
Abstract
Human T cell lymphotropic type 1 (HTLV-1)-associated myelopathy/tropical spastic paraparesis (HAM/TSP) is a chronic viral neuroinflammatory disease, which leads to damage of the central nervous system. Inflammatory responses and mediators are both involved in the pathogenesis of the disease and in determining its outcome. High-Mobility Group Box 1 (HMGB1) is a chromatin-associated nuclear protein acting as a signaling molecule in cells after binding to its receptors. Receptor for advanced glycation end products (RAGE) is a transmembrane multiligand receptor that binds to HMGB1. HMGB1-RAGE signaling has an important role in inflammatory and infectious diseases. Inhibition of HMGB1 activity reduces the inflammation in immune-associated diseases. In the present study, we examined the gene expressions and plasma levels of HMGB1 and its receptor RAGE in HAM/TSP patients, HTLV-1-infected asymptomatic carriers (ACs), and healthy controls. Peripheral blood mononuclear cells were collected from all the groups and complementary DNA (cDNA) was synthesized. HMGB-1 messenger RNA (mRNA) expression was quantified by real-time polymerase chain reaction (PCR) TaqMan method, and plasma levels of HMGB1 and soluble RAGE (sRAGE) were measured by enzyme-linked immunosorbent assay (ELISA). The mRNA expression of HMGB1 was the same among the groups (p > 0.05). No significant difference in the plasma levels of HMGB1 was observed between the groups (p > 0.05). The plasma levels of sRAGE were higher in ACs than HAM/TSP patients, and a significant difference was observed between the two groups (p < 0.001). Our results showed that sRAGE could play a potential role in the control of inflammatory response in HTLV-1 carriers through the inhibition of HMGB1 signaling and potentially could be used as an indicator for evaluation of HAM/TSP developing in HTLV-1-infected individuals.
Introduction
Human T cell Lymphotropic type 1 (HTLV-1) is a pathogenic virus, which is associated with two main types of disease; adult T cell leukemia (ATL) and HTLV-1-associated myelopathy/tropical spastic paraparesis (HAM/TSP) (6,14). ATL is established in approximately 3–5% of infected individuals, while 0.25–3% of HTLV-1 carriers develop HAM/TSP (6,14). Different parts of the world such as southwestern Japan, parts of Africa, central and South America, the north of Caribbean basin (9,10), and northeast of Iran have been recognized as the main endemic regions of the virus (28). HAM/TSP is a chronic progressive demyelinating inflammatory disease of the central nervous system (CNS), which predominately affects the spinal cord (20).
HTLV-1-specific cytotoxic T cells, monocytes, and macrophages play a critical role in controlling the viral replication and in inflammatory responses in HAM/TSP patients (8). These cells produce inflammatory mediators, including tumor necrosis factor alpha (TNF-α), interleukin-1β (IL-1β), IL-6, interferon gamma (IFN-γ), macrophage inflammatory protein 1α (MIP-1α), MIP-1β, and matrix metallopeptidase 9 (MMP-9), which induce and promote the inflammatory immune responses in HAM/TSP patients (8). The HTLV-1 protein Tax is also able to induce proinflammatory responses in the CNS via production of the TNF-α, IL-1β, and IL-6 by macrophages and microglial cells (3,8,14). Thus, it seems that cytokines and chemokines may be associated with an increased risk of HTLV-1 carriers in developing HAM/TSP.
The role of the innate immune response is still not yet fully understood in HTLV-1 pathogenesis (12). However, it has been demonstrated that monocytes, macrophages, and dendritic cells (DCs), which are infected by HTLV-1in-vitro and/or in-vivo, play a physiopathological role in HTLV-1 infection (7,30).
HMGB1 is a multifunctional molecule that has a critical role in innate and adaptive immune responses (13). HMGB1 is secreted actively by monocyte/macrophage in response to inflammation and released passively from necrotic cells (13). Extracellular HMGB1 have significant roles in inflammation, invasion, cell differentiation, cell proliferation, cell migration, and immune response (13). Furthermore, HMGB1 acts as an alarm in molecule and activate the innate immune response (13). It plays a role in functional maturation and migration of DCs to the lymph nodes (13). It regulates many intracellular functions, including DNA replication, transcription, and DNA repair. Receptor for advanced glycation end products (RAGE) is one of the HMGB1 receptors, which plays a crucial role as the mediator of the innate immune responses (13). It is expressed on T and B cells and also DCs, resulting in a connection between the innate and adaptive immune response (1,13,15).
Activation of RAGE is involving in the immediate inflammatory response (1,13,15). More importantly, the perpetuation of RAGE signaling sustains the inflammation and leads to the establishment of chronic inflammatory disorders (1,13,15). Binding of HMGB1 to RAGE or other receptors such as Toll-like receptors (TLRs) as TLR-4 and provokes macrophages and monocytes to release proinflammatory cytokines, chemokines, and adhesion molecules (1,13,15,32). Several studies have shown that the systemic levels of HMGB1 are increased in acute and chronic proinflammatory situations such as sepsis, pancreatitis, diabetes mellitus, rheumatic inflammatory disease, and cancer (1,15,32). RAGE is expressed at low levels in normal conditions, however, high levels of RAGE expression has been demonstrated in different pathological situations such as chronic inflammation, diabetes, and neurodegenerative disorders (1,15,32).
It has been reported that the plasma levels of HMGB1 are increased in human immunodeficiency virus (HIV)-infected individuals compared to the healthy controls (HCs) and associated with the high viral load of HIV (5,23,24,33,34). High levels of HMGB1 concentration has also been demonstrated in the plasma of ATL patients (16). Furthermore, HTLV-1-infected T cell line expresses high HMGB1 messenger RNA (mRNA) expressions in comparison to uninfected T cell lines, suggesting that it may be used as a biomarker in ATL (16). Therefore, in the present study, we examined the plasma levels of HMGB1 and RAGE and also mRNA expression of HMGB1 in HAM/TSP patients and asymptomatic carriers (ACs) of HTLV-1to clarify the role of these markers in the pathogenesis of HTLV-1 infection.
Materials and Methods
Study population
The study population consisted of 21 patients with HAM/TSP, 21 HTLV-1 ACs, and 21 HCs who attended the Department of Neurology, Ghaem Hospital, Mashhad University of Medical Sciences (MUMS), Mashhad, Iran. All patients had fulfilled the diagnostic criteria according to the World Health Organization (WHO). Neurological and physical examinations were carried out by a single neurologist. They had positive serological results in HTLV-1 screening and confirmatory tests. Patients were examined for hepatitis B virus (HBV), hepatitis C virus (HCV), and HIV, and their results were negative.
The ACs group was obtained from individuals who referred to the Mashhad blood transfusion center, and in the primary test on their blood, HTLV-1 infection was positive, but they have no symptoms of the disease. The serological test for HBV, HCV, and HIV were negative for this group. The study was approved by the Ethics Committee of MUMS (Grant No. 931253), and informed consent was obtained from all the subjects.
Sample collection
Blood samples were collected in vacutainer vials containing EDTA. For separating peripheral blood mononuclear cells (PBMCs) immediately after blood samples collection, PBMCs were separated based on density gradient using Ficoll. After centrifugation, PBMCs were washed twice with cold phosphate-buffered saline (PBS) and then harvested. To separate plasma samples, tubes were centrifuged for 15 min at 1,000 × g. Then plasma samples were collected and stored at −70°C until analysis.
RNA extraction and complementary DNA synthesis
Total RNA was extracted from the PBMCs of patients and ACs using a TriPure Isolation Kit (Roche Diagnostic GmbH, Mannheim, Germany) according to the manufacturer's instructions. The quality of extracted RNA was checked by agarose gel electrophoresis and observable visible 18s and 28s ribosomal RNA bands, also RNA purity was quantified with NanoDrop 2000 spectrophotometer (Thermo Fisher Scientific, Inc., Waltham, MA) at 260 and 280 nm wavelength as described previously (22). A total of 3 μg of RNA was reverse-transcribed to complementary DNA (cDNA) with RevertAid First Strand cDNA Synthesis Kit (Thermo Fisher Scientific, Inc.) according to the manufacturer's protocol (22).
Quantitative TaqMan real-time polymerase chain reaction
Primer and probe for HMGB1 were designed based on sequence data on the NCBI (National Center for Biological Information) website database using Beacon Designer Software (version 7.9). To avoid unwanted amplification of genomic DNA, polymerase chain reaction (PCR) primers or probes were designed on exon–exon boundaries. Specificity of primers were rechecked by conventional PCR using temperatures from 55°C to 65°C, and then PCR products were examined by undertaking on agarose gel electrophoresis and visualizing 96 bp bands for glyceraldehyde-3-phosphate dehydrogenase (GAPDH) and 123 bp band for HMGB1. The sequence of primers and probes were as follows: GAPDH forward primer: 5′-AGCCTCAAGATCATCAGCAATG-3′, reverse primer: 5′-GAGTCCTTCCACGATACCAAAG-3′, and Probe: FAM-CTCCTGCACCACCAACTGCTTAGCACC-BHQ-I (22) and for HMGB1 forward primer: 5′-GCCTCCTTCGGCCTTCTTC-3′, reverse primer: 5′-GCAGCAGTGTTATTCCACATCTC-3′ and Probe: FAM-TCTTCGCAACATCACCAATGGACAGGC-BHQ-I.
The real-time PCR was followed by a Rotor-Gene 6000 Cycler (Qiagen). Real-time PCR was carried out by TaqMan method in a 10 μL mixture as a final volume, by using TaqMan universal PCR master mix (Takara Bio, Inc., Kyoto, Japan) and FAM-BHQ-I-labeled probes to measure expression (22). The Reaction followed by one cycle at 94°C for 2 min, 45 cycles at 94°C for 15 sec and 62°C for 1 min (22). For each sample, an endogenous control was performed by using GAPDH. Then, the result of HMGB1 expression was normalized to GAPDH expression as a reference gene.
Measurement of HMGB1 and RAGE
The plasma levels of HMGB1 in HAM/TSP patients and ACs were measured by using the precoated sandwich enzyme-linked immunosorbent assay (ELISA; IBL) according to the manufacturer's protocol; and also soluble RAGE (sRAGE) levels in HAM/TSP patients and carriers were measured using ELISA kit (R&D Systems, Minneapolis, MN) according to the manufacturer's protocol. For coating antibody in the microplate, first capture antibody is reconstituted with 1.0 mL of PBS, then diluted to the working concentration in PBS; Immediately 100 μL of diluted capture antibody is coated in a 96-well microplate and incubated at room temperature for 24 h; Then aspirated each well and washed with wash buffer for 3 times. Plates blocked by adding 300 μL of reagent diluent and incubated for a minimum of 1 h at room temperature (1% bovine serum albumin in PBS; PH 7.2–7.4 used as reagent diluent). After washing, plates were ready for use. All steps were done according to the manufacturer's protocol. Optical densities were determined by an automated ELISA plate reader (MRP4; Hiperion). Results were calculated and compared against standard curve obtained using the standards provided in the kits by the manufacturer's protocols.
Statistical analysis
Statistical analysis was performed using SPSS version 16 (SPSS, Inc., Chicago, IL), and the graphs were drawn using GraphPad Prism version 6.0 (San Diego, CA). To determine the normality of the results a Kolmogorov–Smirnov test was used. The significant differences between nonparametric results were analyzed using Kruskal–Wallis nonparametric test, and parametric results were analyzed using a one-way analysis of variance test. To compare variables between two independent groups, the Mann–Whitney U-test was used. Also, for correlations, data were analyzed using the Spearman's rank correlation coefficient test. Finally, p-values <0.05 were considered statistically significant.
Results
The studied groups consisted of 21 HAM/TSP patients, 21 HTLV-1 ACs, and 21 HCs. The mean age of patients was 46.54 ± 11.28. The mean age of ACs and HCs was 45.62 ± 10.32 and 45.38 ± 11.34, respectively. There were 2 males and 19 females in each group. No significant difference in age and gender was observed among the groups.
No differences in HMGB1 mRNA expression and plasma levels among HAM/TSP patients, ACs, and HCs
The mRNA expression of HMGB1 was quantified by real-time PCR. The mean mRNA expression in HAM/TSP patients and ACs were 4.54 ± 1.854 and 3.75 ± 1.009, respectively, and no significant differences were observed between the two groups (p = 0.084). The HMGB1 mRNA expression in HCs group was 4.093 ± 0.58 and no significant differences were observed with HAM/TSP patients and ACs (p = 0.06, p = 0.11, respectively) (Fig. 1).
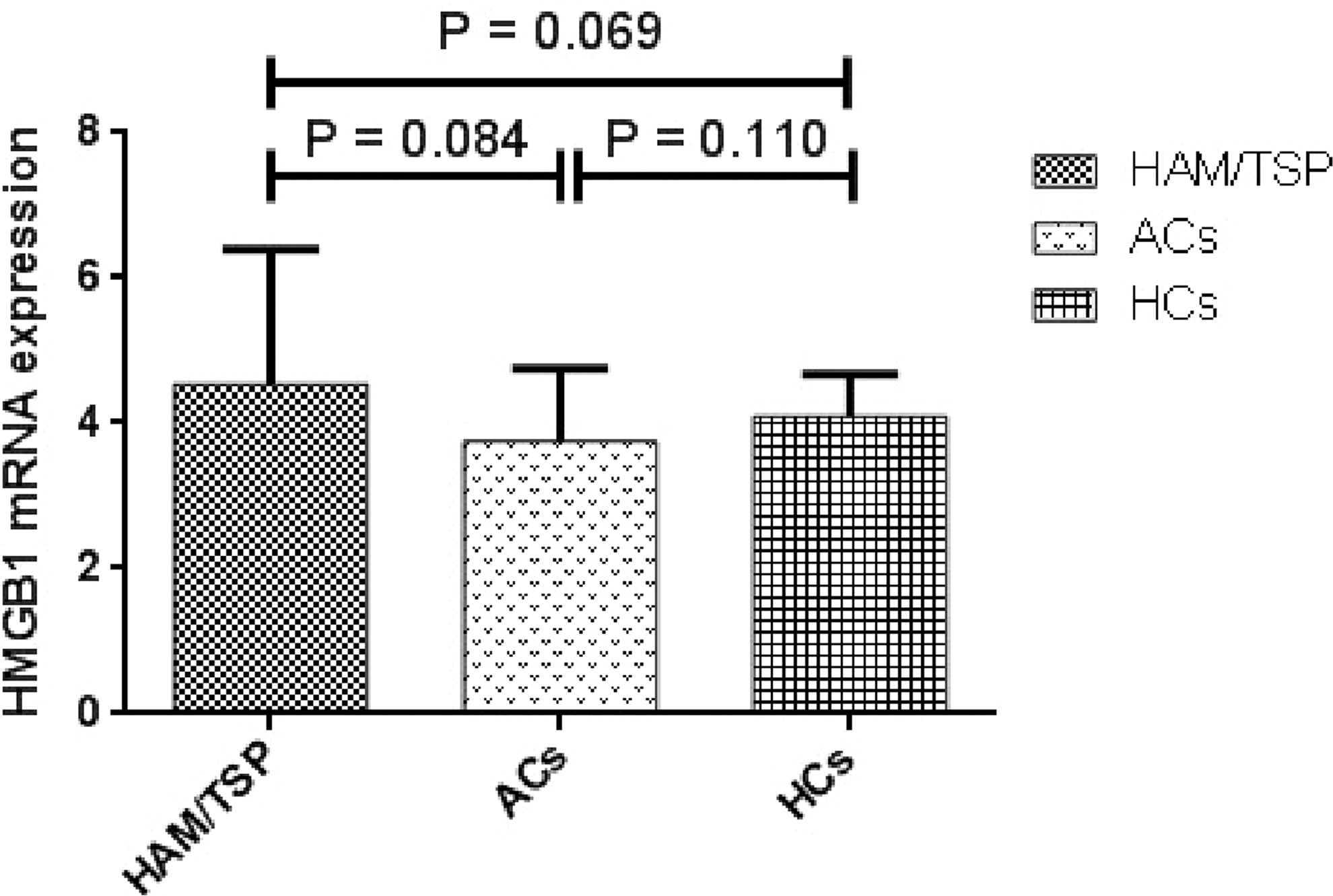
mRNA expression levels of HMGB1 in HAM/TSP patients (ACs) and HCs. No significance in HMGB1 mRNA expression was observed among the groups (p > 0.05). ACs, asymptomatic carriers; HAM/TSP, human T cell lymphotropic type 1-associated myelopathy/tropical spastic paraparesis; HCs, healthy controls; HMGB1, High Mobility Group Box 1; mRNA, messenger RNA.
The mean plasma levels of HMGB1 in HAM/TSP patients, ACs, and HCs were 1.01 ± 0.36, 0.66 ± 0.27, and 0.67 ± 0.11, respectively, and no significant differences were observed between studied groups (p > 0.05) (Fig. 2).
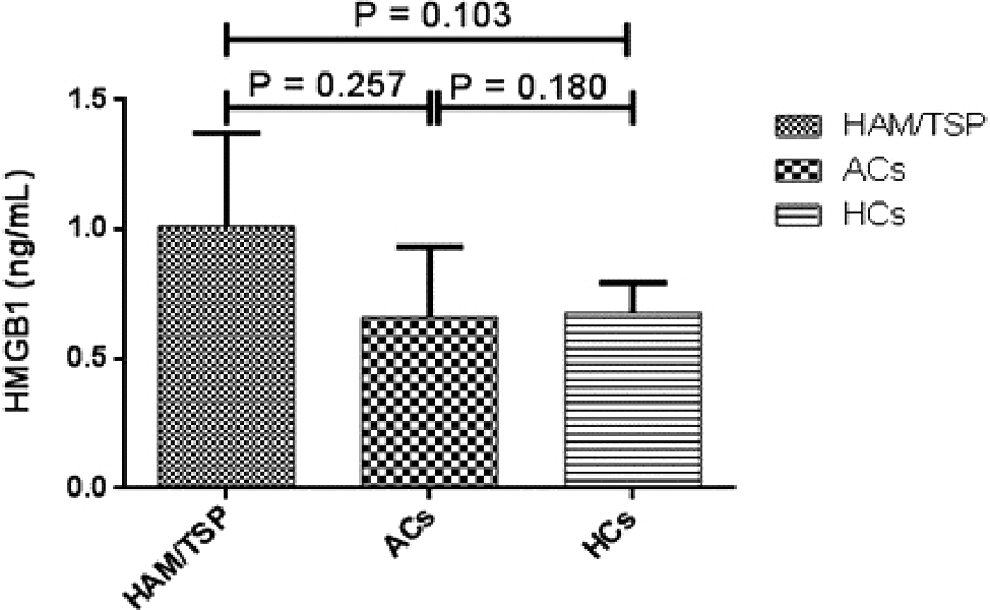
Plasma levels of HMGB1 in patients with HAM/TSP, asymptomatic group (ACs), and HCs: No significant differences were observed between the studied groups (p > 0.05).
Increased plasma levels of sRAGE in ACs and HCs
The plasma levels of sRAGE were higher in ACs (1,894.74 ± 962) in comparison to the HAM/TSP patients (603.53 ± 217) and HCs (1,111.28 ± 635), and significant differences were detected between the HAM/TSP patients and ACs (p < 0.001) and ACs with HCs (p < 0.001) (Fig. 3).
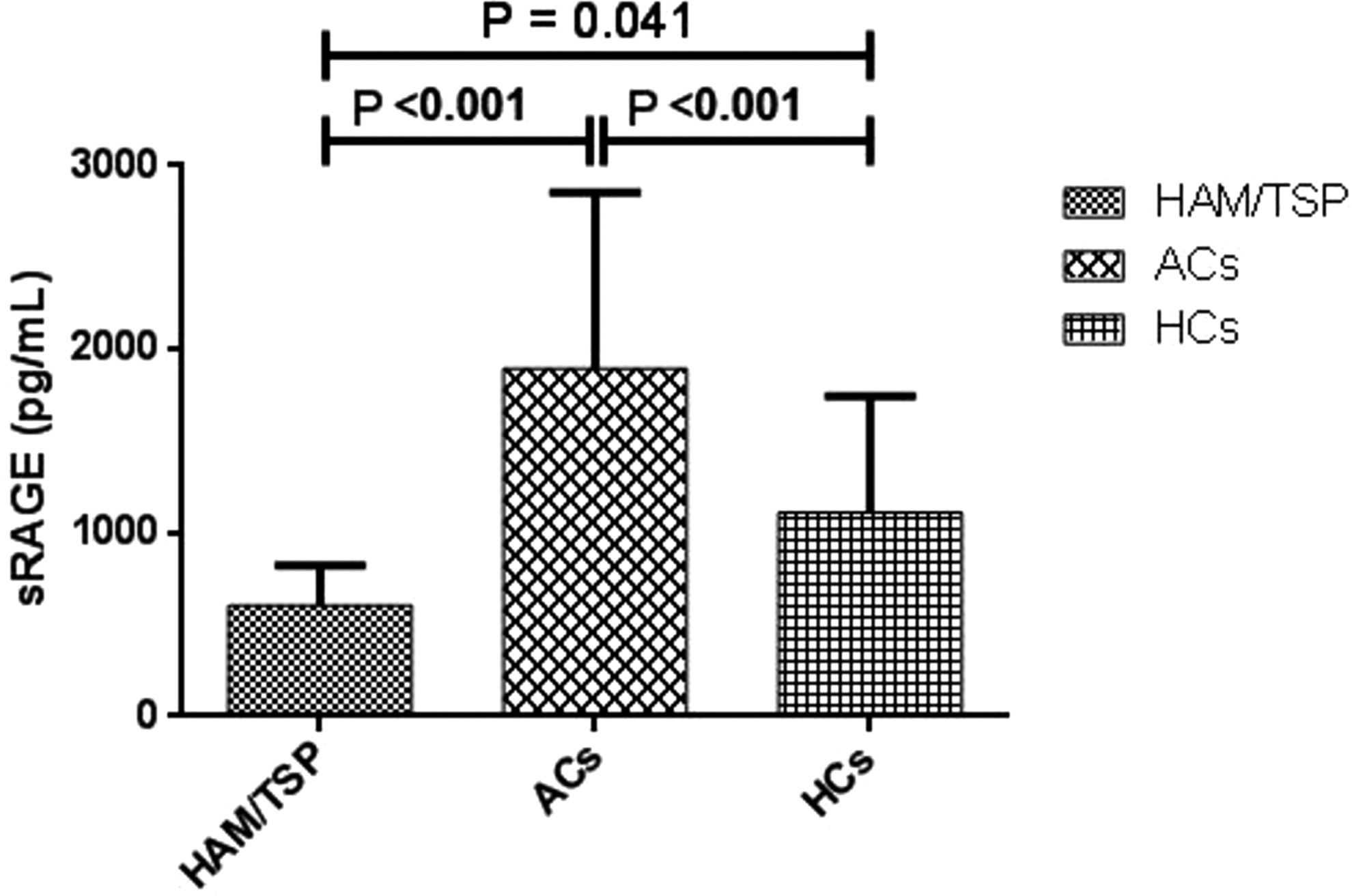
Plasma levels of sRAGE in patients with HAM/TSP, asymptomatic group (ACs), and HCs: the plasma levels of RAGE was higher in ACs compared to HAM/TSP patients and HCs and significant differences were observed between HAM/TSP patients and ACs (p < 0.001) and ACs with HCs (p < 0.001). sRAGE, soluble receptor for advanced glycation end products.
Correlations
In the present study, a positive correlation was only found between the serum levels of HMGB and mRNA expression in HAM/TSP patients (R = 0.47, p < 0.03). There were no significant correlations between age, clinical symptoms, HMGB1 mRNA expression, and protein levels and sRAGE with each other among the studied groups (p > 0.05).
Discussion
It is well known that HAM/TSP is a chronic neuroinflammatory disease, which is associated with high levels of inflammatory cytokines such as TNF-α, IL-1β, IL-6, IFN-γ, MIP-1α, and MIP-1β that have an important role in HAM/TSP pathogenesis (8,29). It has been proposed that extracellular HMGB1 is a potent inducer of proinflammatory cytokines and chemokines such as TNF-α, IL-1β, IL-6, IL-8, and MIP-1α and β (13).
The HTLV-1 Tax has potential transactivation properties, and HMGB1 expression is upregulated by Tax (35,38). It has been also reported that the plasma levels of HMGB1 in the ATL patients are significantly higher than that of HCs (16). In addition, the HMGB1 mRNA expression is also higher expressed in HTLV-1-infected T cell line than uninfected T cell line, suggesting that HMGB1 could be used not only as a biomarker but also as a therapeutic target in patients with ATL (16).
HMGB1 expression and its serum level increases in several diseases such as AIDs (5,23,24,33,34) and multiple sclerosis (2,19). Infection of mice by the respiratory syncytial virus (RSV) increased sRAGE levels, while HMGB1 levels did not upregulate in the infected model (21), suggesting that circulating HMGB1 is positively or negatively associated with sRAGE levels, and in this case, sRAGE is able to regulate HMGB1 activity or abolishes HMGB1 levels. In the present study, the levels of HMGB1 were higher in HAM/TSP patients than that of ACs, but did not reach statistical significance. Moreover, we could not also find any correlation between HMGB1 and sRAGE levels in HAM/TSP patients. It should be taken into account that circulating levels of HMGB1 shows a wide range, even in healthy individuals, thus measurement of this marker is not specific for certain diseases and could not easily be used as a biomarker in the prediction and prognosis of diseases (25).
sRAGE builds up a communication between innate and adaptive immune response (4,15). The interaction between RAGE and its ligands advanced glycation end products (AGEs) activates different cellular signaling pathways, such as nuclear factor-kappa binding (NF-κB) and thus leading to an inflammatory response (4,15). RAGE protects individuals against proinflammatory responses by RSV infection and in the prevention of syncytia formation in the lower airways. Although cellular transmission through membrane RAGE leads to an inflammatory response, the interaction between sRAGE and its ligands decreases the inflammation in RSV-infected individuals (31).
There are different hypothesis regarding the role of sRAGE in inflammation. Several studies reported that sRAGE produces proinflammatory cytokines and causes macrophage migration and differentiation (17,36,37). However, others showed that sRAGE inhibits leukocyte recruitment in inflammatory disease such as inflammatory diabetes, peritonitis, septicemia, and arthritis (11,18,37). It is supposed that sRAGE acts as a decoy receptor, competing with RAGE for its ligand, AGEs (26,27). Investigation of HMGB1 levels in Cerebrospinal fluid (CSF) could help to clarify the role of this marker in the outcome of HTLV-1 infection. One limitation of our study is the small sample size, thus further studies with larger sample sizes are needed to clarify the role of HMGB-1 in the pathogenesis of HTLV-1-associated diseases.
In the present study, we demonstrated that the plasma levels of sRAGE in ACs are significantly higher than HAM/TSP.
It seems that low level of sRAGE in HAM/TSP patients compared with ACs is not able to prevent sRAGE to connect with AGEs. Signaling through membrane RAGE induces an inflammatory response, while the interactions between sRAGE and AGEs prevent binding of circulating AGEs to RAGE and play an important role in the modulation of the inflammatory response in the host. Thus, low levels of sRAGE in HAM/TSP patients, in comparison with ACs, could not inhibit AGEs/RAGE signaling pathway, which might lead to the inflammatory response that is observed in HAM/TSP patients, while high levels of sRAGE in ACs efficiently control the RAGE signaling pathway and inhibit leukocyte recruitment and the inflammatory responses (26).
Thus, high production of sRAGE in ACs could modulate the inflammatory immune response and prevents developing HAM/TSP. Furthermore, anti-AGE–RAGE defense mechanisms such as degradation of AGE and sRAGE, which compete with RAGE for AGE, are present in the body to counterbalance the effects of AGEs and AGE–RAGE interaction (27) and might lead to modulatory and inflammatory responses in ACs and HAM/TSP patients, respectively. It should be taken into account that viral factors such as proviral load, Tax, and HBZ, which are expressed at high levels in HAM/TSP patients compared to ACs (14), might counterbalance the effects of AGEs and AGE-RAGE interaction and downregulate sRAGE and induce an inflammatory response in HAM/TSP patients.
In conclusion, the results of the present study showed that high levels of sRAGE might be associated with a lower risk of HTLV-1 carriers to develop HAM/TSP. Further studies with larger sample sizes are needed to clarify this issue. Furthermore, evaluation of HMGB1 and RAGE at the CNS would be informative to understand the role these markers have in HTLV-1 infection.
Footnotes
Acknowledgment
This work was supported by MUMS (Grant No. 931253).
Author Disclosure Statement
The authors declare that there is no conflict of interest to declare.