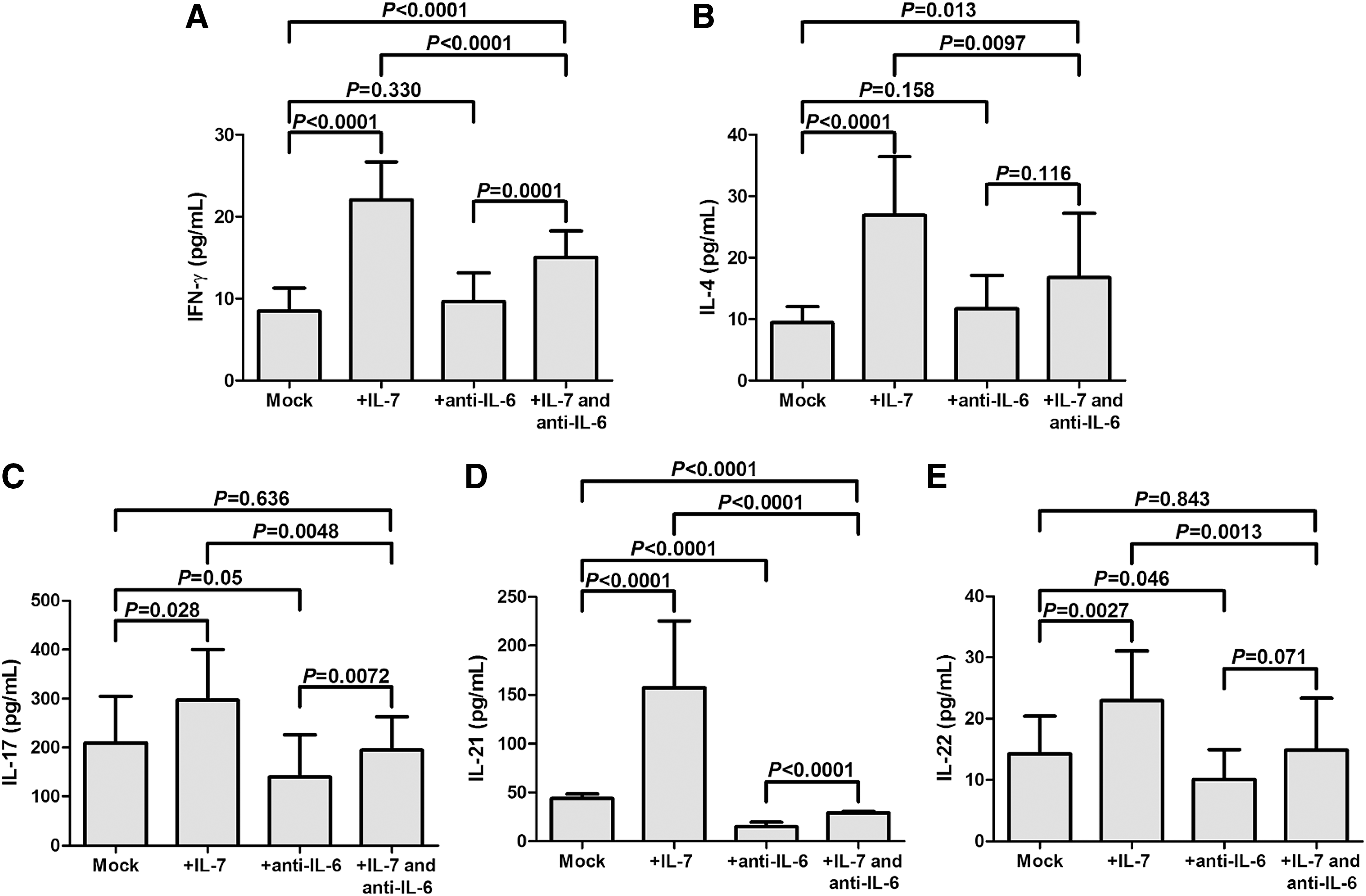
IL-7 promoted cytokine production in CD4+ T cells and HCVcc-infected Huh7.5 cell coculture system, and this process might also be dependent on IL-6 secretion. IL-7-stimulated CD4+ T cells significantly increased cytokine production. Although anti-IL-6 antibody treatment did not directly influence cytokine production, it remarkably eliminated IL-7-induced elevation of cytokine production.
Discussion
In the present study, downregulation of serum IL-7 was negatively correlated with HCV RNA in patients with chronic hepatitis C. Significant reductions of circulating Tfh cells and Tfh-associated cytokines were also investigated in chronic HCV-infected patients. Recombinant IL-7 stimulation elevated Tfh proportion and IL-21/IL-6 production probably through STAT-5 activation. Moreover, IL-7 enhanced CD4+ T cell functions, including cytokine secretion (especially IL-21) and antiviral protein induction, leading to inhibition of HCV replication. This process possibly depended on IL-6 production. The current results indicated that IL-7 depression might contribute to dysfunction of viral-specific CD4+ T cells, which promoted persistent HCV infection.
Changes in serum concentration of IL-7 seemed varied in different chronic viral infections. There was no significant elevation of IL-7 expression in HBV infection in comparison with that in healthy individuals (33). Controversy existed with the IL-7 expression in the serum of HIV-infected patients (3,11,25). However, consistent with our previous findings (9), the current results further confirmed the downregulation of serum IL-7 in chronic HCV infection. IL-7 reduction was remarkably correlated with viral replication, indicating that it might be the virus factor that induced IL-7 decrease. This was supported by the fact that HCVcc-infected Huh7.5 secreted less IL-7 than normal Huh7.5 cells, whereas inhibition of HCV replication by pegylated IFN-α and ribavirin also reconstituted IL-7 secretion in patients with chronic hepatitis C (9). Moreover, IL-7 administration was demonstrated to control chronic lymphocytic choriomeningitis virus infection and decrease systemic inflammation in vivo by enhancement of both CD4+ and CD8+ T cell functions (21,28). Recombinant IL-7 was also clinically used for the treatment of patients with JC virus granule cell neuronopathy. IL-7 therapy restored lymphocytes and reduced JC viral load in the cerebrospinal fluid (29). Although the current in vitro data did not reveal direct antiviral activity of IL-7 to HCV, IL-7 stimulation increased CD4+ T cell function in viral clearance probably through the elevation of cytokine production since there were no significant cell deaths happening in CD4+ T/HCVcc-infected Huh7.5 cell coculture system. Furthermore, IL-7 stimulation also induced mRNA expression of MxA and OAS, which were important type I IFN-stimulated genes (ISG). Hou et al. showed that type I IFN could induce IL-7 expression to maintain CD8+ T cell responses in viral hepatitis (10). However, chronic exposure to type I IFN might lead to IFN tolerance in vivo. IL-7 stimulation restored ISG and IFN mRNA expression in CD4+ T cells from chronic HIV-infected patients with immunological failure to therapy (20). The current data also indicated that IL-7 stimulation resulted in increased type I IFN responsiveness, which was important for HCV viral clearance. The above process was also accompanied by the induction of antiviral proteins and was dependent on IL-6 secretion, indicating the involvement of various immune cells and molecules (e.g., Tfh cells and Toll-like receptors), which were regulated by IL-6 signaling pathway (2,4,12).
Activated CD3+CD4+CXCR5+ICOS+ Tfh cells were important for B cell activation and clearance of various chronic viral infections through production of T cell-dependent viral-specific antibodies (30). Although several studies suggested an increase of circulating Tfh cells in chronic HCV infection (13,32,34), we demonstrated a remarkable reduction of Tfh cells and IL-21 production in the peripheral bloods of patients with chronic hepatitis C, which was consistent with the previous findings in both peripheral blood and liver residency of HCV infection (30). However, the low frequency of Tfh cells and less IL-21 production was sufficient to support B cell responses in chronic HCV infection (30). More importantly, HCV-specific CD4+ T cells revealed phenotypic and functional characteristics of Tfh cells in both acute and chronic HCV infections (24). In the current study, we also used HCV Th peptide to stimulate isolated CD4+ T cells from chronic hepatitis C patients, indicating a potential Tfh characteristic of HCV-specific CD4+ T cells in vitro to control HCV replication. However, the precise mechanism was needed to be further elucidated by in vivo studies.
In recent years, the interaction between IL-7 and Tfh cells became a hot point in the immunopathogenesis of diseases. It was shown that IL-7 signaling through STAT-5 negatively regulated Tfh generation by repression of Bcl-6 (12,17). In contrast, IL-7 was demonstrated to efficiently boost influenza-specific vaccination through enhancement of Tfh cells in lymph nodes (27), indicating the importance of IL-7 in the induction of selected Tfh cell clones. IL-7 stimulation also increased the expression of ICOS, however, slightly decreased CXCR5 level in HIV infection (6). Thus, it was hard to draw a solid conclusion of IL-7 in the regulation of Tfh cells since the IL-7/Tfh interaction might be varied in physiological and different pathological statuses. Herein, we showed that recombinant IL-7 stimulation increased both HCV-specific and nonspecific Tfh cells in isolated CD4+ T cells, which were accompanied by the elevation of IL-21 and IL-6 production. Furthermore, IL-7 also significantly increased the antiviral activity of HCV-specific CD4+ T cells, which were mostly characteristics of cytokines secretion by Th1, Th2, and Tfh cells. This process was also IL-6 dependent and Tfh cells were mostly affected since IL-6 was also a vital regulator for Tfh cell differentiation. The current data indicated that IL-7 might enhance Tfh cell function in controlling viral infection in patients with chronic hepatitis C. Furthermore, in vivo function of IL-7 in modulation of Tfh cells should be assessed for the generation of humoral immune response.
Conclusion
In summary, downregulation of IL-7 and Tfh cells might contribute to viral persistence in HCV infection. The in vitro study revealed an immunopotentiator of IL-7 in chronic HCV infection, which manifested as IL-7-regulated HCV-specific and nonspecific activated Tfh cells. IL-7 could be a potential therapeutic agent for the treatment of chronic hepatitis C.
Footnotes
Author Disclosure Statement
No competing financial interests exist.