
Abstract
There is a growing body of studies that show the important role of NS3 protein from hepatitis C virus in fibrosis. However, mechanisms of the effects of this protein on immune modulation of stellate cells remain to be investigated. Herein, the effect of NS3 protein on the expression level of suppressor of cytokine signaling (SOCS)1/3 and interleukin-24 (IL-24)-related genes was investigated in hepatic stellate cell (HSC), LX-2. Recombinant NS3 protein was added to LX-2 HSC culture. Leptin and standard medium treatments were also included in experiments as positive and negative controls, respectively. Total RNA was extracted from each well at 6, 12, and 24 h after NS3 addition. The expression levels of the fibrotic (transforming growth factor beta 1 [TGF-β], alpha-smooth muscle actin [α-SMA], and COL1A1), inflammatory (IL-6 and IL-24), IL-20R, IL-22R, and immunosuppressive genes (SOCS1 and SOCS3) were evaluated by real-time polymerase chain reaction (PCR). Recombinant NS3 protein induced activated phenotypes of LX-2 with a significant increase in the expression level of α-SMA COL1A1 (p < 0.0001) and TGF-β. Moreover, this exposure led to a meaningful elevation in the expression of IL-6. Furthermore, compared with leptin (control), after the stellate cell treatment with NS3, SOCS1 and SOCS3 gene expression induced at a comparable level. Compared with the control sample, the NS3 protein significantly increased the expression level of IL-24 and its related receptors, IL-20R and IL-22R. This study not only confirmed the previously proved inflammatory and fibrotic effect of this protein but also indicated that high expression levels of SOCS1, SOCS3, and IL-24 have a significant effect on HSC activation. Therefore, these two molecules can be used as a potential therapeutic target candidate.
Introduction
L
Liver fibrosis induced by hepatitis C virus (HCV) is a gradual process mediated by viral proteins such as nonstructural protein-3 (NS3) and core. These proteins have direct effects on liver cell signaling and metabolic pathways, and modulate immune responses indirectly that consequently cause chronic inflammation (28). HSCs present several HCV coreceptors that interact with viral proteins to promote fibrogenesis (33).
NS3 is a protein with dual enzymatic activity, protease and helicase, and plays an essential function in virus replication, pathogenesis, and modulation of innate immune system (25). A protease-deficient NS3 expressing virus has been designed instead of full sequence expressing protein for vaccine purpose to avoid the destructive protease activity of the full protein on immune response and its detrimental effects (13). Our knowledge about the possible effects of the protein in inducing liver fibrosis is still limited (25,29). Kotaro et al. indicated that NS3 protease moiety acts like a transforming growth factor beta 1 (TGF-β) ligand that resulted in fibrosis phenotype induction in HSCs. In fact, this type of signal modulates the immune response to permit the virus to establish a persistent infection (25). Another study demonstrated that the protease activity of NS3 is a determinative factor of in vitro HSC activation (18). Recently, Khanizadeh et al. showed that this fibrotic property partially depends on the regulation of HSC microRNA profile (19).
The importance of interleukin-10 (IL-10) family members, particularly IL-22, in controlling and development of fibrosis has been investigated (30). However, the possible potential of melanoma differentiation-associated gene-7 (mda-7)/IL-24 in fibrosis remains to be characterized. Recently, Jamhiri et al. indicated that the expression of this cytokine and its receptors (IL-22R1 and IL-20R2 subunits) upregulated after HSC activation, suggesting a potential role of IL-24/mda-7 in fibrosis development; however, the exact mechanism remains to be explained (15).
The inflammation processes accelerate the fibrosis mechanism through different pathways such as recruitment of fibrosis molecule secreting cells and induction of stimulatory cytokines. On the contrary, there are some proteins that suppress the immune system; among them is a family of intracellular proteins known as suppressor of cytokine signaling (SOCS) proteins. This family has eight members including SOCS1-7 and CIS that usually act as the key negative modulator of cytokine signaling, the JAK/STAT pathway, and it is suggested that they also have a regulatory role in inflammation control during liver fibrosis (6). Negative regulatory molecules, SOCS1 and SOCS3, harbor a KIR motif attributed to JAK kinase suppression. As a result, these molecules exhibit a similar inhibitory effect on inflammatory cytokine signaling (11). SOCS1-deficient mice were reported to be prone to liver cirrhosis and cancer (11). There is evidence that SOCS3 acts as a pivotal negative regulator of IL-6, leptin, and granulocyte colony stimulating factor, and also as a macrophage inhibitor that decreases production of inflammatory cytokines (24,31). However, it is not yet clear whether NS3 protein performs its fibrotic role through modulation of the expression level of SOCS1 and SOCS3.
In this study, some possible mechanisms for the fibrotic effect of HCV NS3 protein are investigated. The expression levels of two different classes of molecules, IL-24 cytokine and SOCS1/3, were evaluated on LX-2 cells after the recombinant NS3 protein treatment.
Materials and Methods
Cell culture and NS3 treatment
The LX-2 cells that characterized as an immortalized human stellate cell line were kindly donated by Professor Scott Friedman (Mount Sinai School of Medicine, New York). LX-2 cells can be used as a suitable model for molecular and cellular studies on human HSCs because of its typical features of partially active stellate cells (34). The cells were grown in Dulbecco's modified Eagle's medium (DMEM; Gibco) supplemented with 5% fetal bovine serum (Sigma, St. Louis), penicillin–streptomycin, and 2 mM
Real-time polymerase chain reaction analysis
Total cellular RNA was isolated from LX-2 cells by RNA extraction kit (Cinnagen, Inc., Iran). The quantity and quality of the obtained RNA were checked by measuring the ratio of the optical density (OD) of 260/280 nm using Nanodrop™ spectrophotometer that was then stored at −80°C until complementary DNA (cDNA) synthesis. The cDNA was synthesized using 1,000 ng of total RNA in a first-strand cDNA synthesis reaction using RevertAid™ First Strand cDNA Synthesis Kit (Fermentas, Inc.). Quantitative real-time polymerase chain reaction (PCR) was performed using ABI StepOnePlus Real-Time PCR and RealQ Plus 2× Master Mix Green (Ampliqon, Inc.). In each reaction, 200 nM of each primer (Table 1) was added to target the specific sequence. The specific primer pairs sequence for evaluation of messenger RNA (mRNA) levels of TGF-β, alpha-smooth muscle actin (α-SMA), COL1A1, IL-6, SOCS1, SOCS3, IL-24/mda-7, IL-20R2, IL-22R1, and phosphoglycerate kinase (PGK) are listed in Table 1. The human PGK was used as reference gene for normalization of the reactions. RNA extract was used as the negative control of cDNA synthesis. The real-time PCR conditions were set for 10 min at 94°C followed by 40 cycles of 15 sec at 94°C, 60 sec at 60°C, and extension steps. The amplification signals of different samples were normalized to PGK Ct (cycle threshold), and then delta-delta CT (2−ΔΔCT) method was applied for comparing mRNA levels of activated versus control (normal LX-2), which represented as fold change in the data analysis.
IL, interleukin; mda-7, melanoma differentiation-associated gene-7; PGK, phosphoglycerate kinase; SOCS, suppressor of cytokine signaling; TGF-β, transforming growth factor beta 1; α-SMA, alpha-smooth muscle actin.
Statistical analysis
All data are presented as mean ± standard error of the mean of three independent experiments. One-way analysis of variance and Tukey's test were used for comparing the mean values of two groups using Prism software (V6; GraphPad Software, Inc.). Values of p < 0.05 were considered statistically significant.
Results
Recombinant NS3 protein induced a measurable fibrotic and inflammatory impact
The NS3 protein treatment left a fibrosis-like phenomenon on LX-2 cells. The morphological view of LX-2 cells demonstrated that they became activated after NS3 or leptin treatment regarding the presence of veiled projections on cells compared with the control (data not given). Moreover, the quantitative PCR (qPCR) analysis also revealed that both NS3 protein and leptin treatment induced the activation of LX-2 cells as measured through TGF-β, α-SMA, COL1A1 (fibrotic genes), and IL-6 (an inflammatory cytokine) mRNA expression analysis.
Gene product expression analysis revealed an increasing trend for three examined fibrosis-related genes at different times. These results indicated an increase in the level of TGF-β (1.70 and 2.74 folds, respectively) 12 and 24 h after NS3 protein treatment (p = 0.0014 and p < 0.0001). Moreover, the expression level of α-SMA increased up to 1.54, 2.25, and 3.89 times at 6, 12 (p = 0.0133), and 24 h (p = 0.0026) post-NS3 treatment, respectively. The mRNA expression level of COL1A1 increased significantly by 2.59 and 3.06 folds in the presence of NS3 recombinant protein at 12 (p = 0.0033) and 24 h (p < 0.0001), respectively.
Furthermore, the result demonstrated that the expression level of IL-6 increased significantly up to 4.03 times at 12 h, but started to fall down near baseline level (1.99 times at 24 h post-treatment) (p < 0.0001 and p = 0.0149, respectively). A measurable drop in IL-6 mRNA level showed a different trend of regulation by NS3 protein, especially when compared with leptin treatment, as shown in Figure 1. In brief, NS3 exhibited a similar pattern of activity on LX-2 cells comparable with leptin-treated cells.

Effects of recombinant NS3 protein on TGF-β, α-SMA, COL1A1, and IL-6 gene expression levels. The mRNA levels were compared with normal LX-2, as negative group, by qRT-PCR. The expression analyses were performed for measuring TGF-β
SOCSs genes were upregulated after NS3 treatment
To clarify the effect of NS3 protein on the expression profile of SOCSd modulatory pathway, levels of SOCS1 and SOCS3 were quantified by qPCR. In leptin-treated cells (positive control), increasing levels for both genes were detected. Leptin significantly upregulated the expression of SOCS1 and SOCS3 up to 3.25 and 3.57 folds, respectively, compared with control cells.
Similarly, the expression level of SOCS genes increased after NS3 treatment, although with different pattern (Fig. 2). In the case of SOCS1, NS3 recombinant protein induced upregulation up to 3.42 times at 12 h post-treatment (p = 0.0005); however, a significant decrease was observed during the next 12-h incubation (down to 1.2-fold).
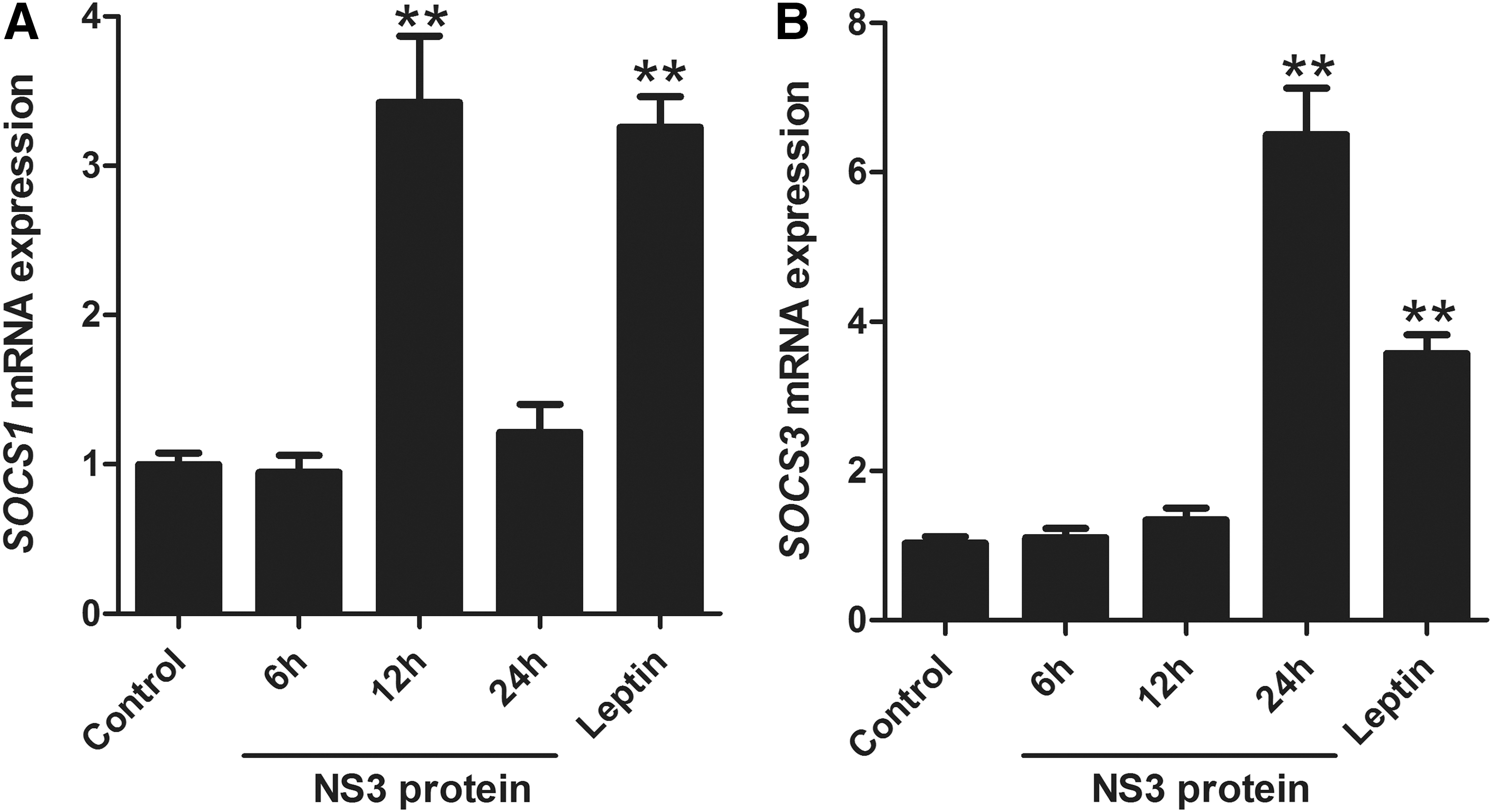
Effects of recombinant NS3 protein on SOCSs genes expression.
In the case of SOCS3, a different pattern was observed in which at the first 12 h, the expression pattern was almost similar to that of negative control (1.34-fold increase), then during the next 12 h, the expression pattern showed a remarkable increase up to 6.58-fold (p < 0.0001). These findings support the role of NS3 protein in stellate cell activation by inducing SOCS gene (responsible for cell homeostasis) production.
IL-24 and its receptors were upregulated by NS3 protein exposure
The expression levels of IL-24, a member of the IL-10 family, and its receptors IL-20R and IL-22R were measured in LX-2 cells treated by recombinant NS3 protein (400 ng/mL). The qPCR demonstrated that the expression levels of these genes were upregulated in the presence of NS3 protein compared with negative control (normal cells).
As shown in Figure 3, the expression level of IL-24 significantly increased up to 2.50, 3.63, and 3.23-folds at 6, 12, and 24 h post-treatment (p = 0.0113, p = 0.0002, and p = 0.0011, respectively). The expression levels of both IL-20R and IL-22R significantly increased up to 1.92 and 2.21 times 24 h post-treatment (p = 0.0041 and p = 0.0191, respectively). Moreover, in the positive control, leptin induced significant upregulation of the expression pattern of all three genes.
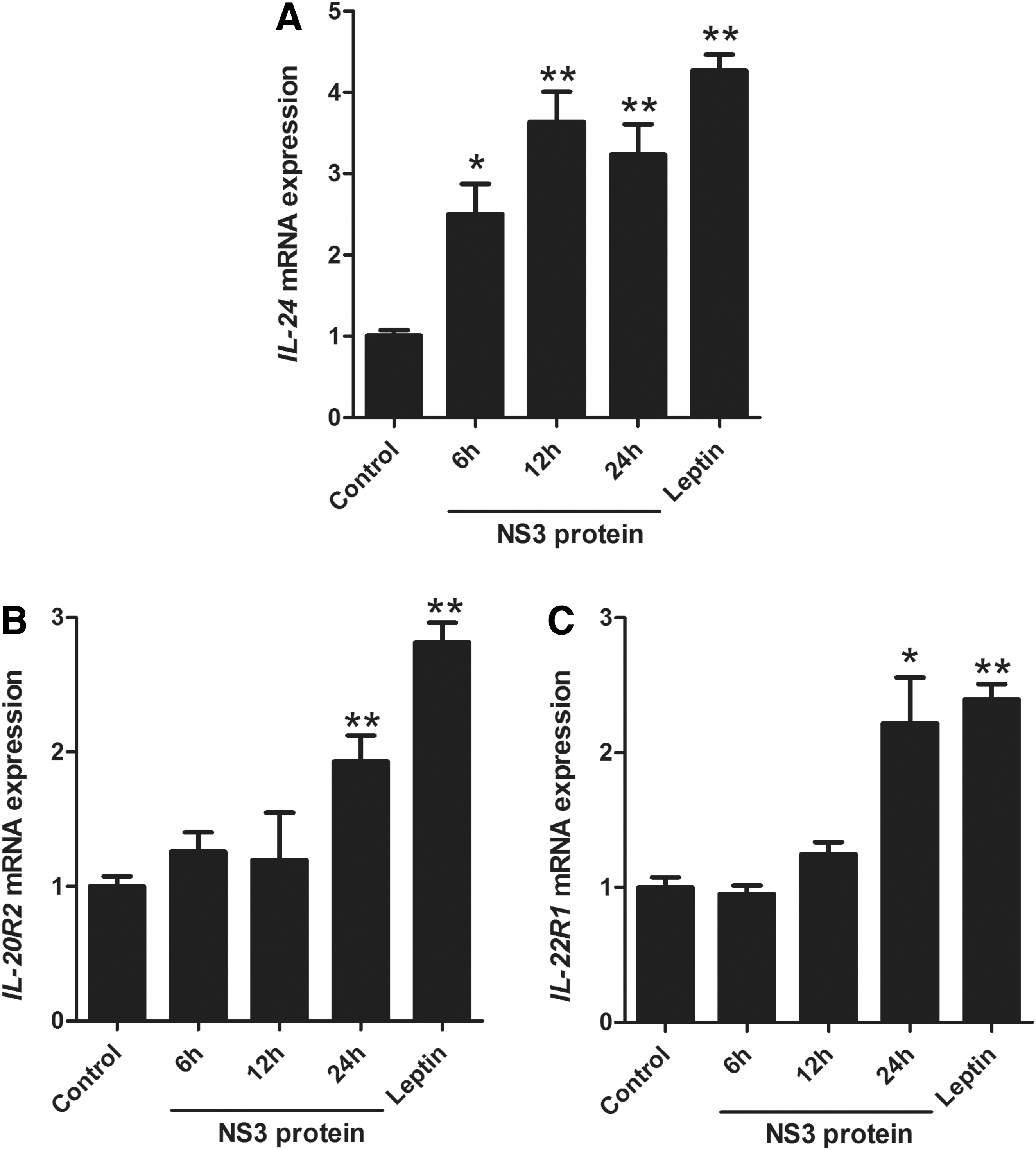
Effects of recombinant NS3 protein on IL-24 and its receptors (IL-20R2 and IL-22R1) gene expression.
Discussion
The encoding product of NS3 gene is determined as an essential factor of HCV pathogenesis in liver tissue (10). Several studies have been conducted to describe the fibrogenic activity of HCV proteins, and it is shown that some viral proteins such as core and NS3 can activate HSCs through the release of inflammatory and fibrotic mediators (3,9,17). However, it is still poorly defined how they relate to the immune response during HSC activation. Although the NS3 protein can exhibit a similar role as TGF-β in stimulating the fibrosis pathway (production of collagen and α-SMA), its activity in the proinflammatory induction of IL-6, IL-24, or immune regulator molecules such as SOCS1 and SOCS3 needs to be explored. This study investigated the effect of NS3 on the changes in the expression levels of SOCS1/3, IL-24, and its related receptors during the stellate cell activation process, in LX-2 cell line, a suitable model.
To determine the effect of NS3 in the fibrosis process, COL1A1, α-SMA, and TGF-β gene expression levels were examined. The results demonstrated that in NS3 and leptin-treated cells, NS3 significantly enhanced the expression of such proteins. NS3 protein acts as a fibrotic molecule and increases the expression level of fibrotic markers that are responsible for HSC activation, which is in agreement with previous findings by Khanizadeh et al. (19) and Sakata et al. (25) about the fibrotic effects of NS3 protein on HSCs.
We also showed that the IL-6 proinflammatory gene expression in treated HSCs was significantly enhanced in the NS3-treated cells, which is a confirmation of the proinflammatory effect of this protein. Our results demonstrated that the level of IL-6 gene expression was enhanced greatly during the first 6 h after the treatment and reached its highest after 12 h. However, there was a decreasing trend afterward. The study by Bataller et al. indicated that the recombinant NS3 protein is able to increase the level of intracellular calcium and reactive oxygen species in rat and human HSCs (3). The exposed HSCs to recombinant NS3 protein trigger various intracellular signaling pathways and secretion of proinflammatory cytokines, further supporting its fibrotic effect (27). It is shown that NS3 protein induces TGF-β production and activates its downstream signaling that triggers the process of fibrosis using Smad pathway, which reveals another part of the fibrosis mechanism (25). The study by Bansal et al. (2) on a transgenic rat expressing NS3-4A showed that its endogenous expression resulted in fibrosis development by activation of rat HSCs, especially at the acute phase, and emphasized that the NS3 protein contributed to fibrosis through induction of liver inflammation in the acute phase (2). Furthermore, NS3/4A changes immunomodulatory effects by triggering nuclear factor kappa B (NF-κB) activation and TNF-α production (2, 4).
Our result showed a similarity in expression patterns of IL-6 and SOCS1. A possible explanation is the direct role of the SOCS1 as a negative feedback in inflammatory stress control because of the cytokine. In agreement with a previous in vivo study, we showed that SOCS1 expression has a positive effect in controlling fibrosis (35). SOCS1 is able to migrate to the nucleus and perform its activity by inhibiting NF-κB action (36). It is expressed in response to lipopolysaccharides stimulation in macrophages, and without this, molecule cells will have a very high expression level of proinflammatory cytokines such as IL-6 (36). Despite the limited studies on the effects of this molecule on fibrosis, it is predictable that this molecule reduces the severity and development of fibrosis by reducing tissue inflammation of the fibrous areas.
Similar results were observed for SOCS3. Consequently, SOCS3 ameliorates fibrosis process by suppressing inflammation (36). Compared with control cells, SOCS3 gene expression increased significantly only after 24 h after exposure to NS3 protein. Therefore, our findings suggested an inhibitory role for SOCS1 and SOCS3 in the inflammation and fibrosis. We showed that in acute induction of fibrosis response, the gene expression of these proteins increases during stress phase and fibrosis induction; thus, it is likely that these molecules play a controlling role in the acute phase of fibrosis. Considering these facts, SOCS1 and SOCS3 could served as potential inhibitory targets in the chronic phases. In general, SOCS3 gene expression showed a slower trend in comparison with the expression of the IL-6 and SOCS1, which indicated control effect and delayed negative feedback role in the process of fibrosis stress. Due to this observation, further comprehensive investigations seem required.
The results demonstrate that SOCS1 gene level was increased after 12 h, whereas SOCS3 gene expression was increased after 24 h. This could be related to the effects of other inflammatory cytokines, but not directly to the effect of NS3. To justify this perspective of blocking cytokine receptors in HSC cells needs to be investigated. SOCS1 is directly affected by NS3 protein stimulation, whereas SOCS3 feedback initiated by cytokine stimulation happens afterward. Furthermore, looking at feedback loop of SOCSs, it is clear that the SOCS1 and SOCS3 stimulation and signaling pathways are partially different. SOCS1 is stimulated by IL-2, interferon gamma (IFN-γ), and granulocyte-macrophage colony-stimulating factor, whereas SOCS3 is stimulated by IL-6, IL-10, and IFN-γ. Moreover, they also have different signaling pathways for negative control (36).
The NS3 protein can activate inflammatory response pathways by binding to the toll-like receptor (TLR) (8). Several mechanisms for the inflammatory process of the HCV proteins have been suggested, for example, core protein through the MAPK pathway, and/or activation of TGF-β1 signaling and TLR2, which can increase the expression of fibrogenic and inflammatory genes. In addition, NS3 protein can increase the expression level of TGF-β1 gene in HSCs (25), which supports the function of NS3 protein on HSC cells and explains how it initiates the inflammatory pathways.
Although the anticancer role of IL-24/Mda-7 has been proven on different types of malignancies, its effects on other diseases especially in fibrosis process require further investigation (7, 23, 32). Therefore, the last objective of this study was to evaluate the importance of IL-24 and its receptors (IL-20R and IL-22R) on LX-2 cells activated by NS3. After exposing HSCs to NS3 protein, the level of gene expression of IL-24 was enhanced during the first 12 h, whereas there was a slightly decreasing trend during the next 12 h after the exposure. Therefore, it is concluded that IL-24 plays a regulatory role in the fibrosis, which was demonstrated in our previous study (16). In that study we indicated that the expression levels of IL-24/mda-7 and its cognate receptors were upregulated after HSC activation (15). However, in another study, we showed that ectopic expression of IL-24/mda-7 could induce apoptosis and senescence on activated HSCs, a promising finding for the application of IL-24 to control fibrosis development (16). This controversial finding is related to the level of IL-24 in cells, in which when the amount of IL-24 is more than normal (ectopic level), there is a greater chance of apoptosis.
IL-22 cytokine mediates tissue repair in liver cells through several mechanisms (in general, this cytokine protects the tissues from damage). For example, it is suggested that induction of HSC senescence by IL-22 is mediated by initiation of JAK/STAT3, SOCS3, and p53 signaling pathways, resulting in the repression of liver fibrosis (5, 26). In addition, the transgenic mice of IL-22 are protected from liver fibrosis through increased HSC senescence by binding to IL-10R2 and IL-22R1, increasing p53 expression, and upregulating STAT3 (22). Liver damage is similar to other chronic wound healing response, which consequently leads to liver fibrosis. Moreover, it is also reported that rat IL-24/mda-7-like molecule (C49A) and the mouse IL-24/mda-7-like molecule (FISP) increased during wound repair (26). This observation led to the assumption that in vivo expression of IL-24/mda-7 may have an effect on fibrogenesis process.
To date, most experiments have focused on IL-22, one of the best members of this subfamily that has been reviewed. Similar to IL-24/mda-7, IL-22 promotes the HSC senescence through STAT3 and p53 molecules, resulting in the repression of liver fibrosis. It is also suggested that IL-22 induction of HSC senescence was mediated by the initiation of a STAT3-SOCS3-p53 signaling pathway (20). In addition, the IL-22 transgenic mice are protected from liver fibrosis through increased HSC senescence by binding to IL-10R2 and IL-22R1, increasing p53 expression, and upregulating STAT3 (20).
Acute induction of fibrosis response showed that the IL22 gene expression was increased in the acute phase of stress and expression of fibrosis. Thus, similar to TGF-β, this protein is likely to be an inhibitor of fibrosis in acute phase. A previous study showed that IL-22 has antifibrotic properties in mouse model and in vivo conditions (21). Except our previous study, the possible fibrotic role of this protein has not been previously discussed (16). Therefore, investigation of the actual antifibrosis role using appropriate models is suggested in future studies.
Our results also indicated an increasing trend for expression of IL-20R and IL-22R, in which after12 h of exposure of HSCs to NS3 protein, lower expression levels of IL-20R and IL-22R were observed than of the control cells, whereas after 24 h, twofold increase was observed in the receptor gene expression. Overexpression of the cytokine receptors by HSCs after being exposed to NS3 protein indicated that they need IL-24 and its receptors to maintain their stable conditions. This means that to cope with fibrotic stress, HSCs increase the expression of receptor genes (IL-22R and IL-R20) and reduce the inhibitory effect of IL-24 protein to moderate fibrosis.
For further studies we suggest considering experiments on animal model, considering another protein as control, evaluation of the protein level, and assessment of a higher number of inflammatory cytokine.
Conclusion
In conclusion, activation of LX-2 cells by exposing to recombinant NS3 protein led to an increase in the expression level of SOCS1 and SOCS3 modulatory genes as a negative feedback response. Moreover, the expression levels of IL-24, IL-22R, and IL-20R were increased. These increasing levels suggest their possible roles in fibrosis process. Taken together, this study suggests a new potential target for further investigation.
Footnotes
Acknowledgment
This research was funded by Shiraz University of Medical Sciences (Nos. 13145 and 11783).
Author Disclosure Statement
No competing financial interests exist.